
Biocontrol of Invasive Conical Snails by the Parasitoid Fly Sarcophaga villeneuveana in South Australia 20 Years after Release


Abstract
:Simple Summary
Abstract
1. Introduction
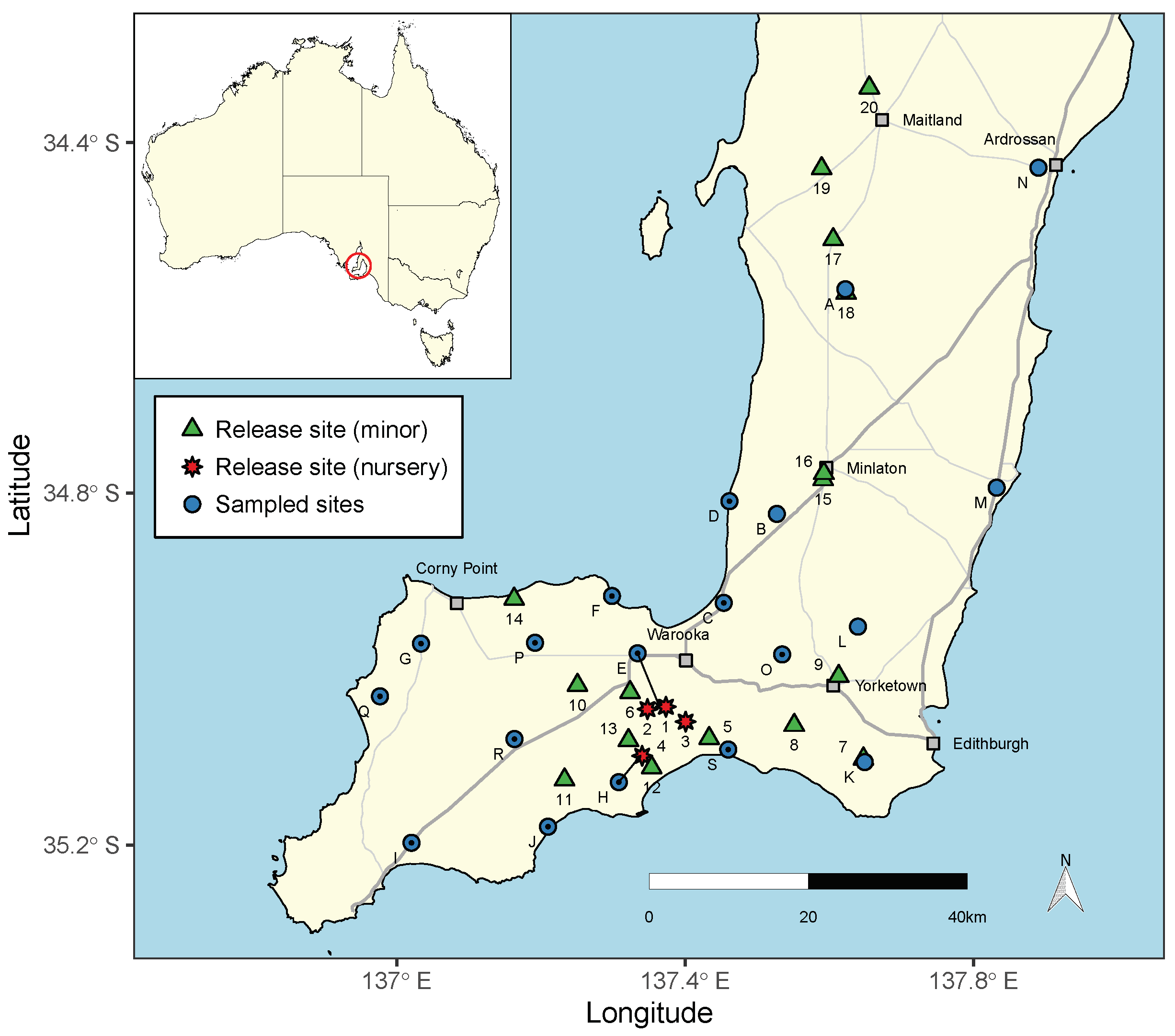
2. Methods
2.1. Field Sampling and Data Collection
2.2. Data Analysis
3. Results
3.1. Abundance of Conical Snails
3.2. Parasitism of Conical Snails by S. villeneuveana
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker, G.H. The Biology and Control of White Snails (Mollusca: Helicidae), Introduced Pests in Australia; CSIRO: Melbourne, Australia, 1986. [Google Scholar] [CrossRef]
- Baker, G.H. The population dynamics of the Mediterranean snails Cernuella virgata, Cochlicella acuta (Hygromiidae) and Theba pisana (Helicidae) in pasture-cereal rotations in South Australia: A 20-year study. Aust. J. Exp. Agric. 2008, 48, 1514–1522. [Google Scholar] [CrossRef]
- Atlas of Living Australia Website. Species Page, Cochlicella acuta. Available online: https://bie.ala.org.au/species/urn:lsid:biodiversity.org.au:afd.taxon:1996efa6-2db2-424f-a822-555e2d67cdc5 (accessed on 21 August 2021).
- Atlas of Living Australia Website. Species Page, Prietocella barbara. Available online: https://bie.ala.org.au/species/urn:lsid:biodiversity.org.au:afd.taxon:e9ba197e-e04d-4a5c-9b58-9ccdcb20d4c5 (accessed on 21 August 2021).
- Leonard, E.; Hopkins, D.C.; Baker, G.H. (Eds.) Bash Em Burn Em Bait Em: Integrated Snail Managament in Crops and Pastures; South Australian Research and Development Institute: Adelaide, Australia, 2003. [Google Scholar]
- Baker, G.H.; Hawke, B.G.; Vogelzang, B.K. Life history and population dynamics of Cochlicella acuta (Müller) (Gastropoda: Helicidae) in a pasture-cereal rotation. J. Molluscan. Stud. 1991, 57, 259–266. [Google Scholar] [CrossRef]
- Coupland, J.B.; Baker, G.H. Host distribution, larviposition behaviour and generation time of Sarcophaga penicillata (Diptera: Sarcophagidae), a parasitoid of conical snails. B Entomol. Res. 1994, 84, 185–189. [Google Scholar] [CrossRef]
- Coupland, J.B.; Espiau, A.; Baker, G.H. Seasonality, longevity and infection efficiency of Salticella fasciata (Diptera, Sciomyzidae), a candidate for the biological control of pest helicid snails. Biol. Control 1994, 4, 32–37. [Google Scholar] [CrossRef]
- Coupland, J.B. Diptera associated with snails collected in south-western and west-Mediterranean Europe. Vertigo 1994, 3, 19–26. [Google Scholar]
- Coupland, J.B.; Baker, G.H. The potential of several species of terrestrial Sciomyzidae as biological control agents of pest helicid snails in Australia. Crop. Prot. 1995, 14, 573–576. [Google Scholar] [CrossRef]
- Coupland, J.B. Influence of snail feces and mucus on oviposition and larval behavior of Pherbellia cinerella (Diptera: Sciomyzidae). J. Chem. Ecol. 1996, 22, 183–189. [Google Scholar] [CrossRef]
- Coupland, J.B.; Baker, G.H. Search for biological control agents of invasive Mediterranean snails. In Biological Control: A Global Perspective; CABI: Wallingford, UK, 2007; pp. 7–12. [Google Scholar] [CrossRef]
- Leyson, M.; Hopkins, D.C.; Charwat, S. Release and establishment in South Australia of Sarcophaga penicillata (Diptera: Sarcophagidae), a biological control agent for Cochlicella acuta (Mollusca: Hygromiidae). In Slugs & Snails: Veterinary & Environmental Perspectives, BCPC Symposium Proceedings No. 80; Dussart, G.B.J., Ed.; British Crop Protection Council: Farnham, UK, 2003; pp. 295–300. [Google Scholar]
- Thomann, T.; Jourdan, M.; Richet, R.; Sheppard, A.; Baker, G.H. Parasitism of the conical snail, Cochlicella acuta, by the fly, Sarcophaga villeneuveana, in south-western Europe. BioControl 2020, 65, 673–679. [Google Scholar] [CrossRef]
- Fendane, Y.; Richet, R.; Thomann, T.; Jourdan, M.; Baker, G.; Ghamizi, M.; Sheppard, A. First records of flesh flies (Diptera: Sarcophagidae) emerging from terrestrial snails in Morocco. Afr. Entomol. 2018, 26, 124–130. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Hijmans, R.J. geosphere: Spherical Trigonometry. R Package Version 1.5-10. 2019. Available online: https://CRAN.R-project.org/package=geosphere (accessed on 21 August 2021).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N. A protocol for conducting and presenting results of regression-type analyses. Methods Ecol. Evol. 2016, 7, 636–645. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level / Mixed) Regression Models. R Package Version 0.4.1. 2021. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 21 August 2021).
- Fox, J.; Weisberg, S. An R companion to applied regression. J. Open Source Softw. 2019, 6, 3139. [Google Scholar] [CrossRef]
- Lenth, R.V. emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.6.1. 2021. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 21 August 2021).
- Harrison, X.A. A comparison of observation-level random effect and beta-binomial models for modelling overdispersion in binomial data in ecology & evolution. PeerJ 2015, 3, e1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grevstad, F.S. Factors influencing the chance of population establishment: Implications for release strategies in biocontrol. Ecol. Appl. 1999, 9, 1439–1447. [Google Scholar] [CrossRef]
- Possingham, H.; Shea, M. Optimal release strategies for biological agents: An application of stochastic dynamic programming to population management. J. Appl. Ecol. 2000, 37, 77–86. [Google Scholar] [CrossRef]
- Hopper, K.R.; Roush, R.T. Mate finding, dispersal, number released, and the success of biological control introductions. Ecol. Entomol. 1993, 18, 321–331. [Google Scholar] [CrossRef]
- Jourdan, M.; Thomann, T.; Kriticos, D.J.; Bon, M.C.; Sheppard, A.; Baker, G.H. Sourcing effective biological control agents of conical snails, Cochlicella acuta, in Europe and north Africa for release in southern Australia. Biol. Control 2019, 134, 1–14. [Google Scholar] [CrossRef]
- Jourdan, M.; Thomann, T.; Richet, R.; Fendane, Y.; Ghamizi, M.; Bon, M.C.; Sheppard, A.; Baker, G.H. Genetic variability in the parasitic fly, Sarcophaga villeneuveana, in south-western Europe and Morocco. BioControl 2020, 65, 59–70. [Google Scholar] [CrossRef]
- Quicke, D.L.J. Parasitic wasps; Chapman & Hall: London, UK, 1997. [Google Scholar]
- Olson, D.M.; Takasu, K.; Lewis, W.J. Food needs of adult parasitoids: Behavioral adaptations and consequences. In Plant-Provided Food for Carnivorous Insects: A Protective Mutualism and Its Applications; Wäckers, F.L., van Rijn, P.C.J., Bruin, J., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 137–147. [Google Scholar] [CrossRef]
- Jervis, M.A.; Ellers, J.; Harvey, J.A. Resource acquisition, allocation, and utilization in parasitoid reproductive strategies. Annu. Rev. Entomol. 2008, 53, 361–385. [Google Scholar] [CrossRef] [Green Version]
- Collatz, J.; Romeis, J. Flowers and fruits prolong survival of Drosophila pupal parasitoids. J. Appl. Entomol. 2021, 145, 629–634. [Google Scholar] [CrossRef]
- Hawley, J.; Simpson, S.J.; Wilder, S.M. Flesh flies regulate the consumption of 3 macronutrients to maximize lifespan and egg production. Behav. Ecol. 2016, 27, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Hein, S.; Dorn, S. The parasitoid of a fruit moth caterpillar utilizes fruit components as nutrient source to increase its longevity and fertility. Biol. Control 2008, 44, 341–348. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Collatz, J.; Dubsky, V.; Entling, M.H.; Najar-Rodriguez, A.J.; Jacot, K. Tailored flower strips promote natural enemy biodiversity and pest control in potato crops. J. Appl. Ecol. 2016, 53, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Gilchrist, A.S.; Cameron, E.C.; Sved, J.A.; Meats, A.W. Genetic consequences of domestication and mass rearing of pest fruit fly Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Entomol. 2012, 105, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Francuski, L.; Djurakic, M.; Ludoški, J.; Hurtado, P.; Pérez-Bañón, C.; Ståhls, G.; Rojo, S.; Milankov, V. Shift in phenotypic variation coupled with rapid loss of genetic diversity in captive populations of Eristalis tenax (Diptera: Syrphidae): Consequences for rearing and potential commercial use. J. Econ. Entomol. 2014, 107, 821–832. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | Total | |||
|---|---|---|---|---|---|
| Jan | Apr | Jan | Apr | ||
| No. sites with C. acuta | 14 | 19 | 19 | 18 | 19 |
| No. sites with fly detected | 8 | 13 | 13 | 13 | 13 |
| No. suitable snails >5 | 37,347 | 9512 | 26,888 | 11,926 | 85,673 |
| No. live snails | 36,261 | 8352 | 23,872 | 8252 | 76,737 |
| No. dead snails (unknown) | 597 | 349 | 2499 | 3548 | 6993 |
| No. shells with fly larva | 1 | 15 | 8 | 1 | 25 |
| No. shells with fly pupa | 54 | 55 | 31 | 25 | 165 |
| No. shells with dead fly | 23 | 56 | 35 | 0 | 114 |
| No. shells with open fly pupa | 411 | 685 | 443 | 100 | 1639 |
| No. flies emerged during rearing | 104 | 189 | 69 | 23 | 385 |
| Parasitism rate (all sites) | 1.31% | 8.53% | 1.92% | 1.06% | 2.27% |
| Parasitism rate (fly-positive sites) | 1.78% | 10.72% | 2.30% | 1.18% | 2.85% |
| 2019 | 2020 | Total | |||
|---|---|---|---|---|---|
| Jan | Apr | Jan | Apr | ||
| No. sites with C. barbara | 5 | 9 | 11 | 5 | 11 |
| No. sites with fly detected | 3 | 5 | 7 | 4 | 7 |
| No. suitable snails >5 | 684 | 698 | 872 | 158 | 2412 |
| No. live snails | 638 | 636 | 820 | 124 | 2218 |
| No. dead snails (unknown) | 41 | 8 | 40 | 32 | 121 |
| No. shells with fly larva | 0 | 1 | 1 | 0 | 2 |
| No. shells with fly pupa | 0 | 5 | 1 | 1 | 7 |
| No. shells with dead fly | 0 | 2 | 2 | 0 | 4 |
| No. shells with open fly pupa | 5 | 46 | 8 | 1 | 60 |
| No. flies emerged during rearing | 1 | 9 | 0 | 0 | 10 |
| Parasitism rate (all sites) | 0.73% | 7.74% | 1.38% | 1.27% | 3.03% |
| Parasitism rate (fly-positive sites) | 0.91% | 8.88% | 1.39% | 1.54% | 3.39% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muirhead, K.A.; Perry, K.D. Biocontrol of Invasive Conical Snails by the Parasitoid Fly Sarcophaga villeneuveana in South Australia 20 Years after Release. Insects 2021, 12, 865. https://doi.org/10.3390/insects12100865
Muirhead KA, Perry KD. Biocontrol of Invasive Conical Snails by the Parasitoid Fly Sarcophaga villeneuveana in South Australia 20 Years after Release. Insects. 2021; 12(10):865. https://doi.org/10.3390/insects12100865
Chicago/Turabian StyleMuirhead, Kate A., and Kym D. Perry. 2021. "Biocontrol of Invasive Conical Snails by the Parasitoid Fly Sarcophaga villeneuveana in South Australia 20 Years after Release" Insects 12, no. 10: 865. https://doi.org/10.3390/insects12100865
APA StyleMuirhead, K. A., & Perry, K. D. (2021). Biocontrol of Invasive Conical Snails by the Parasitoid Fly Sarcophaga villeneuveana in South Australia 20 Years after Release. Insects, 12(10), 865. https://doi.org/10.3390/insects12100865