
Temporal Variation Dominates in Local Carabid Beetle Communities in Korean Mountains

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Field Survey
2.2. Environmental Factors
2.3. Analysis of Communities
2.4. Variance Partitioning
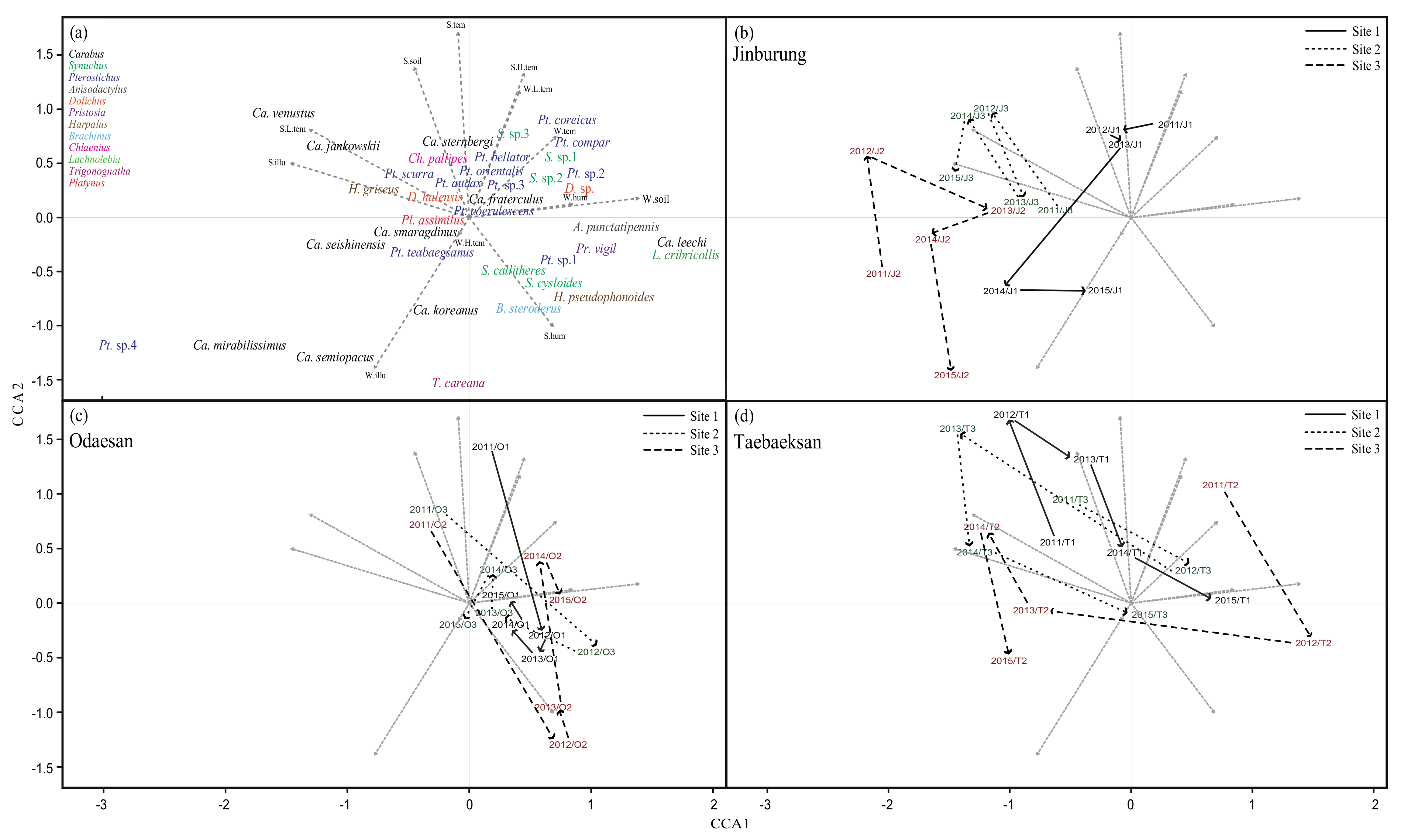
3. Results
3.1. Spatial and Temporal Variation in Environmental Factors
3.2. Spatial and Temporal Variation in Communities
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Site | Latitude Longitude | Slope | Altitude (m) | Gradient (°) | Dominant Tree Species | Understorey Vegetation Species | |
|---|---|---|---|---|---|---|---|
| Jinbu-ryung | J1 | 38°16′00.31″ N 128°23′07.14″ E | NW | 694 | 20 | Quercus mongolica, Tilia mandshurica, Synplocos chinensis | Synurus deltoides, Impatiens textori, Isodon excisus |
| J2 | 38°15′53.01″ N 128°20′01.94″ E | N | 1038 | 10 | Quercus mongolica, Acer pseudosieboldianum, Tilia mandshurica | Carex siderosticta, Hepatica asiatica, Primula jesoana | |
| J3 | 38°17′03.36″ N 128°21′40.76″ E | W | 323 | 10 | Styrax obassia, Quercus mongolica, Pinus densiflora | Pseudostellaria palibiniana, Dioscorea quinqueloba, Isodon japonicus | |
| Odae-san | O1 | 38°44′31.86″ N 128°37′11.08″ E | W | 741 | 25 | Quercus mongolica, Ulmus davidiana var. japonica, Fraxinus rhynchophylla | Brachybotrys paridiformis, Carex siderosticta, Lespsdeza japonica |
| O2 | 38°45′58.29″ N 128°36′20.22″ E | SW | 1062 | 32 | Quercus mongolica, Tilia mandshurica, Acer pseudosieboldianum | Sasa borealis, Carex siderosticta, Viola albida | |
| O3 | 38°47′10.76″ N 128°37′09.83″ E | N | 668 | 20 | Fraxinus rhynchophylla, Pinus densiflora, Acer pseudosieboldianum | Disporums milacinum, Arisaema amurense, Isodon japonicus | |
| Tae-baek-san | T1 | 37° 7′22.90″ N 128°57′32.88″ E | W | 968 | 28 | Betula davurica, Pinus densiflora, Quercus mongolica | Athyrium niponicum, Syneilesis palmata, Festuca ovina |
| T2 | 37°09′38.22″ N 128°53′10.35″ E | W | 1274 | 36 | Quercus mongolica, Ulmus davidiana var. japonica, Fraxinus rhynchophylla | Astilbe koreana, Carex humilis, Viola albida | |
| T3 | 37°06′36.02″ N 128°51′30.24″ E | SW | 704 | 35 | Quercus mongolica, Fraxinus rhynchophylla, Tilia mandshurica | Sasa borealis, Tripterygium regelii, Carex siderosticta | |
| Jinburyung | Odaesan | Taebaeksan | Total | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | 2011 | 2012 | 2013 | 2014 | 2015 | 2011 | 2012 | 2013 | 2014 | 2015 | 2011 | 2012 | 2013 | 2014 | 2015 | |
| Carabus (Eucarabus) sternbergi | 12 | 74 | 53 | 22 | 30 | 115 | 55 | 59 | 199 | 77 | 21 | 15 | 88 | 31 | 56 | 905 |
| Carabus (Leptocarabus) seishinensis | 69 | 31 | 41 | 62 | 53 | 25 | 29 | 34 | 61 | 41 | 22 | 1 | 41 | 87 | 63 | 660 |
| Carabus (Leptocarabus) semiopacus | 3 | 26 | 14 | 21 | 103 | 10 | 44 | 26 | 50 | 7 | 4 | 85 | 393 | |||
| Carabus (Leptocarabus) koreanus | 8 | 1 | 9 | |||||||||||||
| Carabus (Morphocarabus) venustus | 1 | 27 | 26 | 22 | 4 | 7 | 8 | 3 | 13 | 13 | 2 | 126 | ||||
| Carabus (Tomocarabus) fraterculus | 2 | 5 | 2 | 1 | 5 | 12 | 9 | 3 | 1 | 5 | 3 | 48 | ||||
| Carabus (Coptolabrus) smaragdinus | 5 | 7 | 3 | 2 | 6 | 7 | 3 | 5 | 11 | 13 | 3 | 65 | ||||
| Carabus (Coptolabrus) jankowskii | 1 | 8 | 12 | 3 | 6 | 3 | 9 | 5 | 4 | 7 | 1 | 25 | 35 | 17 | 136 | |
| Carabus (Acoptolabrus) mirabilissimus | 7 | 1 | 1 | 1 | 1 | 1 | 12 | |||||||||
| Carabus (Acoptolabrus) leechi | 1 | 1 | ||||||||||||||
| Synuchus callitheres | 4 | 9 | 19 | 1 | 57 | 23 | 56 | 74 | 100 | 97 | 2 | 10 | 19 | 86 | 557 | |
| Synuchus cycloides | 3 | 3 | 10 | 53 | 112 | 66 | 82 | 4 | 101 | 434 | ||||||
| Synuchus nitidus | 29 | 31 | 41 | 23 | 59 | 4 | 65 | 83 | 259 | 115 | 22 | 29 | 50 | 88 | 898 | |
| Synuchus sp.1 | 44 | 2 | 1 | 7 | 10 | 31 | 22 | 5 | 67 | 38 | 31 | 12 | 10 | 5 | 285 | |
| Synuchus sp.2 | 3 | 2 | 5 | 4 | 95 | 39 | 3 | 14 | 10 | 173 | ||||||
| Synuchus sp.3 | 44 | 14 | 58 | |||||||||||||
| Platynus assimilis | 7 | 3 | 10 | 11 | 27 | 42 | 27 | 56 | 2 | 2 | 40 | 227 | ||||
| Poecilus versicolor | 5 | 23 | 20 | 7 | 26 | 13 | 43 | 9 | 43 | 28 | 8 | 37 | 37 | 299 | ||
| Pterostichus orientalis | 2 | 12 | 7 | 5 | 41 | 30 | 40 | 7 | 7 | 11 | 18 | 180 | ||||
| Pterostichus audax | 11 | 17 | 12 | 19 | 41 | 7 | 3 | 38 | 52 | 69 | 13 | 10 | 40 | 62 | 394 | |
| Pterostichus scurra | 6 | 30 | 9 | 12 | 30 | 19 | 15 | 36 | 43 | 49 | 2 | 6 | 31 | 104 | 392 | |
| Pterostichus compar | 14 | 11 | 8 | 31 | 5 | 31 | 29 | 4 | 10 | 66 | 209 | |||||
| Pterostichus bellator | 1 | 10 | 8 | 6 | 27 | 19 | 33 | 31 | 42 | 3 | 6 | 7 | 8 | 25 | 226 | |
| Pterostichus teabaegsanus | 1 | 10 | 1 | 1 | 4 | 17 | ||||||||||
| Pterostichus coreicus | 18 | 18 | ||||||||||||||
| Pterostichus sp.1 | 5 | 5 | 1 | 14 | 6 | 16 | 20 | 2 | 2 | 71 | ||||||
| Pterostichus sp.2 | 9 | 2 | 4 | 17 | 19 | 51 | ||||||||||
| Pterostichus sp.3 | 4 | 5 | 9 | |||||||||||||
| Dolichus halensis | 9 | 7 | 10 | 1 | 2 | 12 | 10 | 34 | 4 | 12 | 28 | 129 | ||||
| Dolichus sp. | 10 | 6 | 16 | |||||||||||||
| Pristosia vigil | 4 | 1 | 9 | 15 | 35 | 6 | 72 | 119 | 105 | 94 | 3 | 12 | 3 | 15 | 62 | 547 |
| Anisodactylus punctatipennis | 1 | 1 | 13 | 5 | 20 | |||||||||||
| Harpalus griseus | 1 | 1 | 3 | 2 | 7 | |||||||||||
| Brachinus stenoderus | 5 | 5 | 10 | |||||||||||||
| Lachnolebia cribricollis | 3 | 3 | ||||||||||||||
| Chlaenius pallipes | 4 | 4 | 1 | 9 | ||||||||||||
| Trigonognatha coreana | 1 | 1 | ||||||||||||||
| Total | 217 | 335 | 268 | 250 | 516 | 368 | 536 | 806 | 1362 | 1087 | 143 | 52 | 276 | 424 | 967 | 7607 |
| Mountain | Site within a Mountain | Year | ||||
|---|---|---|---|---|---|---|
| χ22 | p | χ26 | p | χ24 | p | |
| Summer mean air temperature | 0.032 | 0.968 | 4.756 | 0.0005 | 1.041 | 0.398 |
| Summer highest mean air temperature | 1.196 | 0.313 | 10.41 | < 0.0001 | 0.224 | 0.923 |
| Summer lowest mean air temperature | 1.780 | 0.181 | 2.052 | 0.068 | 1.809 | 0.146 |
| Summer mean soil temperature | 4.862 | 0.013 | 2.250 | 0.046 | 1.776 | 0.153 |
| Summer mean air humidity | 1.935 | 0.157 | 4.142 | 0.001 | 2.769 | 0.039 |
| Summer mean illumination | 5.777 | 0.006 | 3.241 | 0.007 | 0.332 | 0.855 |
| Winter mean air temperature | 0.180 | 0.836 | 5.371 | 0.0002 | 3.773 | 0.011 |
| Winter highest mean air temperature | 0.835 | 0.441 | 10.93 | <0.0001 | 0.975 | 0.432 |
| Winter lowest mean air temperature | 0.368 | 0.694 | 2.582 | 0.024 | 4.047 | 0.008 |
| Winter mean soil temperature | 1.394 | 0.259 | 2.348 | 0.038 | 2.317 | 0.074 |
| Winter mean air humidity | 0.157 | 0.855 | 3.901 | 0.002 | 1.280 | 0.294 |
| Winter mean illumination | 7.518 | 0.002 | 13.07 | <0.0001 | 0.663 | 0.622 |
| Mountain | Sites within a Mountain | Year | ||||
|---|---|---|---|---|---|---|
| χ22 | p | χ26 | p | χ24 | p | |
| Diversity | 5.207 | 0.010 | 1.443 | 0.213 | 5.379 | 0.001 |
| Richness | 9.408 | 0.0004 | 2.171 | 0.054 | 6.646 | <0.0001 |
| Abundance | 9.506 | 0.0003 | 2.273 | 0.044 | 4.856 | 0.003 |
| CCA1 | 16.950 | <0.0001 | 7.526 | <0.0001 | 0.315 | 0.866 |
| CCA2 | 10.100 | 0.066 | 1.448 | 0.211 | 2.137 | 0.930 |

References
- Huston, M. A general hypothesis of species diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Niemelä, J.; Halme, E. Habitat associations of carabid beetles in fields and forests on the Åland Islands, SW Finland. Ecography. 1992, 15, 3–11. [Google Scholar] [CrossRef]
- Niemelä, J.; Haila, Y.; Halme, E.; Pajunen, T.; Punttila, P. Small-Scale heterogeneity in the spatial distribution of carabid beetles in the southern Finnish taiga. J. Biogeogr. 1992, 19, 173–181. [Google Scholar] [CrossRef]
- Butaye, J.; Jacquemyn, H.; Honnay, O.; Hermy, M. The species pool concept applied to forests in a fragmented landscape: Dispersal limitation versus habitat limitation. J. Veg. Sci. 2002, 13, 27–34. [Google Scholar] [CrossRef]
- Summerville, K.S.; Crist, T.O. Structure and conservation of lepidopteran communities in managed forests of northeastern North America: A review. Can. Entomol. 2008, 140, 475–494. [Google Scholar] [CrossRef]
- Garden, J.G.; Mcalpine, C.A.; Possingham, H.P.; Jones, D.N. Habitat structure is more important than vegetation composition for local-level management of native terrestrial reptile and small mammal species living in urban remnants: A case study from Brisbane, Australia. Austral Ecol. 2007, 32, 669–685. [Google Scholar]
- Rich, L.N.; Miller, D.A.; Robinson, H.S.; McNutt, J.W.; Kelly, M.J. Using camera trapping and hierarchical occupancy modelling to evaluate the spatial ecology of an African mammal community. J. Appl. Ecol. 2016, 53, 1225–1235. [Google Scholar] [CrossRef]
- Kaspari, M.; O’Donnell, S.; Kercher, J.R. Energy, density, and constraints to species richness: Ant assemblages along a productivity gradient. Am. Nat. 2000, 155, 280–293. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C. Geographic range size and determinants of avian species richness. Science 2002, 297, 1548–1551. [Google Scholar] [CrossRef] [Green Version]
- Morales, M.A.; Dodge, G.J.; Inouye, D.W. A phenological mid-domain effect in flowering diversity. Oecologia 2005, 142, 83–89. [Google Scholar] [CrossRef]
- van der Putten, W.H.; Bardgett, R.D.; de Ruiter, P.C.; Hol, W.H.G.; Meyer, K.M.; Bezemer, T.M.; Bradfod, M.A.; Christensen, S.; Eppinga, M.B.; Fukami, T.; et al. Empirical and theoretical challenges in aboveground–belowground ecology. Oecologia 2009, 161, 1–14. [Google Scholar] [CrossRef]
- Butterfield, J.; Luff, M.L.; Baines, M.; Eyre, M.D. Carabid beetle communities as indicatorsof conservation potential in upland forests. For. Ecol. Manag. 1995, 79, 63–77. [Google Scholar] [CrossRef]
- Allegro, G.; Sciaky, R. Assessing the potential role of ground beetles (Coleoptera, Carabidae) as bioindicators in poplar stands, with a newly proposed ecological index (FAI). For. Ecol. Manag. 2003, 175, 275–284. [Google Scholar] [CrossRef]
- Thiele, H.U. Carabid Beetles in Their Environments; Springer: Berlin, Germany, 1977. [Google Scholar]
- Lövei, G.L.; Sunderland, K.D. Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 1996, 41, 231–256. [Google Scholar] [CrossRef]
- Botes, A.; McGeoch, M.A.; Chown, S.L. Ground-dwelling beetle assemblages in the northern Cape Floristic Region: Patterns, correlates and implications. Austral Ecol. 2007, 32, 210–224. [Google Scholar] [CrossRef]
- Park, Y.; Kim, J.; Jang, T.; Chae, H.; Takami, Y. Local climate mediates spatial and temporal variation in carabid beetle communities in three forests in Mount Odaesan, Korea. Ecol. Entomol. 2017, 42, 184–194. [Google Scholar] [CrossRef]
- Park, Y.; Jang, T.; Jeong, J.; Chae, H.; Kim, J. Local climate mediates spatial and temporal variation in carabid beetle communities on Hyangnobong, Korea. J. For. Environ. Sci. 2017, 33, 161–171. [Google Scholar]
- Park, Y.H.; Won, D.; Jang, T.; Chae, H.; Kim, J.; Kubota, K.; Kim, J. Community structure and distribution of ground beetles (Coleoptera: Carabidae) in Baekdudaegan Mountain, Gangwon-do, Korea. For. Sci. Technol. 2013, 11, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.M. The Agroecology of Carabid Beetles; Intercept Ltd.: Andover, UK, 2002; p. 356. [Google Scholar]
- Kim, J.L.; Lee, C.E. Breeding patterns in southern Korean Carabina (Coleoptera, Carabidae). Proc. Entomol. Soc. Wash. 1992, 94, 136–150. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1963. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q. Variation partitioning by partial redundancy analysis (RDA). Environ. Off. J. Int. Env. Soc. 1997, 8, 75–85. [Google Scholar] [CrossRef]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Luff, M.L. Biology of polyphagous ground beetles in agriculture. Agric. Zool. Rev. 1987, 2, 237–278. [Google Scholar]
- French, B.W.; Elliott, N.C.; Berberet, R.C.; Burd, J.D. Effects of riparian and grassland habitats on ground beetle (Coleoptera: Carabidae) assemblages in adjacent wheat fields. Environ. Entomol. 2001, 30, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Alley, T.R. Competition theory, evolution, and the concept of an ecological niche. Acta Biotheor. 1982, 31, 165–179. [Google Scholar] [CrossRef]
- Welden, C.W.; Slauson, W.L. The intensity of competition versus its importance: An overlooked distinction and some implications. Q. Rev. Biol. 1986, 61, 23–44. [Google Scholar] [CrossRef]
- Keddy, P.A.; Shipley, B. Competitive hierarchies in herbaceous plant communities. Oikos 1989, 54, 234–241. [Google Scholar] [CrossRef]
- Gröning, J.; Hochkirch, A. Reproductive interference between animal species. Q. Rev. Biol. 2008, 83, 257–282. [Google Scholar] [CrossRef] [Green Version]
- den Boer, P.J. Exclusion, competition or coexistence? A question of testing the right hypotheses. J. Zool. Syst. Evol. Res. 1985, 23, 259–274. [Google Scholar] [CrossRef]
- Okuzaki, Y.; Takami, Y.; Sota, T. Resource partitioning or reproductive isolation: The ecological role of body size differences among closely related species in sympatry. J. Anim. Ecol. 2010, 79, 383–392. [Google Scholar] [CrossRef]

| Mountain | Site within a Mountain | Year | Error | |
|---|---|---|---|---|
| Summer mean air temperature | 0.002 | 0.512 *** | 0.094 | 0.392 |
| Summer highest mean air temperature | 0.054 | 0.644 *** | 0.022 | 0.280 |
| Summer lowest mean air temperature | 0.078 | 0.235 | 0.153 | 0.534 |
| Summer mean soil temperature | 0.188 * | 0.145 * | 0.151 | 0.516 |
| Summer mean air humidity | 0.084 | 0.395 ** | 0.219 * | 0.302 |
| Summer mean illumination | 0.216 ** | 0.203 ** | 0.203 | 0.549 |
| Winter mean air temperature | 0.009 | 0.536 *** | 0.274 * | 0.182 |
| Winter highest mean air temperature | 0.038 | 0.670 *** | 0.089 | 0.203 |
| Winter lowest mean air temperature | 0.017 | 0.347 * | 0.288 ** | 0.347 |
| Winter mean soil temperature | 0.062 | 0.281 * | 0.188 | 0.469 |
| Winter mean air humidity | 0.007 | 0.457 ** | 0.114 | 0.422 |
| Winter mean illumination | 0.264 ** | 0.480 *** | 0.062 | 0.194 |
| Total (Pooled for 5 Years) | 2011 | 2012 | 2013 | 2014 | 2015 | Temporal ß | |
|---|---|---|---|---|---|---|---|
| Jinburyung (pooled for three sites) | 1.19 | 0.96 | 1.11 | 1.08 | 1.08 | 1.14 | 0.367M |
| J1 | 1.18 | 0.64 | 1.08 | 0.96 | 0.98 | 1.11 | 0.327 |
| J2 | 1.04 | 0.81 | 0.77 | 0.93 | 0.85 | 0.96 | 0.326 |
| J3 | 1.15 | 0.70 | 1.02 | 0.92 | 1.03 | 1.11 | 0.393 |
| Spatial ß within Jinburyung | 0.239 | 0.318 | 0.352 | 0.340 | 0.293 | 0.327 | - |
| Odaesan (pooled for three sites) | 1.28 | 1.13 | 1.15 | 1.20 | 1.20 | 1.31 | 0.332 |
| O1 | 1.24 | 1.02 | 1.01 | 1.13 | 1.06 | 1.13 | 0.327 |
| O2 | 1.26 | 1.07 | 0.92 | 1.11 | 1.18 | 1.25 | 0.359 |
| O3 | 1.24 | 0.62 | 1.00 | 1.16 | 1.10 | 1.22 | 0.324 |
| Spatial ß within Odaesan | 0.254 | 0.407 | 0.431 | 0.486 | 0.430 | 0.475 | - |
| Taebaeksan (pooled for three sites) | 1.26 | 1.01 | 0.82 | 1.00 | 1.13 | 1.22 | 0.573 |
| T1 | 1.20 | 0.87 | 0.36 | 0.86 | 1.11 | 1.07 | 0.568 |
| T2 | 1.18 | 0.71 | 0.83 | 0.91 | 0.90 | 1.10 | 0.575 |
| T3 | 1.19 | 0.65 | 0.76 | 0.77 | 0.96 | 1.17 | 0.562 |
| Spatial β within Taebaeksan | 0.281 | 0.354 | 0.320 | 0.317 | 0.389 | 0.374 | - |
| Spatial β between mountains | 0.472 | 0.446 | 0.527 | 0.450 | 0.520 | 0.404 | - |
| Mountain | Sites within a Mountain | Year | Error | |
|---|---|---|---|---|
| Diversity | 0.264 ** | 0.051 | 0.373 ** | 0.312 |
| Richness | 0.204 *** | 0.006 | 0.562 *** | 0.229 |
| Abundance | 0.352 *** | 0.062 * | 0.313 ** | 0.273 |
| CCA1 | 0.404 *** | 0.167 *** | 0.048 *** | 0.381 |
| CCA2 | 0.110 | 0.131 | 0.173 | 0.586 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.; Jang, T.; Kim, J.; Kim, S.-K.; Kim, I.-K.; Kim, C.-J.; Takami, Y. Temporal Variation Dominates in Local Carabid Beetle Communities in Korean Mountains. Insects 2021, 12, 1019. https://doi.org/10.3390/insects12111019
Park Y, Jang T, Kim J, Kim S-K, Kim I-K, Kim C-J, Takami Y. Temporal Variation Dominates in Local Carabid Beetle Communities in Korean Mountains. Insects. 2021; 12(11):1019. https://doi.org/10.3390/insects12111019
Chicago/Turabian StylePark, Yonghwan, Taewoong Jang, Jongkuk Kim, Su-Kyung Kim, Il-Kwon Kim, Chang-Jun Kim, and Yasuoki Takami. 2021. "Temporal Variation Dominates in Local Carabid Beetle Communities in Korean Mountains" Insects 12, no. 11: 1019. https://doi.org/10.3390/insects12111019
APA StylePark, Y., Jang, T., Kim, J., Kim, S.-K., Kim, I.-K., Kim, C.-J., & Takami, Y. (2021). Temporal Variation Dominates in Local Carabid Beetle Communities in Korean Mountains. Insects, 12(11), 1019. https://doi.org/10.3390/insects12111019