1. Introduction
The superfamily Elateroidea includes four families with species possessing a defensive “clicking” mechanism based on an advanced promesothoracic locking structure [
1]. The largest of these, the click-beetles (Elateridae), is one of the major beetle radiations with more than 10,000 extant species [
2] and a rich fossil history [
3].
The second largest family, false click-beetles (Eucnemidae), includes more than 1700 extant species [
4]. Their fossil history remained quite obscure until Muona [
5] reviewed the early records and reported 18 genera with 34 species from the Baltic amber. Most of these were described from Eocene samples, but five genera (
Microrhagus Dejean,
Sieglindea Muona,
Erdaia Muona,
Euyptychus LeConte,
Fornax Laporte) were reported both from the Miocene and Eocene.
The first Mesozoic eucnemids were described from lower Cretaceous China [
6] and upper Jurassic Australia [
7]. Otto [
8] and Li et al. [
9] added one species each from the late Cretaceous Burmese amber. Our knowledge of the Mesozoic fauna was greatly expanded when Muona et al. [
10] showed that the previously described Chinese elaterid fossils included one Jurassic and six early Cretaceous eucnemids, all of them belonging to extant monotypic subfamilies (Pseudomeninae Muona, Schizophilinae Muona, and Palaeoxeninae Muona).
Finally, Muona [
11] reported 15 genera and 22 species of eucnemids from the late Cretaceous Burmese amber deposits. Of these, eight genera and 20 species were described as new. One genus had Eocene and Miocene Baltic amber species as well as extant ones (
Euryptychus LeConte), one genus (
Sieglindea Muona) was previously known from Eocene and Miocene Baltic amber, and three genera had extant species (
Epiphanis Eschscholtz,
Jenibuntor Muona,
Myall Muona). In addition to the subfamily Palaeoxeninae, these samples included the first appearance in fossil record of the subfamilies Eucnemiinae Eschscholtz, Melasinae Fleming, and Macraulacinae Fleutiaux—the three dominant Eucnemidae clades today.
Here, we report the first fossil species belonging to the eucnemid subfamily Anischiinae Fleutiaux. The placement of the fossil follows the guidelines presented in Muona et al. [
10].
3. Results
Systematic palaeontology
Class: Insecta Linnaeus, 1758;
Order: Coleoptera Linnaeus, 1758;
Superfamily: Elateroidea Leach, 1815;
Family: Eucnemidae Eschscholtz, 1829;
Subfamily: Anischiinae Fleutiaux, 1936;
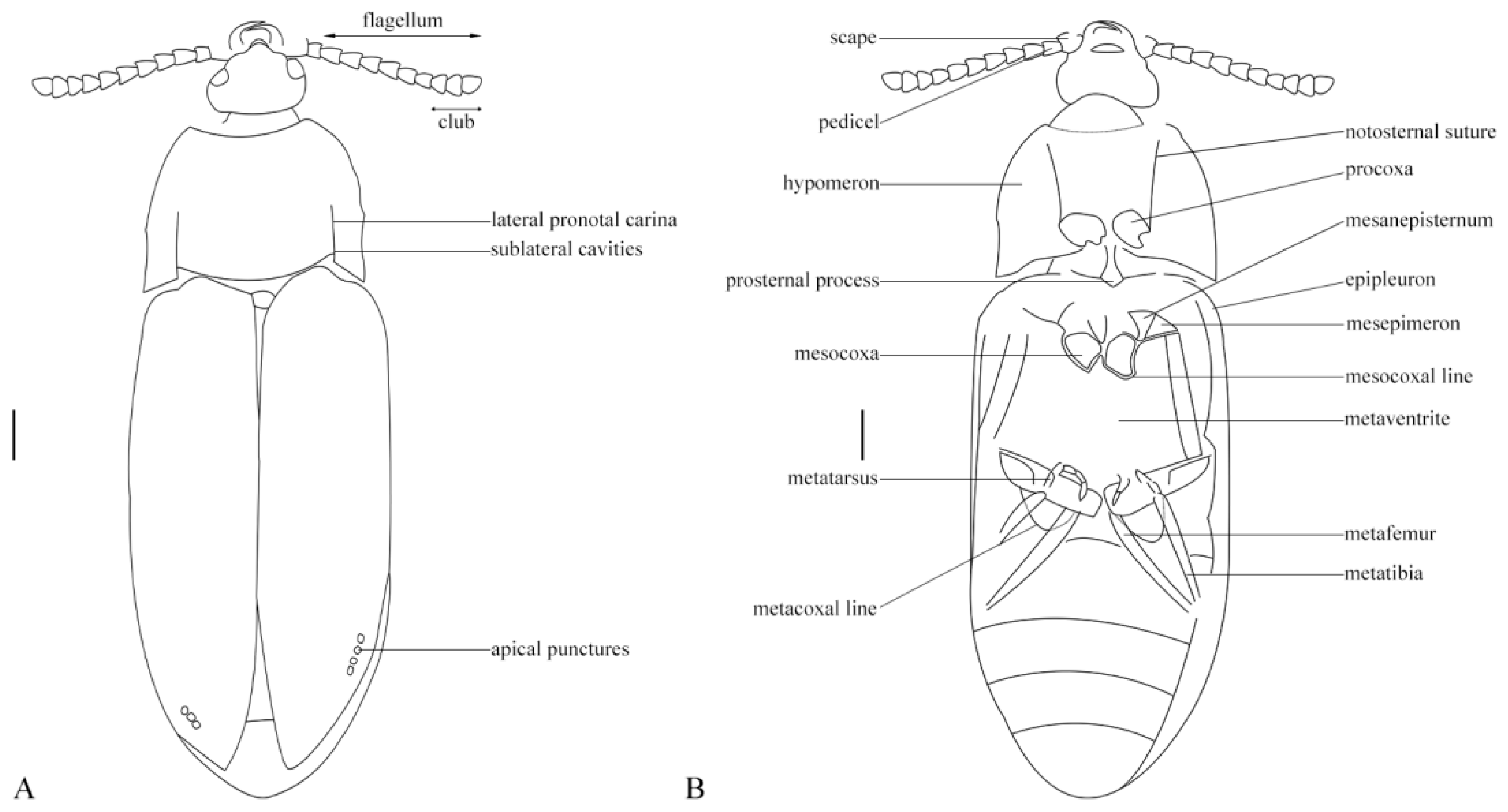
Elateroidea sensu stricto synapomorphy: promesothoracic clicking mechanism present.
Elateroidea
sensu stricto diagnostic other features: form elongated, flattened; metacoxal plates enlarged metaventrite without transverse suture (
Figure 1 and
Figure 2).
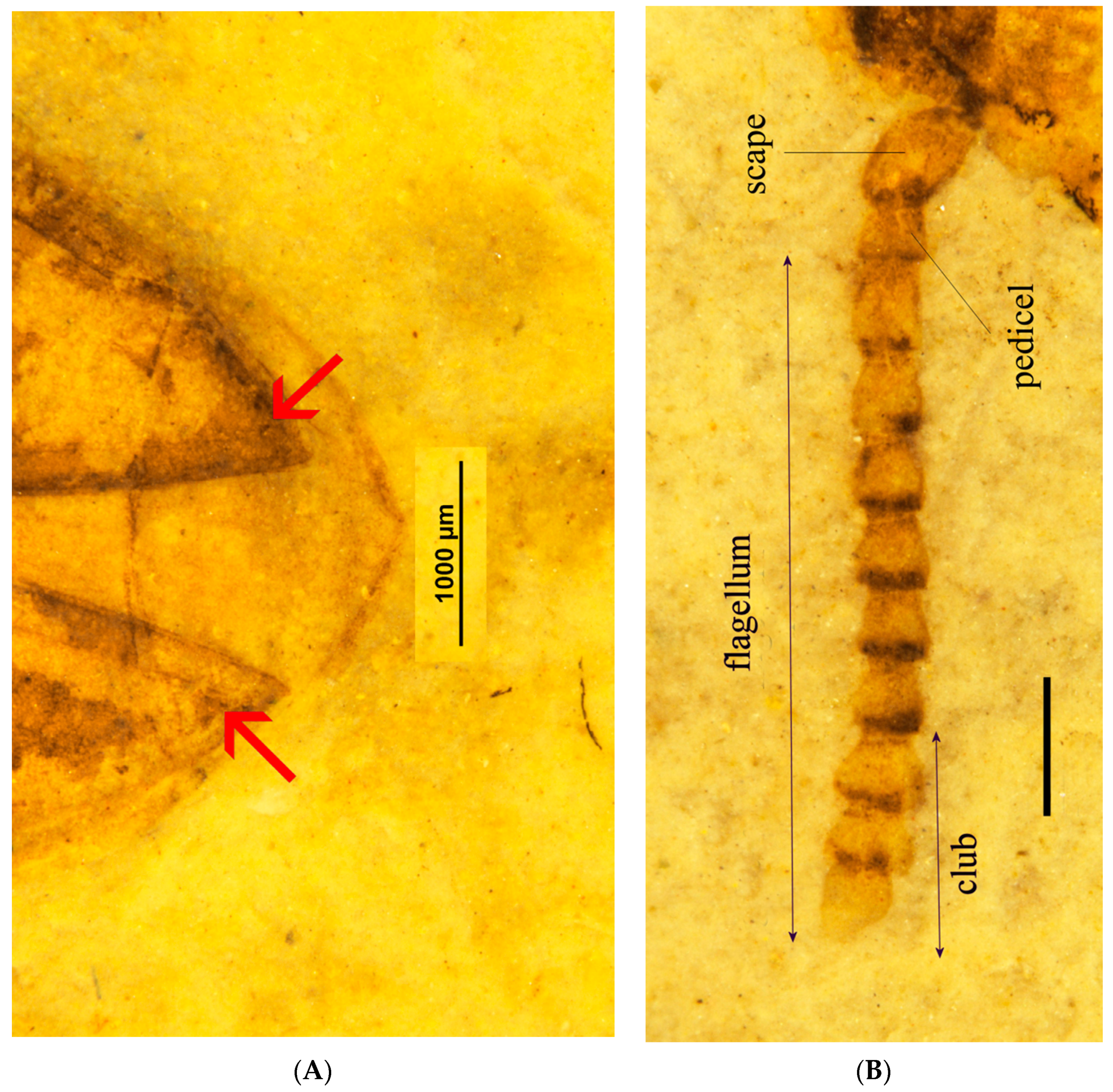
Eucnemidae synapomorphies: elytra with sutural and lateral striae apically with enlarged excretory pores (
Figure 3A); pedicel attached to scape subapically (
Figure 3B).
Eucnemidae other diagnostic feature: antennae with three last flagellomeres enlarged, forming a loose club (
Figure 3B).
Anischiinae synapomorphies: pronotal disc with pair of lateral grooves with carinate edge, extending anteriorly from interlocking cavities at base to middle of disc (
Figure 1,
Figure 2 and
Figure 3); pronotal hind edge with well-developed interlocking device, including pair of deep lateral cavities for receiving paired processes on anterior edges of elytra, anterior edge of each elytron carinate, and produced at middle to form lobe fitting into cavity on pronotum (
Figure 1).
Anischiinae other diagnostic characters: flagellomeres flattened, stout, bulbous in profile, antennae apically abruptly expanded to form weak, three-segmented club (
Figure 3B); labrum attached underneath frontoclypeal region, faintly visible (
Figure 1A and
Figure 3B).
Genus: Rheanischia Li, Chang and Muona new genus.
Type species: Rheanischia brevicornis Li, Chang and Muona new species.
Etymology: a combination of Rhea, one of the female titans in Greek mythology, and Anischia, the type-genus of the subfamily, indicating that species in this genus are much larger than previously known Anischiinae.
3.1. Diagnosis
The promesothoracic clicking mechanism, elytra with apical excretory pores, and the subapically attached pedicel place this genus in Eucnemidae. Within Eucnemidae, it shares the unique pronotal cavities and the interlocking system between pronotum and elytra with the extant genus
Anischia (
Figure 1 and
Figure 4A). The stout clavate antennae and prominent postcoxal lines on abdominal sternite one are further features typical of that genus (
Figure 4B).
Anischia differs from
Rheanischia by being much smaller, having a more rounded body, lacking elytral striae except the sutural one, having prominent vestiture (
Figure 4A) acutely angled coxal lines on first visible ventrite, and having hardly visible metacoxal plates (
Figure 4B).
3.2. Description
Body large, flat, and wide (
Figure 1 and
Figure 2). Antennae stout, relatively short, scape short, flagellomeres robust to moderately slender, seven to nine clavate (
Figure 3B). Head prognathous, wide, subtriangular, mandibles elongated, exposed, fairly large, labrum feebly exposed under the somewhat protruding frontoclypeal region, eyes medium-sized, prominent (
Figure 3B). Pronotum with acute hind angles, slightly diverging craniad, with complete lateral carinae separating disk and hypomera, disk with laterally defined cavities extending over half of the basal length, caudal margin forming interlocking system with base of elytra (
Figure 1 and
Figure 2). Elytra with well-developed sharp sutural striae, other striae distinct but fine, with minute punctures, sutural striae and lateralmost striae apically with excretory punctures (
Figure 1). Hypomera without antennal grooves, meso- and meatathoracic sclerites plesiomorphic, metaventrite wide and extensive, without transverse suture, procoxal cavities nearly circular, moderately spaced, mesocoxal cavities moderately spaced with complete coxal line, metacoxal femoral plates large, subtriangular, strongly narrowing laterad, abdominal sternite one with prominent coxal lines (
Figure 2). Male genitalia exposed in holotype, but vaguely visible, dorsoventreally flattened, appearing to be simply trilobed and having a complete subgenital plate.
Legs long and slender, femora simple, slender, tibiae long, metatibiae with rows of spines, tarsi slender (
Figure 1).
Rheanischia brevicornis Li, Chang and Muona new species.
Etymology: brevicornis refers to stout antennae.
Holotype: male, in Key Laboratory of Insect Evolution and Environmental Changes, College of Life Science, Capital Normal University, Beijing, China, CNU-COL-LB2018344. Dimensions: Body length, 15.28 mm; head width, 1.94 mm; pronotum width, 4.29 mm; length 2.97 mm; elytron length, 9.79 mm; combined elytral width, 5.20 mm.
Paratype: sex unknown, CNU-COL-LB2018313. The specimen has a major part of the abdomen and elytra broken off. Deposited in the same collection as the holotype.
3.3. Diagnosis
Within Anischiinae characterized by the wide, parallel-sided elaterid-like body, short, stout antennae, and widely curved coxal lines on first visible ventrite.
3.4. Description
Antennae stout, scape oval, longer and thicker than flagellomere 1, pedicel slightly longer than wide, length of flagellomere 2, 1.4X its width, 3–6 about equal, longer than wide, subconical, flagellomeres 7–8 more dilated, distinctly wider than long, flagellomere 9 slenderer than 7 or 8, oval, small (
Figure 3B). Pronotal width 1.4X its length, widest at hind angles, very slightly arcuate or straight anteriorly, close to parallel in basal fifth, but narrowing evenly anteriorly. Elytra with distinct, fine striae, interstices convex at apex, all striae visibly punctate, large apical excretory punctures concentrated on sutural striae and lateral ones.
4. Discussion
The most striking external feature in extant
Anischia species is the structure of the pronotum. The interlocking system between pronotum and elytra as well as the deep, laterally defined grooves on pronotal disk are unique within Eucnemidae. Similar structures are found in Elateridae: Cardiophorinae and Negastriinae [
22], but these taxa lack synapomorphies defining Eucnemidae: pedicel attached subapically to scape, labrum attached underneath the fronotclypeal region, and larval legs extremely reduced. Cardiophorinae and Negastrinae share an apomorphy not found in
Anischia, female bursa with a structure called colleterial glands—a putative synapomorphy for some or all Elateridae [
22], but see also [
23]. The actual nature of these gland-like features is unclear, however.
Douglas (pers. comm.) has pointed out to us that some Cardiophorinae elaterids have enlarged punctures apically on elytra, a feature characterizing the majority of Eucnemidae. Douglas [
23] coded this character (his number 90) as missing for an unidentified Central American
Anischia species. We have checked several
Anischia species and have been able to verify that
Anischia mexicana Fleutiaux (Mexico),
Anischia boliviana Fleutiaux (Bolivia),
Anischia ruandana (Basilewsky) (Ivory Coast), and several undescribed species from Panama, Colombia, Brazil, and Peru have large excretory punctures apically, located in longitudinal sutural depressions, but
Anischia bicolor Lawrence (New Caledonia),
Anischia kuscheli Lawrence (New Caledonia),
Anischia stupenda Fleutiaux (New Guinea), and four undescribed species from Vanuatu, Fiji, and the Philippines have at most the shallow sutural apical depressions on elytra. This may show a further evolutionary development of this character. Whether this reflects the presence of two separate clades remains to be seen.
The overall appearance of the large extinct
Rheanischia species (
Figure 1) is strikingly different from that of the extant
Anischia species (
Figure 4A). The systematic position of
Anischia in Eucnemidae was firmly established with a total evidence approach only quite recently [
24]. This result was confirmed in an analysis with molecular sampling as well [
25]. The discovery of
Rheanischia shows that the seemingly odd eucnemid,
Anischia, is actually a derived member of an ancient clade present in early Cretaceous, today surviving as a small pantropical, morphologically modified group.
Excluding the Anischiinae synapomorphies from consideration for a moment,
Rheanischia becomes a fairly “normal” basal eucnemid. The abdomen appears to be connate; body form is flattened and elaterid-like; antennae have three enlarged apical flagellomeres; elytra have striae; metacoxal plates are large and strongly narrowing laterad; metatibiae have spine-combs and the male aedeagus appears to be trilobed, dorsoventrally flattened. All these features suggest a position close to
Palaeoxenus Horn, another extant eucnemid clade known from the Yixian fauna as well [
6]. This relationship is basically what Lawrence et al. suggested [
24]. Muona and Teräväinen [
26], however, were not able to find a placement for
Anischia within Eucnemidae on the basis of its unusual larva. The discovery of Anischiinae fossils with a set of plesiomorphic elateroid features combined with the Anischiinae synapomorphies offered support for the earlier hypothesis [
24], and clarified the exact position of Anischiinae within Eucnemidae. The polymorphism of apical elytral excretory punctures in extant species is exceptional, but as stated in Muona and Teräväinen [
10], this feature has been lost secondarily in some derived Macraulacini as well.
The
Anischia larval structures, including large endodont mandibles, head without lateral teeth, and segmented legs, should be accepted as plesiomorphic features, and the whole “derived eucnemid clade” (“DE” in [
26]) should be regarded as a sister-group to it. The informal group DE consists of Palaeoxeninae and its sister group, all other derived lignicolous eucnemids, i.e., Eucneminae, Macraulacinae, Melasinae and Phlegoninae. The larval structures are the key synapomorphies uniting those beetles: exodont mandibles, head capsule with highly developed lateral structures, and total loss of segmented legs [
26]. The evolutionary sequence for the lignicolous Eucnemidae
sensu [
26], including Anischiinae is thus: (Pseudomeninae) ((Schizophilinae) ((Anischiinae ((Palaeoxeninae) ((Phlegoninae) (Melasinae, Eucneminae, Macraulacinae)))))).
If the present host choices of these clades are seen as reflecting the original ones, this hypothesis means that the eucnemids invaded either angiosperms or gymnosperms multiple times. The larva of Pseudomenes bakewelli (Bonvouloir) has been collected from “rotten log” (Blundell’s Creek, Brindabella Mts., ACT, Australia) but unfortunately without more precise data. Black cypress pine (Callitris edlicheri) could occur at the site, but many species of Eucalyptus are also possible candidates as to the host. The host tree remains unknown.
Schizophilus subrufus (Randall) breeds in angiosperms [
27], as does
Anischia [
24].
Palaeoxenus dohrnii (Horn) breeds in gymnosperms (6, Otto, in litt.; Muona, pers. obs.). The host for Phlegoninae species is not known.
Chang et al. [
6] and Muona et al. [
10] showed that basal lignicolous Eucnemidae diversification had taken place in the Jurassic and the Cretaceous. Here, we demonstrated that Anischiinae were present in the Cretaceous as well. Thus, both Palaeoxeninae breeding in gymnosperms and Anischiinae breeding in angiosperms belong to clades older than the main angiosperm radiation. This dating strongly supports the hypothesis that lignicolous eucnemids first bred in gymnosperms and invaded angiosperms later on.
Anischia larvae with endodont mandibles would thus represent an adaptive radiation from gymnosperms to angiosperms without major cephalic modifications, whereas
Palaeoxenus larvae, although breeding in gymnosperms, shares the highly typical head modifications found in more than a thousand extant species developing in angiosperms [
26]. This scenario indicates a problem of more general nature. Care should be taken when extrapolating from the structural specializations of modern insect larvae their way of life in distant past. On the other hand, insect larvae developing in wood infested by fungi may succeed with simple-looking morphological structures just well as with complex ones.
The feeding methods used by these beetles may have simplified these evolutionary switches. Eucnemidae use extra-oral digestion, feeding on the bracket-fungus hyphae in the infected wood. In the few cases studied, the fungus species appears to be the deciding factor, not the tree species [
5,
26] and Muona, unpublished). As many bracket-fungi utilize both angiosperms and gymnosperms as hosts, the potential for a beetle to switch over from one tree species to another one is clear.
Anischiinae and Paleoxeninae represent two early lineages of Eucnemidae. They are separate clades, however, as both groups have solid synapomorphies defining them. Both groups diversified before the main angiosperm radiation in Late Jurassic–Early Cretaceous, but their subsequent histories are very different. Palaeoxeninae, still developing in conifers, survive only in the Southern Californian Mountains [
6], whereas Anischiinae, utilizing fungi growing on angiosperms, are found in the tropics of South America, Africa, Asia, and the Pacific [
24,
28,
29]. Eucnemidae clades have been able to find worldwide success utilizing fungi attacking angiosperms, whereas sticking to the coniferous route has turned out to be an evolutionary dead-end.



{kind=link}
{kind=link}
{kind=link}
{kind=link}



