
Species Composition of Thrips (Thysanoptera: Thripidae) in Strawberry High Tunnels in Denmark


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of Thrips in Strawberry Tunnels
2.2. Fast-Slide-Preparation of Adults
2.3. Morphological Identification of Adults
2.4. Molecular Identification of Two Species of Larvae
2.5. DNA Extraction
2.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR) and Output
2.7. Statistical Analysis
3. Results
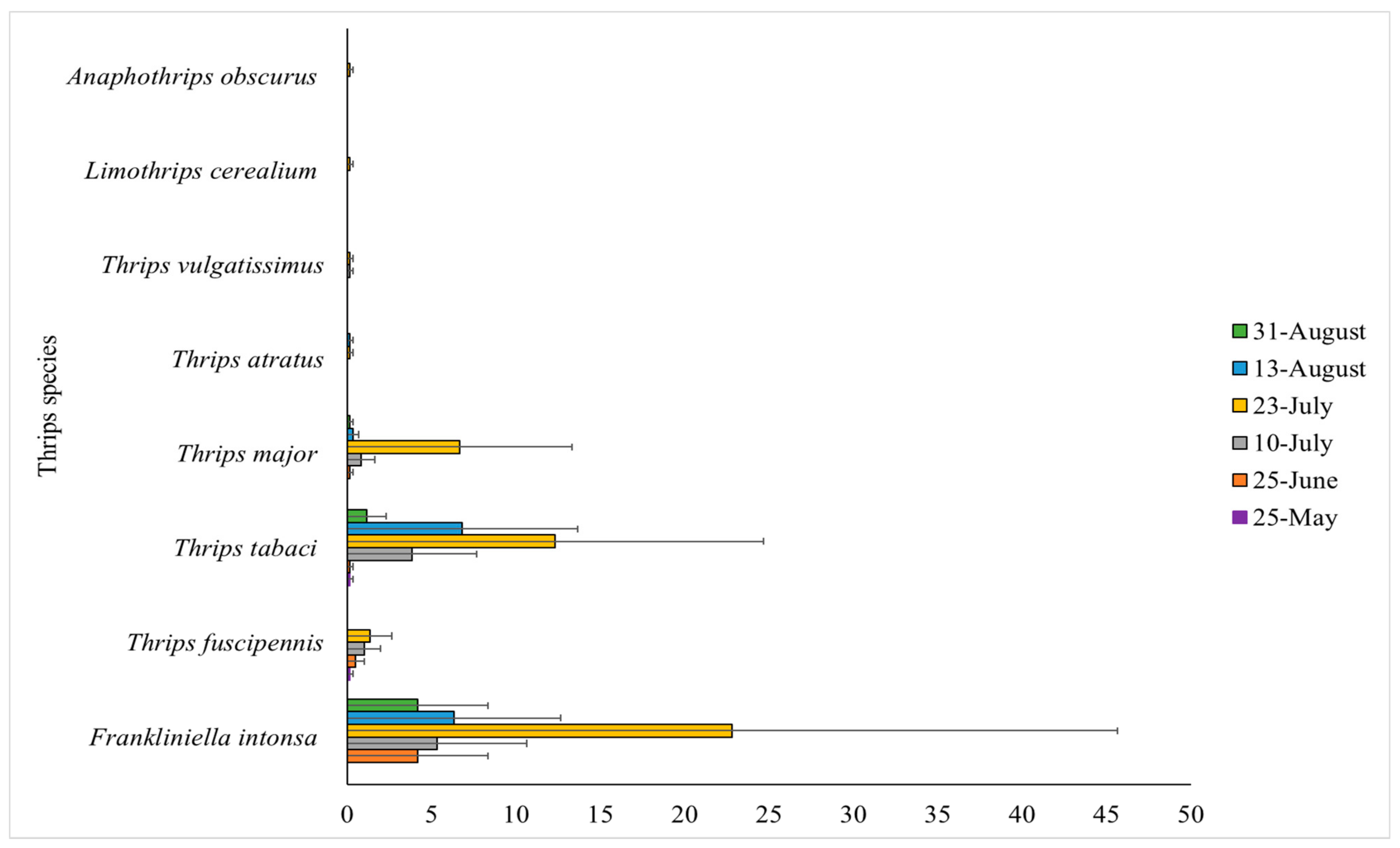
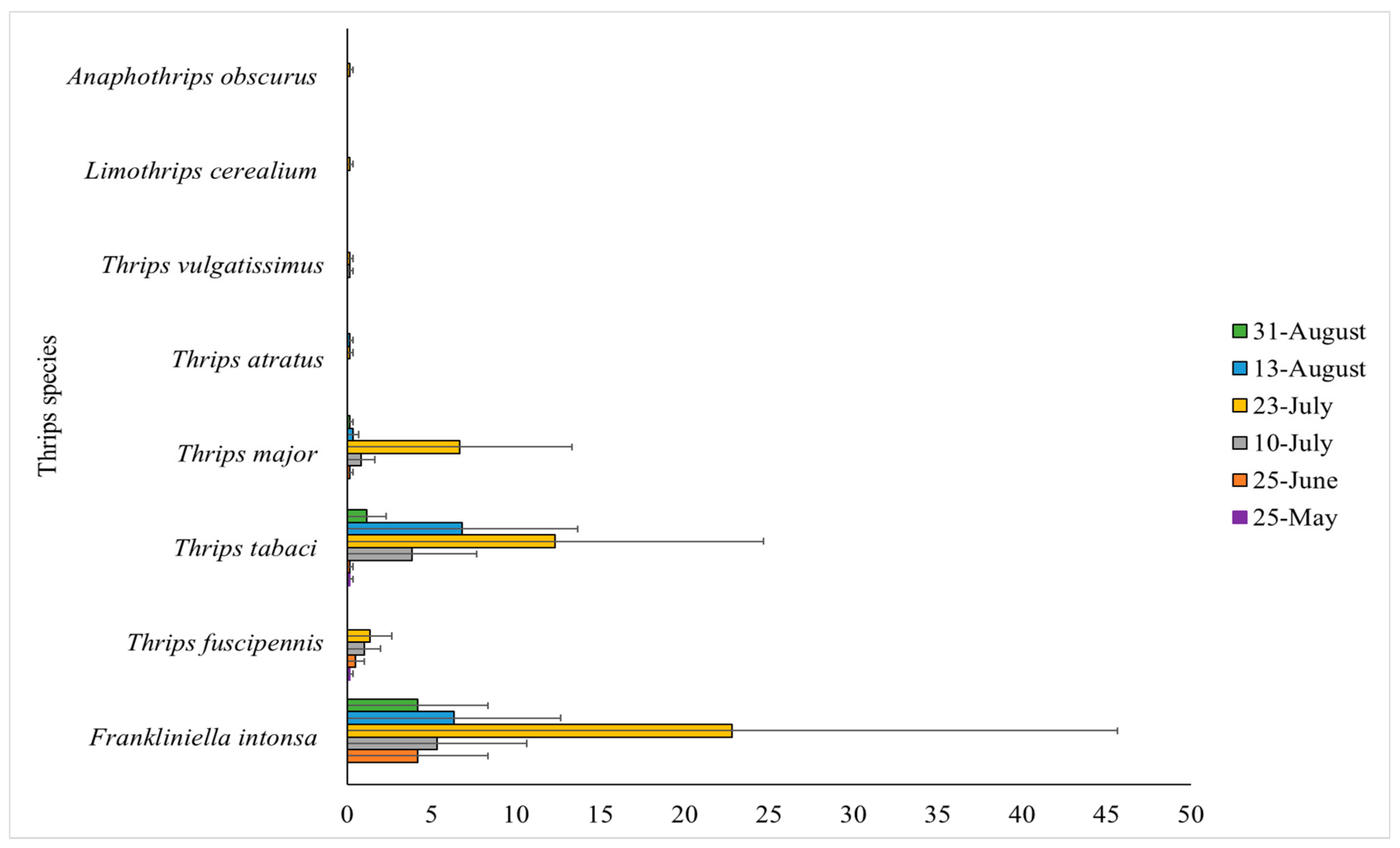
3.1. Species Composition of Thrips Adults and Larvae during the Production Season
3.2. Species Found at Other Locations in Denmark
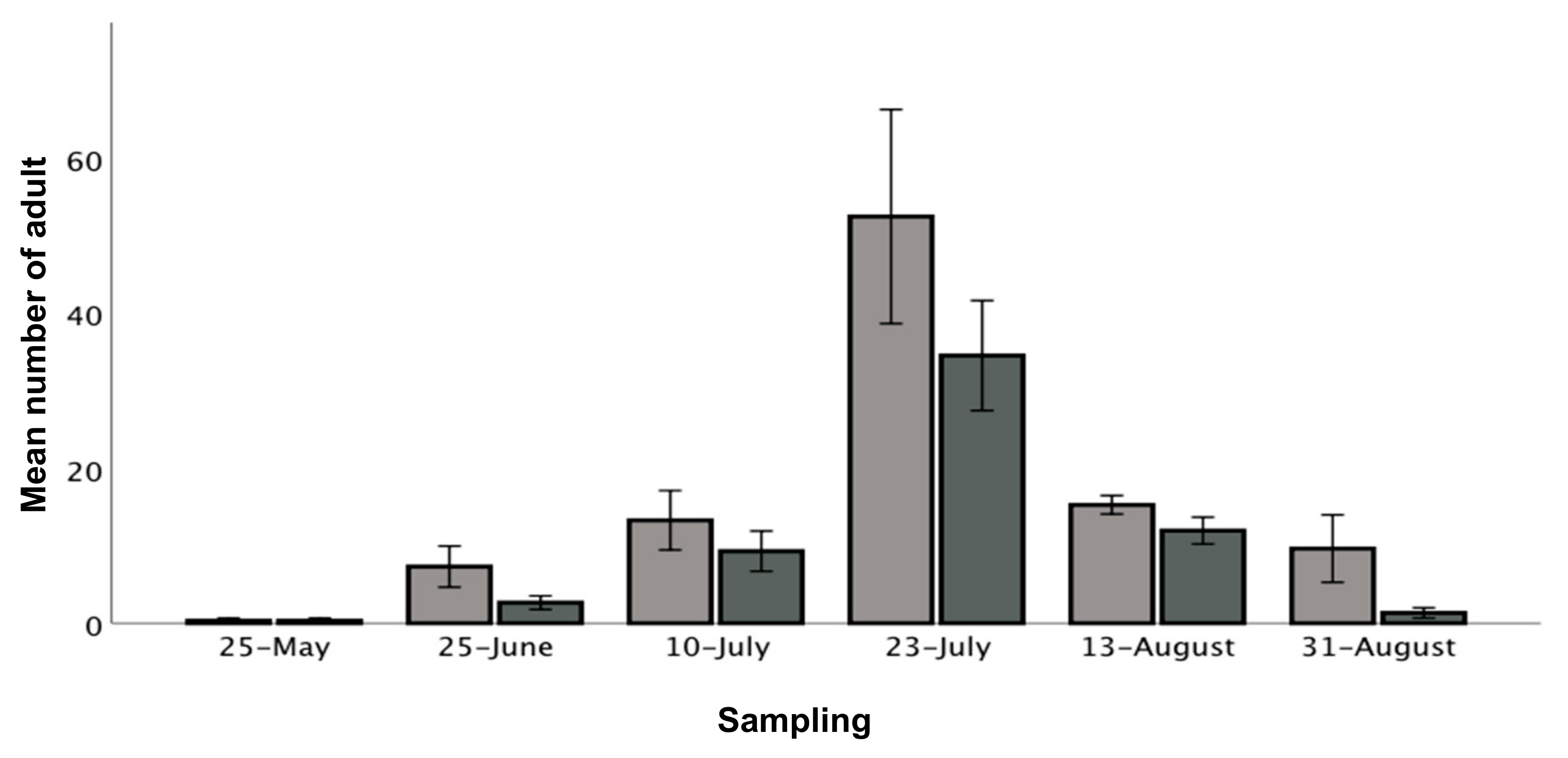
3.3. Effect of Cultivar, Time and Location on the Overall Density of Adult Thrips
4. Discussion
4.1. Species Composition and Distribution of Thrips
4.2. Difference in Abundance of Frankliniella Intonsa and Thrips Tabaci
4.3. Temporal and Spatial Patterns of Adult Thrips
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, T. Thrips as Crop Pests, 1st ed.; CAB International: New York, NY, USA, 1997; pp. 11–12. [Google Scholar]
- Lewis, T. Thrips: Their Biology, Ecology and Economic Importance, 1st ed.; Academic Press: London, UK, 1973; p. 9. [Google Scholar]
- Parker, B.L.; Skinner, M.; Lewis, T. Thrips Biology and Management, 1st ed.; Springer International Publishing: New York, NY, USA, 1995; pp. 19–20. [Google Scholar]
- Mound, L. Natural and disrupted patterns of geographical distribution in Thysanoptera. J. Biogeogr. 1983, 10, 119–133. [Google Scholar] [CrossRef]
- Mound, L.; Walker, A. A Fauna of New Zealand, 1st ed.; Science Information Division: Wellington, New Zealand, 1982; p. 12. [Google Scholar]
- McDonald, J.R.; Bale, J.S.; Walters, K.F.A. Effect of temperature on development of the western flower thrips. Eur. J. Entomol. 1998, 95, 301–306. [Google Scholar]
- Stacey, D.; Fellowes, M.D.E. Temperature and the development rates of thrips: Evidence for a constraint on local adaptation? Eur. J. Entomol. 2002, 99, 399–404. [Google Scholar] [CrossRef] [Green Version]
- Danmarks Statestik: Det Dyrkede Areal Efter Afgrøde, Enhed, Område, og Tid. Available online: https://www.statistikbanken.dk/statbank5a/SelectVarVal/saveselections.asp (accessed on 5 June 2019).
- HortiAdvice IPM Dyrkningsvejledning for Jordbær. Report; HortiAdvice: Odense, Denmark, 2016; pp. 1–6. [Google Scholar]
- Sønsteby, A.; Karhu, S. Strawberry production, growth and development in northern climates. Int. J. Fruit Sci. 2005, 5, 107–114. [Google Scholar] [CrossRef]
- Daugaard, H. Tabletop produktion af jordbær. Det Jordbrugsvidenskabelige Fak. Århus Univ. 2007, 175, 1–6. [Google Scholar]
- Raffle, S.; Irving, R.; Moore, G. Extending the UK Strawberry Season Using a Range of Plant Types and Growing Systems; Factsheet from Horticultural Development Company: Warwickshire, UK, 2010; pp. 1–9. [Google Scholar]
- Atakan, E.; Pehlivan, S.; Kiminsu, A. Pest status of Western Flower Thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), in tunnel-grown strawberry. Turk. Entomoloji Derg. 2016, 40, 61–71. [Google Scholar] [CrossRef]
- Alford, D. Agricultural Entomology, 1st ed.; Blackwell Science: Cambridge, UK, 1999; pp. 90–93. [Google Scholar]
- Steiner, M.Y.; Goodwin, S. Management of thrips (Thysanoptera:Thripidae) in Australian strawberry crops: Within-plant distribution characteristics and action thresholds. Aust. J. Entomol. 2005, 44, 175–185. [Google Scholar] [CrossRef]
- Sampson, C. Management of pesticide-resistant western flower thrips on tunnel- grown strawberry: A study of the reasons for successes and failures on commercial production sites. Rep. AHDB 2014, 1, 1–22. [Google Scholar]
- Coll, M.; Shakya, S.; Shouster, I.; Steinberg, Y. Decision-making tools for Frankliniella occidentalis management in strawberry: Consideration of target markets. Entomol. Exp. Appl. 2007, 122, 59–67. [Google Scholar] [CrossRef]
- Sampson, C.; Kirk, W.D.J. Predatory mites double the economic injury level of Frankliniella occidentalis in strawberry. BioControl 2016, 61, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Sampson, C. Sustainable management of the Western Flower Thrips in strawberry crops. Out. Pest Man 2018, 10, 1–6. [Google Scholar] [CrossRef]
- Jensen, N.L.; Berry consultant at HortiAdvice, Odense, Denmark. Personal Communication, 2021.
- Cross, J. Integrated Control of Thrips on Strawberries; Factsheet from Horticultural Development Company: Kent, CT, USA, 2003; pp. 1–8. [Google Scholar]
- Mound, L.A.; Morison, G.D.; Pitkin, B.R.; Palmer, J.M. Handbook for the identification of British insect: Thysanoptera, 1st ed.; Royal Entomological Society of London: London, UK, 1976; pp. 14–18, 29–32, 46, 56. [Google Scholar]
- Kobro, S. Checklist of Nordic Thysanoptera. Nor. J. Entomol. 2011, 58, 20–26. [Google Scholar]
- Kirk, W.D.J.; Terry, I. The spread of the Western Flower Thrips Frankliniella occidentalis (Pergande). Agric. For. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of Western Flower Thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S. Insecticide resistance in Western Flower Thrips Frankliniella occidentalis. Integr. Pest Manag. Rev. 2000, 5, 131–146. [Google Scholar] [CrossRef]
- Molnár, A.; Pap, Z.; Fail, J. Observing population changes of thrips (Thysanoptera) species damaging forced pepper and their natural enemies. Int. J. Hortic. Sci. 2008, 14, 55–60. [Google Scholar] [CrossRef]
- Shipp, J.L.; Whitfield, G.H. Functional response of the predatory mite, Amblyseius cucumeris (Acari: Phytoseiidae), on Western Flower Thrips, Frankliniella occidentalis (Thysanoptera: Thripidae). Environ. Entomol. 1991, 20, 694–699. [Google Scholar] [CrossRef]
- Shipp, J.L.; Wang, K.; Binns, M.R. Economic injury levels for Western Flower Thrips (Thysanoptera: Thripidae) on greenhouse cucumber. J. Econ. Entomol. 2009, 93, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.S.; Lim, U.T. Life history characteristics of Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae) in constant and fluctuating temperatures. J. Econ. Entomol. 2015, 108, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennison, J.; Hough, G. Maintaining the Expertise for Developing and Communicating Practical Integrated Pest Management (IPM) Solution for Horticulture. Annual Report from AHDB. 2015, pp. 1–44. Available online: https://ahdb.org.uk/cp-089-maintaining-the-expertise-for-developing-and-communicating-practical-integrated-pest-management-ipm-solutions-for-horticulture-emt-hdc-hta-fellowship (accessed on 18 January 2021).
- Li, W.D.; Zhang, P.J.; Zhang, J.M.; Zhang, Z.J.; Huang, F.; Bei, Y.W.; Lin, W.C.; Lu, Y.B. An evaluation of Frankliniella occidentalis (Thysanoptera: Thripidae) and Frankliniella intonsa (Thysanoptera: Thripidae) performance on different plant leaves based on life history characteristics. J. Insect Sci. 2015, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Van Rijn, P.C.J.; Mollema, C.; Steenhuis-Broers, G.M. Comparative life history studies of Frankliniella occidentalis and Thrips tabaci (Thysanoptera: Thripidae) on cucumber. Bull. Entomol. Res. 1995, 85, 285–297. [Google Scholar] [CrossRef]
- Deligeorgidis, P.; Ipsilandis, C.; Vaiopoulou, M.; Deligeorgidis, N.; Stavridis, D.; Sidiropoulos, G. The competitive relation between Frankliniella occidentalis and Thrips tabaci: The impact on life-cycle and longevity. J. Entomol. 2006, 3, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Bhuyain, M.M.H.; Lim, U.T. Interference and exploitation competition between Frankliniella occidentalis and F. intonsa (Thysanoptera: Thripidae) in laboratory assays. Fla. Entomol. 2019, 102, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Helyer, N.L.; Brobyn, P.J. Chemical control of Western Flower Thrips (Frankliniella occidentalis Pergande). Ann. Appl. Biol. 1992, 121, 219–231. [Google Scholar] [CrossRef]
- Riudavets, J. Predators of Frankliniella occidentalis and Thrips tabaci: A review. Wagening. Agric. Univ. Pap. 1995, 95, 46–78. [Google Scholar]
- MacIntyre Allen, J.K.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Resistance of Thrips tabaci to pyrethroid and organophosphorus insecticides in Ontario, Canada. Pest Manag. Sci. 2005, 61, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Frescata, C.; Mexia, A. Biological control of thrips (Thysanoptera) by Orius laevigatus (Heteroptera: Anthocoridae) in organically-grown strawberries. Biol. Agric. Hortic. 1996, 13, 141–148. [Google Scholar] [CrossRef]
- Xu, X.; Borgemeister, C.; Poehling, H.M. Interactions in the biological control of Western Flower Thrips Frankliniella occidentalis (Pergande) and Two-Spotted Spider Mite Tetranychus urticae Koch by the predatory bug Orius insidiosus say on beans. Biol. Control 2006, 36, 57–64. [Google Scholar] [CrossRef]
- Messelink, G.J.; Van Steenpaal, S.E.F.; Ramakers, P.M.J. Evaluation of phytoseiid predators for control of Western Flower Thrips on greenhouse cucumber. BioControl 2006, 51, 753–768. [Google Scholar] [CrossRef]
- Messelink, G.J.; van Maanen, R.; van Steenpaal, S.E.F.; Janssen, A. Biological control of thrips and whiteflies by a shared predator: Two pests are better than one. Biol. Control 2008, 44, 372–379. [Google Scholar] [CrossRef] [Green Version]
- Mehle, N.; Trdan, S. Traditional and modern methods for the identification of thrips (Thysanoptera) species. J. Pest Sci. 2012, 85, 179–190. [Google Scholar] [CrossRef]
- Silveira, L.C.P.; Haro, M.M. Fast slide preparation for thrips (Thysanoptera) routine identifications. Eur. J. Entomol. 2016, 113, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Walter, D.; Krantz, G. Collectiong, rearing, and preparing specimens. In A Manual of Acarology, 2nd ed.; Texas Tech University: Texas, TX, USA, 2009; pp. 83–96. [Google Scholar]
- Mound, L.; Collins, D.; Hastings, A. Thysanoptera Britannica et Hibernica-Thrips of the British Isles. Available online: https://keys.lucidcentral.org/keys/v3/british_thrips/the_key/britishthysanoptera_2017.html (accessed on 1 February 2019).
- Trdan, S.; Andjus, L.; Raspudić, E.; Kač, M. Distribution of Aeolothrips intermedius Bagnall (Thysanoptera: Aeolothripidae) and its potential prey Thysanoptera species on different cultivated host plants. J. Pest Sci. 2005, 78, 217–226. [Google Scholar] [CrossRef]
- Stopar, K.; Trdan, S.; Bartol, T. Thrips and natural enemies through text data mining and visualization. Plant Prot. Sci. 2020, 57, 47–58. [Google Scholar] [CrossRef]
- Brunner, P.; Flemming, C.; Frey, J. A molecular identification key for economically important thrips species (Thysanoptera: Thripidae) using direct sequencing and a PCR-RFLP-based approach. Agric. For. Entomol. 2002, 4, 127–136. [Google Scholar] [CrossRef]
- Yeh, W.B.; Tseng, M.J.; Chang, N.T.; Wu, S.Y.; Tsai, Y.S. Agronomically important thrips: Development of species-specific primers in multiplex PCR and microarray assay using internal transcribed spacer 1 (ITS1) sequences for identification. Bull. Entomol. Res. 2015, 105, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Single-species detection. In Environmental DNA: For Biodiversity Research and Monitoring, 1st ed.; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Tuovinen, T.; Lindqvist, I. Effect of introductions of a predator complex on spider mites and thrips in a tunnel and an open field of pesticide-free everbearer strawberry. J. Berry Res. 2014, 4, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Lim, U.T.; Mainali, B.P. Optimum density of chrysanthemum flower model traps to reduce infestations of Frankliniella intonsa (Thysanoptera: Thripidae) on greenhouse strawberry. Crop Prot. 2009, 28, 1098–1100. [Google Scholar] [CrossRef]
- Mintu, C.; Reyes, C.P. Frankliniella intonsa (Trybom), a thrips species infesting strawberry in La Trinidad, Benguet Province, Philippines. Asia Life Sci. 2018, 27, 377–383. [Google Scholar]
- Atakan, E. Population densities and distributions of the Western Flower Thrips (Thysanoptera: Thripidae) and its predatory bug, Orius niger (Hemiptera: Anthocoridae), in strawberry. Int. J. Agric. Biol. 2011, 13, 638–644. [Google Scholar]
- Upton, M. Aqueous gum-chloral slide mounting media: A historical review. Bull. Entomol. Res. 1993, 83, 267–274. [Google Scholar] [CrossRef]
- Kobayashi, T. Regeneration of old slide specimens mounted in Hoyer’s medium. Chronomus J. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Raspudić, E.; Ivezić, M.; Brmež, M.; Trdan, S. Distribution of Thysanoptera species and their host plants in Croatia. Acta Agric. Slov. 2009, 93, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Atakan, E.; Uygur, S. Winter and spring abundance of Frankliniella spp. and Thrips tabaci Lindeman (Thysan., Thripidae) on weed host plants in Turkey. J. Appl. Entomol. 2005, 129, 17–26. [Google Scholar] [CrossRef]
- Orosz, S. Investigation of Thysanoptera Populations in Sweet Pepper Greenhouses and in Their Surroundings. Ph.D. Thesis, Szent István University, Budapest, Hungary, 2012. [Google Scholar]
- Brown, S.; Bennison, J.; Boxworth, A. Determining the Threat of Rose Thrips (Thrips fuscipennis) in UK Strawberry Crops. Report from AHDB. 2018, pp. 1–28. Available online: https://www.linkedin.com/authwall?trk=gf&trkInfo=AQEOlTEmRQybDwAAAXfxku0Qt6RYm8eE85v7Bzr98x83Z_JKEoqS1t83RqWztl7JJHQ4vaIrF_fUwiu05J_jz_m2CgWlcZ8bazzLYGHfrVRpgtm4VMwcgARyTciF8za-BIYcEx0=&originalReferer=&sessionRedirect=https%3A%2F%2Fuk.linkedin.com%2Fpublic-profile%2Fin%2Fsam-brown-1873ab12b%3FchallengeId%3DAQG-Ny0GZ4yZzQAAAXfxMn20efHLtHctJGaX11eEXByRYMwtpGsZCiEa7iz4englsrE4DsF2r-b-S1KqBfYOnNEM0oR39rhsEg%26submissionId%3D89eaffa1-206c-6816-8465-ff2634451651 (accessed on 18 January 2021).
- Sampson, C.; Kirk, W.D.J. Can mass trapping reduce thrips damage and is it economically viable? Management of the Western Flower Thrips in strawberry. PLoS ONE 2013, 8, 238–245. [Google Scholar] [CrossRef] [Green Version]
- Sampson, C. Management of the Western Flower Thrips on Strawberry. Ph.D. Thesis, Keele University, Keele, UK, 2014. [Google Scholar]
- Brødsgaard, H. Cold hardiness and tolerance to submergence in water in Frankliniella occidentalis. Environ. Entomol. 1993, 22, 647–653. [Google Scholar] [CrossRef]
- Factsheet from Norsk Landbruksrådgivning; Jordbærsorter: Ås, Norway, 2016; pp. 1–12.
- Crawley, R.; Pattrasudhi, R. Interspecific competition between insect herbivores: Asymmetric competition between Cinnabar Moth and the Ragwort Seed-Head Fly. Ecolo. Entomol. 1988, 13, 243–249. [Google Scholar] [CrossRef]
- Denno, R.F.; McClure, M.S.; Ott, J.R. Interspecific interactions in phytophagous insects-competiton reexamined and resurrected. Ann. Rev. Entomol. 1995, 40, 297–331. [Google Scholar] [CrossRef]
- Sampson, C.; Kirk, W. Flower stage and position affect population estimates of the Western Flower Thrips, Frankliniella occidentalis (Pergande), in strawberry. Acta Phytopathol. Entomol. Hung. 2012, 47, 133–139. [Google Scholar] [CrossRef]
- Leis, M.; Martinelli, A. Plant Patent on the Strawberry Cultivar Murano. U.S. Patent US PP25,070 P3, 1–4 November 2014. [Google Scholar]
- Rahman, T.; Spafford, H.; Broughton, S. Variation in preference and performance of Frankliniella occidentalis (Thysanoptera: Thripidae) on three strawberry cultivars. J. Econ. Entomol. 2010, 103, 1744–1753. [Google Scholar] [CrossRef]
- Rahman, T.; Broughton, S.; Spafford, H. Effect of spinosad and predatory mites on control of Frankliniella occidentalis in three strawberry cultivars. Entomol. Exp. Appl. 2011, 138, 154–161. [Google Scholar] [CrossRef]
- Ayalew, G.; Sciarretta, A.; Baumgärtner, J.; Ogol, C.; Löhr, B. Spatial distribution of Diamondback Moth, Plutella xylostella L. (Lepidoptera: Plutellidae), at the field and the regional level in Ethiopia. Int. J. Pest Manag. 2008, 54, 31–38. [Google Scholar] [CrossRef]
- Labou, B.; Brévault, T.; Sylla, S.; DIatte, M.; Bordat, D.; DIarra, K. Spatial and temporal incidence of insect pests in farmers’ cabbage fields in Senegal. Int. J. Trop. Insect Sci. 2017, 37, 225–233. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Species | June | July | August | Total | Percentage of Larvae | p-Values |
|---|---|---|---|---|---|---|
| Frankliniella intonsa | 36 | 34 | 17 | 87 | 60% | June and July p > 0.05 |
| June and August p < 0.001 | ||||||
| August and July p < 0.001 | ||||||
| Thrips tabaci | 0 | 8 | 20 | 28 | 20% | June and July p > 0.05 |
| June and August p < 0.001 | ||||||
| August and July p < 0.001 | ||||||
| Other | 8 | 11 | 11 | 30 | 20% | No difference between months (p > 0.05) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, H.; Sigsgaard, L.; Kobro, S.; Jensen, N.L.; Jacobsen, S.K. Species Composition of Thrips (Thysanoptera: Thripidae) in Strawberry High Tunnels in Denmark. Insects 2021, 12, 208. https://doi.org/10.3390/insects12030208
Nielsen H, Sigsgaard L, Kobro S, Jensen NL, Jacobsen SK. Species Composition of Thrips (Thysanoptera: Thripidae) in Strawberry High Tunnels in Denmark. Insects. 2021; 12(3):208. https://doi.org/10.3390/insects12030208
Chicago/Turabian StyleNielsen, Helene, Lene Sigsgaard, Sverre Kobro, Nauja L. Jensen, and Stine K. Jacobsen. 2021. "Species Composition of Thrips (Thysanoptera: Thripidae) in Strawberry High Tunnels in Denmark" Insects 12, no. 3: 208. https://doi.org/10.3390/insects12030208
APA StyleNielsen, H., Sigsgaard, L., Kobro, S., Jensen, N. L., & Jacobsen, S. K. (2021). Species Composition of Thrips (Thysanoptera: Thripidae) in Strawberry High Tunnels in Denmark. Insects, 12(3), 208. https://doi.org/10.3390/insects12030208