Simple Summary
Endemic Collembola species are bioindicators of environmental quality since native species abundance is particularly sensitive to environmental disturbances. Oceanic island biota generally present high percentages of endemic species, and the vulnerability of these species is higher than those of the continents. The objective of this work was to carry out a survey of the Collembola species of the order Poduromorpha in the Brazilian oceanic islands and synthesize a distribution list of this order for Brazil. Our results reveal four new species of Collembola Poduromorpha for Brazilian oceanic islands that may be useful for the conservation strategies of these island regions and a contributor to the knowledge of the order in Brazil.
Abstract
We present new species and records of Poduromorpha for the Brazilian oceanic islands and synthesis of this order in Brazil. Friesea noronhaensis sp. nov., Friesea rochedoensis sp. nov., Willemia insularum sp. nov. and Paraxenylla zeliae sp. nov. are described and a diagnosis of the morphospecies Acherontiella sp. Lima and Zeppelini 2015 is provided. We present comparative tables, distribution and taxonomic keys of the Friesea, Arlesia, Brachystomella, Acherontiella, Paraxenylla, Xenylla, and Willemia found in the Brazilian oceanic islands and their respective congeners recorded in Brazil.
1. Introduction
The order Poduromorpha Börner, 1913 is represented by Collembola with an elongated body, thorax with three separate segments and prothorax with chaetae. Members of this order, about 3400 species, occur in all zoogeographic regions [1].
Abrantes et al. [2] summarized a list of Brazilian Collembola with 102 species of Poduromorpha and 2 morphospecies (Mesogastrura cf. ojcoviensis and Mesaphorura sp. gr. atlantica), none of them recorded in Brazilian oceanic islands.
Oceanic islands generally have a high percentage of endemic species. However, in Brazil, the only survey of the Collembola fauna in oceanic islands was done by Lima and Zeppelini in 2015 [3], who presented 36 new records of Collembola, among them 4 species and 3 morphospecies of the Poduromorpha order: Friesea cubensis Potapov and Banasko, 1985; Brachystomella agrosa Wray, 1953; Xenylla yucatana Mills, 1938; Isotogastrura mucrospatulata Palacios-Vargas, Lima and Zeppelini, 2013; Acherontiella sp.; Arlesia sp. and Willemia sp., all from Fernando de Noronha Archipelago.
We present results of recent surveys of the Archipelago of São Pedro e São Paulo, Atol das Rocas (Rocas Atoll) and the taxonomic advances for Fernando de Noronha Archipelago after Lima and Zeppelini [3]. Friesea noronhaensis sp. nov., Friesea rochedoensis sp. nov., Willemia insularum sp. nov. and Paraxenylla zeliae sp. nov are described. We provide comparative tables, distribution, remarks and taxonomic keys to Brazilian species of the genera Friesea, Arlesia, Brachystomella, Acherontiela, Paraxenylla, Xenylla, Willemia, Isotogastrura, as well as an update of the distribution and habitat synthesis of the Brazilian Poduromorpha, including contributions after Abrantes et al. [2].
2. Materials and Methods
2.1. Poduromorpha Survey of the Brazilian Oceanic Islands
This survey was conducted in 3 Brazilian oceanic islands (2 Archipelagos and 1 Atoll) located in the Equatorial Atlantic Ocean, all belonging to the Brazilian national territory, as follows:
Fernando de Noronha Archipelago—AFN (3°50′ S, 32°15′ O), São Pedro e São Paulo Archipelago—ASPSP (0°55′ N, 29°20′ O) and Atol das Rocas (Rocas Atoll)—AT (3°51′ S, 33°48′ O) about 360, 1010 and 267 km from the American continent, respectively (Figure 1).
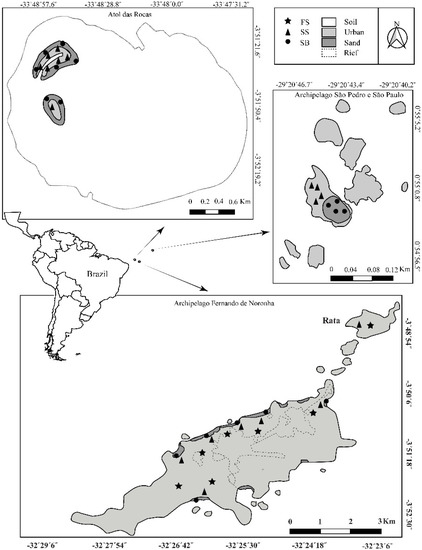
Figure 1.
Brazilian oceanic island of the Equatorial Atlantic Ocean. Map show distribution of sites sampled in Fernando de Noronha archipelago, Atol das Rocas (Rocas Atoll) and São Pedro e São Paulo Archipelago.
2.2. Sample Procedure
The specimens were collected in three coastal environmental areas: the intertidal zone in sand or rock areas, without vegetation, referred to as sandy beach (SB); the sloping land closest to the intertidal zone, with vegetation, referred to as slope forest (SF) and the forest more distant from the beach, referred to as top forest (TF), this latest area only exists in Fernando de Noronha.
Specimens were preserved in ethanol, cleared in Nesbitt’s fluid and then mounted on glass slides in Hoyer’s medium [4]. Figures were elaborated in computer graphic software, photographs and measurements were obtained using a Zeiss Axio Scope A1 microscope with Axiocam 105 color, and Zen 2 Blue software, and deposited in the Coleção de Referência de Fauna de Solo at the Universidade Estadual da Paraíba, João Pessoa, PB, Brazil.
2.3. Taxonomic Procedure
The nomenclature of furcal development stages follows Cassagnau 1958. Antennal S-chaetae are numbered from S1 to S8 [5]. Dorsal chaetotaxy nomenclature follows the chaetal group system [6,7,8].
2.4. Species List of Brazilian Poduromorpha
Species records, collection location, habitat and biotope information for each species were modified from [2], included: newly described species, new records published between 2012 and 2019 and a survey of the Brazilian oceanic islands between 2012 and 2017. Publications that do not provide identifications to continental species were omitted except genus and morpho-species from Brazilian oceanic islands. Information on the world distribution of species was based on [9] and original new records. Biogeographical distribution regions according to [10], modified by [9,11] as follows: Boreal (Bor) include regions 1–8, Neotropical (Neo) regions 24–30, South African (SAf) region 31, Paleotropical (Pal) regions 9–23, Australian (Aus) regions 32–34, and Antarctic (Ant) regions 35–37. Species restricted to Brazil were based on Culik and Zeppelini [9], defining the Brazilian biogeographic subregions: Amazon (Amz), North and Central Brazil (NCB) and Pampa (Pam) corresponding to biogeographic regions 26, 27 and 29, respectively [9]. Species distributed, at least, in four of the major regions (Neo, Pal, etc.) are considered cosmopolitan (Cos).
2.5. Abbreviations
Abd—abdominal segment, Ant—antennal segment, Cd—cephalic diagonal, PAO—postantennal organ, Oc—ocular, ta—anterior trichobothria, Th—thoracic segment, Tita—tibiotarsus, tm—medial trichobothria, tl—lateral trichobothria, tp—posterior trichobothria, Di—dorsointernal group of chaetae, De—dorsoexternal group of chaetae, DL—dorsolateral group of chaetae, CRFS-UEPB—Coleção de Referência da Fauna de Solo-Universidade Estadual da Paraíba, IUCN—International Union for Conservation of Nature. Brazilian states abbreviations according to Table 1.

Table 1.
Abbreviations of names of the Brazilian states and oceanic islands. Number of Poduromopha species recorded and region location of each state and oceanic island. Brazilian continental region designations: N—north, NE—northeast, CW—west (central part), SE—southeast, S—south. Brazilian insular region: EA—equatorial Atlantic; SA—South Atlantic. Map shows the geographic position of Brazilian states and oceanic islands.
3. Results
The Brazilian Poduromorpha fauna is recorded in the following states and oceanic islands (Table 1).
3.1. Poduromorpha Survey of the Brazilian Equatorial Oceanic Islands
The Poduromorpha fauna of the Brazilian oceanic islands is represented by 9 species, as follows:
3.1.1. Neanuridae Börner, 1901
Friesea noronhaensis sp. nov. Lima and Zeppelini
Type material. Holotype male on the slide, Brazil, Fernando de Noronha Archipelago, Rata Island, (3°48′45.61″ S; 32°23′26.17″ O), SF, 19.vii.2012, E.C.A. Lima and A.S. Ferreira leg., deposited at the CRFS-UEPB # 14574. Paratypes: 2 females and 2 males on slides, same data as holotype, deposited at the CRFS-UEPB # 14572, 14573, 15060, 1563.
Description. Habitus typical of the genus. Color in ethanol dark gray. Body with secondary integuments. Body length average: 512 µm (n = 5); holotype measurements in Table 2.
Table 2.
Holotype measurements for Friesea noronhaensis sp. nov., Friesea rochedoensis sp. nov., Willemia insularum sp. nov., Acherontiella sp. and Paraxenylla zeliae sp. nov. All measurements in µm.
Antenna. Antenna shorter than cephalic diagonal (Table 2). Ant IV with 6 S-chaetae (S1–4, S7–8), 1 small ventrolateral S-microchaeta and 1 small subapical organite. Apical bulb simple; with 5 dorsal clavate chaetae and about 12 smooth chaetae ventrally. Sensory organ of Ant III consisting of 2 small subcylindrical, bent, internal S-chaetae covered by a fold of the integument, 2 subcylindrical guard S-chaetae Sgd and Sgv, and 1 ventral S-microchaeta. Ant II with 12 chaetae; Ant I with 7 chaetae (Figure 2a,b).
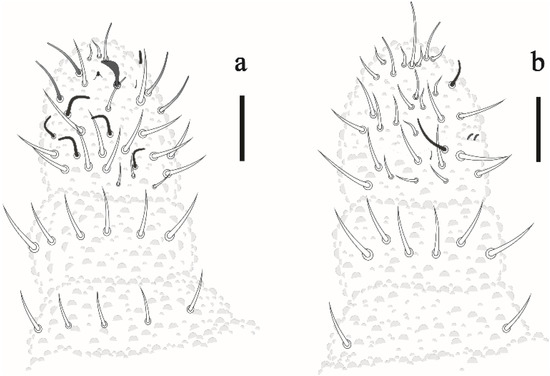
Figure 2.
Friesea noronhaensis sp. nov. (a) dorsal view of antenna (b) ventral view of antenna. Scale bar: 10 μm.
Head. Eyes 8 + 8 in dark ocular plate; PAO absent; chaetae a0, d0 and 3 Oc present (Figure 3). Pre-labral/labral formula: 4/3,3,4. Mandible with seven teeth, three apical, a medial and three basal (Figure 4a). Maxilla typical of the genus (Figure 4b). Labium with papillated chaeta L (Figure 4c).
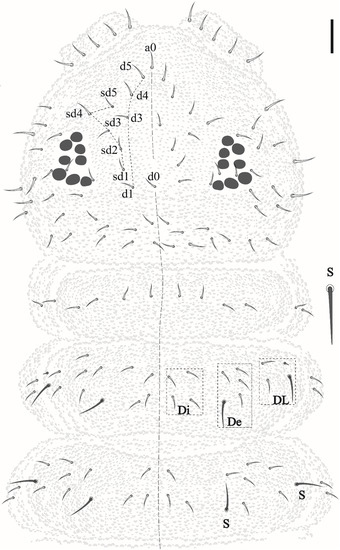
Figure 3.
Friesea noronhaensis sp. nov, dorsal view of head and Th I–III. Scale bar 10 μm.
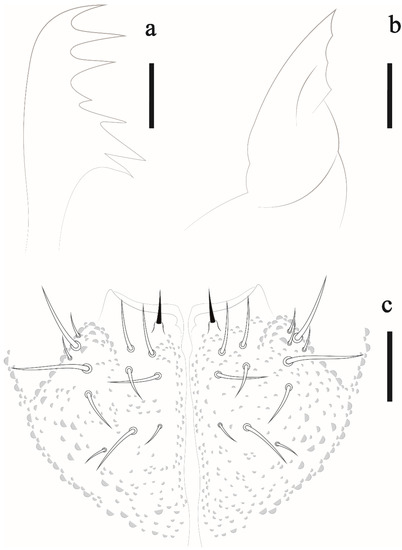
Figure 4.
Friesea noronhaensis sp. nov. (a) mandible, scale bar: 5 μm. (b) maxilla, scale bar: 5 μm. (c) labium, scale bar: 10 μm.
Head and body chaetotaxy composed of slightly rugged acuminate ordinary chaetae. S-chaetae smooth, except on Th II–III in DL that are roughly blunt. S-chaetae measuring about twice the size of anterior ordinary chaetae in Th II–III and Abd I–IV. Abd V–VI with posterior strong rough, blunt chaetae longer than others of the body. Sensillar formula by half tergum: 022/11111 (Figure 3 and Figure 5).
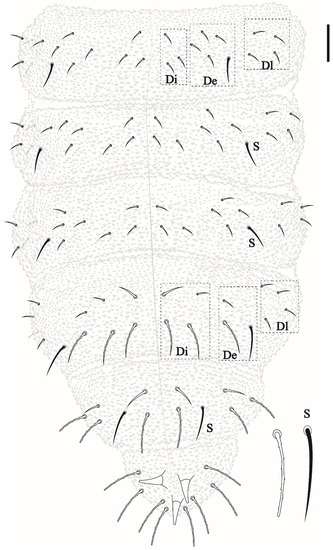
Figure 5.
Friesea noronhaensis sp. nov. 8, dorsal view of abdominal segments I–VI. Scale bar 10 μm.
Thoracic chaetotaxy. Th I: 4 + 4; Th II: 13 + 13 (Di: 4 chaetae; De: 4 chaetae + 1 S-chaetae; Dl: 2 chateae, 1 S-chaetae +1 ms); Th III: 11 + 11 (Di: 3 chaetae; De: 4 chaetae +1 S-chaetae; Dl: 2 chaetae +1 S-chaetae) see Figure 3.
Abdominal chaetotaxy. Abd I–III: 12 + 12 (Di: 3; De: 4 + 1 S-chaetae; Dl: 4). Abd IV: 11 + 11 (Di: 4; De: 2 + 1 S-chaetae; Dl: 4). Abd V: 6 + 6 (3 anterior acuminate chaetae and 3 posterior blunt chaetae measuring about twice the size than anterior plus 1 + 1 S-chaetae in p2) chaetae. Abd VI with 8 clavate chaetae and 3 anal spines (Figure 5), 2 in a1 position and the third as MO.
Appendages. Tita I–III: 18, 18, 17 chateae, with 3, 4, 4 longer and clavate tenent hair, respectively (Figure 6a); Femora I–III: 10, 14, 14 chaetae; trochanters I–III: 5, 5, 5 chaetae; coxae I–III: 3, 7, 7 chaetae; subcoxae I–III with 0, 2, 2 chaetae; epicoxae I–III with 1, 2, 2. Unguis without teeth. Ventral tube with 4 + 4 chaetae. Tenaculum with 2 + 2 teeth and furca state 1 (Figure 6b). Genital plate of female and male with 9 and 28 chaetae, respectively. Anal valves with 16 + 16 chaetae, each anal valve with 2 hr chaetae (Figure 6c).

Figure 6.
Friesea noronhaensis sp. nov. (a) Tita III, scale bar: 10 μm. (b) furca, scale bar: 10 μm. (c) ventral view of abdominal segment II–VI, scale bar: 20 μm.
Etymology. Species name derives from the type locality, popularly known as Noronha.
Remarks. Friesea noronhaensis sp. nov. is similar to F. cubensis Potapov and Banasco, 1985; F. furculata Deharveng and Bedos, 1991; F. rubeni Deharveng and Bedos 1991 and F. australica Greenslade and Deharveng, 1997 by developed furca, dens with 3 chaetae and separate mucro. Friesea cubensis and the new species presents 3 anal spines, while F. furculata, F. rubeni and F. australica feature 0, 2 and 4, respectively. The new species differs from F. cubensis by posterior chaetae blunt versus acuminate in Abd V–VI and tenent hair clavate versus acuminate. For morphological comparisons between Friesea species recorded in Brazil, see Table 3.
Table 3.
Morphological characters of Friesea species record from Brazil. S Abd VI—number of the spine on Abd VI; M Abd VI—modified chaetae on Abd VI; Ten h—tenent hair; BD—body length in μm; AP—apical vesicle; TL—type locality; “?”—missing date; number in Furca column refers of furca “State” according to [12].
Distribution. Friesea noronhaensis sp. nov. is registered only in the Fernando de Noronha Archipelago on Fernando de Noronha Island (SF sites) and Rata Island (SF and TF sites). The specimens were collected in superficial soil and leaf.
Friesea rochedoensis sp. nov. Lima and Zeppelini
Type material. Holotype female on slide, Brazil, São Pedro e São Paulo Archipelago, Belmonte Island, (0°54′59.98″ N; 29°20′44.03″ O), guano over rocks, May 2017, E.C.A. Lima and D. Zeppelini leg., deposited at the CRFS-UEPB # 14571. Paratypes: 2 males on slides, Brazil, São Pedro e São Paulo Archipelago, Belmonte Island, (0°54′59.74″ N; 29°20′44.57″ O), guano over rocks, 24 April 2017, E.C.A. Lima and D. Zeppelini leg., deposited at the CRFS-UEPB # 14569, 14570.
Description. Habitus typical of the genus. Light blue color in ethanol. Body with well-developed secondary integuments. Body length average: 995 µm (n = 5); holotype measurements as in Table 2.
Antenna. Antenna shorter than cephalic diagonal (Table 2). Ant IV dorsally with 6 S-chaetae (S1–4, S7–8), 1 small ventrolateral S-microchaeta and 1 small subapical organite; apical bulb simple ventrally; with 3–4 dorsoapical and 10–12 clavate plus 5 smooth ventroapical chaetae. Sensory organ of Ant III consisting of 2 small subcylindrical, bent, internal S-chaetae covered by a fold of the integument, 2 subcylindrical guard S-chaetae Sgd and Sgv, and 1 ventral S-microchaeta. (Figure 7a,b). Ant I with 7 chaetae; Ant II with 12 chaetae.
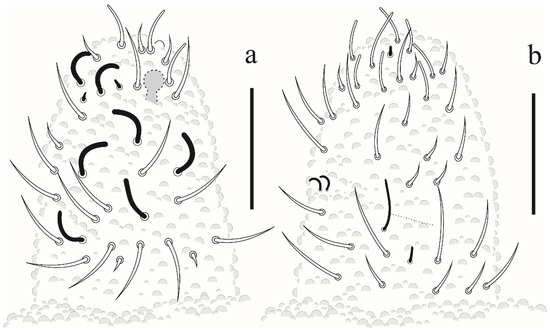
Figure 7.
Friesea rochedoensis sp. nov. (a) dorsal view of antenna, scale bar: 20 μm. (b) ventral view of antenna, scale bar: 20 μm.
Head (Figure 8a). Eyes 8 + 8 in dark ocular plate; PAO absent; chaetae a0, d0 and 3 Oc present. Pre-labral/labral formula: 4/3,3,4. Mandible with seven teeth, three apical, a medial and three basal (Figure 8b). Maxilla typical of the genus. Labium with papillated chaeta L.
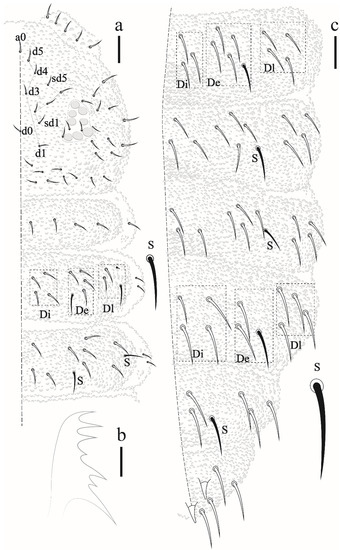
Figure 8.
Friesea rochedoensis sp. nov. (a) dorsal view of head and Th I–III, scale bar: 20 μm. (b) mandible, scale bar: 20 μm. (c) dorsal view of abdominal segments I–VI, scale bar: 20 μm.
Head and body chaetotaxy composed of slightly rough acuminate ordinary chaetae and S-chaetae about 1/3 longer than ordinary chaetae in Th I–II. Abd I–V S-chaetae subequal or slightly longer than ordinary posterior chaetae. Sensillar formula by half tergum: 022/11111.
Thoracic chaetotaxy (Figure 8a). Th I: 4 + 4; Th II: 13 + 13 (Di: 4 chaetae; De: 4 chaetae + 1 S-chaetae; Dl: 2 chaetae, 1 S-chaetae + 1 ms); Th III: 11 + 11 chaetae (Di: 3 chaetae; De: 4 chaetae +1 S-chaetae; Dl: 2 chaetae +1 S-chaetae).
Abdominal chaetotaxy (Figure 8c). Abd I–IV: 12 + 12 (Di 3; De 5; Dl 4, posterior chaetae about 1/4 longer than size anterior chaetae). Abd V: 6 + 6 (3 anterior chaetae and 3 posterior of which one S-chaetae subequal chaetae). Abd VI with 6 chaetae and 3 anal spines.
Appendages. Tita I–III: 18, 18, 17 chaetae, with 3, 4, 4 longer and clavate tenent hairs, respectively; Femora I–III: 10, 11, 10 chaetae; trochanters I–III: 5, 5, 5 chaetae; coxae I–III: 3, 7, 7 chaetae; subcoxae I–III with 0, 2, 2 chaetae; epicoxae I–III: 0, 2, 2. Unguis short, without tooth on inner edge. Ventral tube with 4 + 4 chateae. Tenaculum with 2 + 2 teeth and furca type 1. Genital plate of female (Figure 9a) and male with 17–19 and 26–28 chaetae (Figure 9b). Anal valves with 16 + 16 and 2 hr chaetae (Figure 9c).
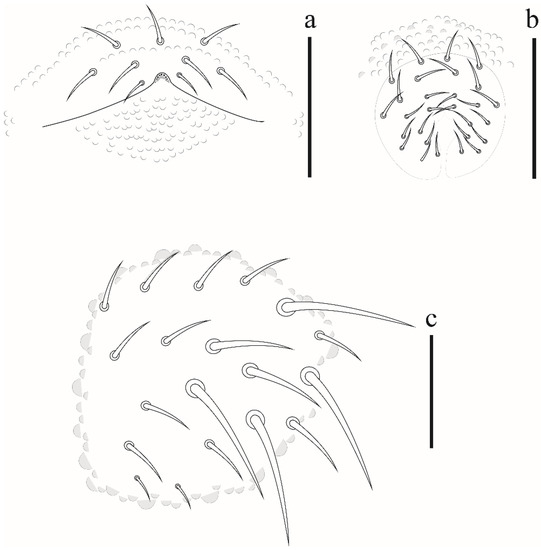
Figure 9.
Friesea rochedoensis sp. nov. (a) female genital plate, scale bar: 20 μm. (b) male genital plate, scale bar: 20 μm. (c) left anal valve, scale bar: 20 μm.
Etymology. Species name derives from the old name of the type locality that was previously known as “Rochedo de São Pedro e São Paulo”.
Remarks. F. rochedoensis sp. nov. is similar to F. noronhaensis sp. nov. by chaetotaxy, eyes number, mandible, furca type, number of anal spines and clavate tenent hair. Differs from F. noronhaensis by color (light blue vs. dark gray), for absent posterior blunt clavate chaetae in Abd V–VI, by the size (body length 995 vs. 512 µm) and a number of clavate chaetae dorsal and ventroapical of the Ant IV (3–4, 10–12 vs. 5, 0). Friesea rochedoensis sp. nov. has hardly differentiated S-chaetae (especially on Abd V) from ordinary chaetae, whereas F. noronhaensis sp. nov. and F. cubensis S-chaetae are easily differentiated from ordinary chaetae. Friesea rochedoensis sp. nov. has chaetae a1 and p1 subequal in Abd V, while F. noronhaensis sp. nov and F. cubensis have p1 greater than a1 in Abd V (Figure 10a–c). For others, morphological comparisons between Friesea species recorded in Brazil, see Table 3.
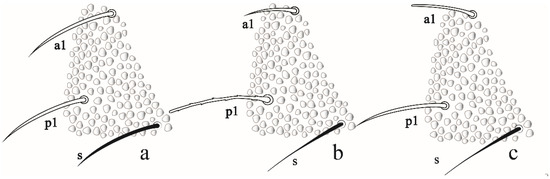
Figure 10.
Comparasion between a1, p1 and S-chaetae of abdominal segment V in: (a) Friesea rochedoensis sp. nov. (b) Friesea noronhaensis sp. nov. (c) Friesea cubensis (modified from [6]).
Distribution. Friesia rochedoensis sp. nov. is registered only for Belmont island of the São Pedro e São Paulo Archipelago. The specimens were collected from seabird guano deposits over rocks. For the congeners registered in Brazil, see Figure 11.
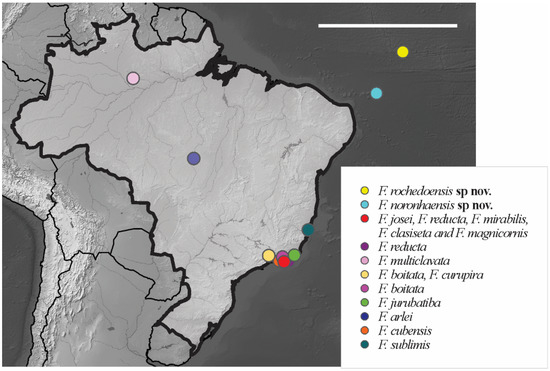
Figure 11.
Distribution map for Brazilian species of the genus Friesea Dalla Torre, 1895. Scale bar: 3000 km.
Identification Key for the Species of Friesea Recorded in Brazil
1 Without furca...................................................................................................................... 2
– With furca............................................................................................................................ 7
2 1 + 1 eyes................................................................ F. curupira Queiroz and Mendonça, 2015
– With more than 1 + 1 eyes.................................................................................................... 3
3 Modified chaetae of Abd VI are spiniform...................... F. josei Palacios-Vargas, 1986
– Modified chaetae of Abd VI are clavate.......................................................................... 4
4 With 10 clavate chaetae on the abd VI............................................................................. 5
– Less than 10 clavate chaetae on Abd VI.......................................................................... 6
5 Th I with 3 + 3 chaetae....................................................................... F. reducta Denis, 1931
– Th I with 2 + 2 chaetae....................... F. multiclavata Neves, Mendonça and Queiroz, 2019
6 With 6 slightly clavate chaetae on abd VI........... F. boitata Queiroz and Mendonça, 2015
– With 8 clavate chaetae on Abd VI................... F. jurubatiba Silveira and Mendonça, 2018
7 4 + 4 eyes........................................................................ F. arlei Massoud and Bellinger, 1963
– 8 + 8 eyes................................................................................................................................ 8
8 Developed furca (state 1 according to [12])..................................................................... 9
– Reduced furca (state 2 according to [12])........................................................................ 11
9 Modified chaetae of Abd VI clavate and rugged, clavate tenent hair........................................................................................................... F. noronhaensis sp. nov.
– Modified chaetae of the Abd VI acuminate................................................................... 10
10 Tenent hair acuminate......................................... F. cubensis Potapov and Banasko, 1985
– Tenent hair clavate......................................................................... F. rochedoensis sp. nov.
11 With clavate chaetae on Abd VI.................................................................................... 12
– Without clavate chaetae on Abd VI..................................... F. mirabilis (Tullberg, 1871)
12 Ant IV with 5 sensilla..................................................................................................... 13
– Ant IV with 6 sensilla................................................................ F. claviseta Axelson, 1900
13 Anal spine are the same size............................................. F. sublimis Macnamara, 1921
– Posterior anal spine smaller than anterior............................ F. magnicornis Denis, 1931
Arlesia albipes (Folsom, 1927)
Brazilian Oceanic Island Records. FN (3°52′8.88″ S; 32°26′13.57″ W), SF, 01 August 2012, E.C.A. Lima, A.S. Ferreira leg., CRFS# 15144–15145.
Brazilian Occurrence. In Brazil, this species is widely distributed in the states PA, PE, PB, PI, MG and RJ [13] (Figure 12). In Brazilian oceanic island Arlesia albipes recorded only in Fernando de Noronha Archipelago on Fernando de Noronha Island (SF sites). The specimens were collected on superficial soil and leaf litter.
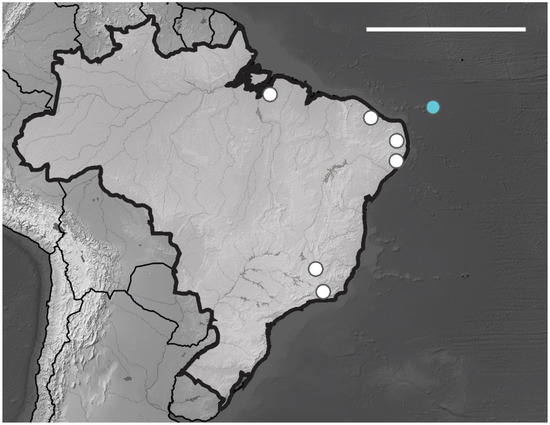
Figure 12.
Distribution map for Arlesia albipes (Folsom, 1927) records in Brazil. Blue circle, Fernando de Noronha; white circle, Brazilian states. Scale bar: 3000 km.
Biogeographic Distribution. Type locality of Arlesia albipes is Margarita Swamp, Canal Zone, collected by J. Zetek and I. Molino on June 28, 1923, in a termite mound near the base of a tree stump and deposited at U.S.N.M. # 40383. According to [1], this species has been found in the following biogeographic regions: Caribbean mainland (24a), Antillean and South Florida (24b), Venezuela and Guyana (25), Amazon (26), Northeast and Central Brazilian (27) and Andean (28). Neotropical species.
Remarks. The specimens found in the Fernando de Noronha Archipelago are in accordance with the original description [14] and updates [15,16]. Lima and Zeppelini [3] recorded as A. sp. nov. For comparisons between Arlesia species recorded in Brazil, see Table 4.
Table 4.
Morphological characters of Arlesia Handschin, 1942 species recorded from Brazil. BD—body length in mm; AP—apical bulb vesicles; SF—sensory formula; MT—mandibular teeth; TL—type locality.
Identification Key for the Species of Arlesia Handschin, 1942 Recorded in Brazil
1 Mandible with 4 teeth…………………..……………...…...… A. albipes (Folsom, 1927)
– Mandible with more than 4 teeth…………………...……………..……..……..2
2 Black with yellow strips, mandible with 6 teeth……………………………………… ………………………………………………….. A. arleana Mendonça and Fernandes, 1999
– Light gray, mandible with 5 teeth…….... A. intermedia Fernandes and Mendonça, 2004
3.1.2. Brachystomellidae Stach, 1949
Brachystomella agrosa Wray, 1953
Brazilian Oceanic Island Records. FN (3°50′54.21″ S; 32°25′45.56″ O), TF, 20 Jul 2012, E.C.A. Lima and D. Zeppelini leg., CRFS# 14400–14401, 14409–14414, 14422. FN (3°50′58.48″ S; 32°26′3.83″ O), SF, 25 July 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14415–14418. FN (3°51′14.32″ S; 32°26′12.62″ O), TF, 25 July 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14450–14455, 14457–14461, 15094–15096, 15098. FN (3°50′18.66″ S; 32°24′3.09″ O), SF, 07 August 2012, E.C.A. Lima and D.D. Silva CRFS# 14456. FN (3°51′3.73″ S; 32°26′0.38″ O), SF, 27 July 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14399, 14419–14421, 14423, 14426–14427. FN (3°50′36.76” S; 32°25′12.64” O), SF, 26 Jul 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 9565–9568, 9593, 14449. FN (3°50′48.92” S; 32°25′13.61″ O), TF, 26 July 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 9562–9564, 9571–9592. FN (3°48’45.06″ S; 32°23′14.07″ O), TF, 19 Jul 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 15063, 15069. FN (3°52′8.88” S; 32°26′13.57″ O), SF, 01 August 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14402–14403, 14405, 14407, 14425, 14429, 14437, 14440. FN (3°52′8.88″ S; 32°26′13.57″ O), SF, 14 Aug 2012, D.D. Silva leg, CRFS# 14408. FN (3°51′50.41″ S; 32°26′5.44″ O), TF, 01 August 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14406. FN (3°51′52.31″ S; 32°26′38.80″ O), TF, 31 July 2012, E.C.A. Lima and A.S. Ferreira leg., CRFS# 14424, 14428, 14435, 14436, 14438–14439, 14443–14444.
Brazilian Occurrence. In Brazilian oceanic island B. agrosa recorded in Fernando de Noronha Archipelago on Fernando de Noronha and Rata Island (SF, TF sites), the specimens were collected in superficial soil and leaf litter. In Brazil, this species is widely distributed and has been registered in the following states CE, BA, PI, PE, PB, RN, ES, SP and RJ [13] (Figure 13).
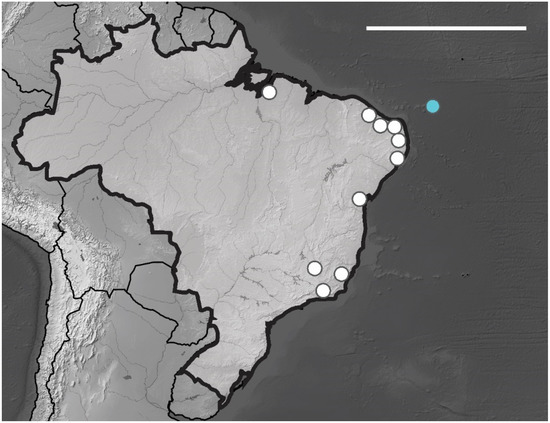
Figure 13.
Distribution map for Brachystomella agrosa Wray, 1953 recorded in Brazil. Blue circle, Fernando de Noronha; white circles, Brazilian states. Scale bar: 3000 km.
Biogeographic Distribution. The type locality of this species is Maricao, Puerto Rico, collected by M. Capriles, take on Road 27, at 120 m altitude. According to [1], this species has been found in the following biogeographic regions: Caribbean mainland (24a), Antillean and South Florida (24b), Venezuela and Guiana (25), Amazon (26), Northeast and Central Brazil (27). Neotropical species.
Remarks. In addition to the original description [17], several authors contributed to the description morphological of this taxon, such as: [18,19,20,21]. The specimens of B. agrosa examined agree with the above descriptions. For comparisons between Brachystomella species recorded in Brazil, see Table 5.
Table 5.
Morphological characters of Brachystomella Agren, 1903 species recorded from Brazil.; AP—apical bulb vesicles; PAO— PAO vesicle number; Mx T—maxilla teeth; Ten h—tenent hair; TT— tenaculum teeth; DC—dental chaetae number; TL—type locality; “?” —missing date
Identification Key for the Species of Brachystomella Ågren, 1903 Recorded in Brazil
1 2 + 2 eyes……….…………….………………………. B. garayae Queiroz and Weiner, 2011
– With more than 2 + 2 eyes………………………….………………………………...….. 2
2 7 + 7 eyes……………………………………………………... B. septemoculata Denis, 1931
– 8 + 8 eyes…………………………………………………………………………………… 3
3 With 3 dental chaetae……………………… B. villalobosi Cassagnau and Rapoport, 1962
– With more than 3 dental chaetae……………………………………………………….. 4
4 With 5 dental chaetae……………………………………………………………………. 5
– With 6 dental chaetae………………………... B. ceciliae Fernandes and Mendonça, 2004
5 Th I with 2 + 2……………………………………………………………………………… 6
– Th I with 3 + 3………………………………………………….. B. parvula (Schäffer, 1896)
6 tenent hair acuminate……………………………………...………………..………….. 7
– tenent hair capitated……………………………….. B. platensis Najt and Massoud, 1974
7 Apical bulb simple……………………………………………………………………….. 8
– Apical bulb with 3 vesicles……………………………………………………………….. 9
8 PAO with 5 vesicles…………………………..……...…………... B. contorta Denis, 1931
– PAO with 4 vesicles………………………….……………………. B. agrosa Ågren, 1903
9 PAO with 6 vesicles……………………………….. B. saladaensis Weiner and Najt, 2001
– PAO with 4 vesicles………………...…… B. nordestina Souza, Bellini and Weiner, 2018
3.1.3. Hypogastruridae Börner, 1906
Acherontiella sp.
Examined material. Female on slide, Brazil, Fernando de Noronha Archipelago, Fernando de Noronha Island, (3°50′36.76″ S; 32°25′12.64″ O), SF, 26 July 2012, E.C.A. Lima and A.S. Ferreira leg., deposited at the CRFS-UEPB # 14568.
Diagnosis. Color in ethanol white. Tegumental granulation rather strong. Body length: 746 µm; measurements as in Table 4.
Ant IV with 5 globular S-chaetae which 2 lateroexternal very large, placed in cavities, and the other 3 are smaller; dorsoexternal microsensillum and small subapical organite; present small non-retractile apical vesicle in ventro subapical position. Ant III and IV fused dorsally, the ventral separation well-marked. Sensory organ of Ant III consisting of two small internal sensilla, two sub-cylindrical (ring-shaped) guard sensilla and ventral microsensillum. Ant II–I with 11 and 7 chaetae, respectively. Eyes and PAO absent. Buccal cone typical of the genus. Pre-labral/labrum formula 4/554 chaetae, anterior labral chaetae spiniform. Dorsal chaetotaxy with rather short, ordinary chaetae, with long thin S-chaetae, their formula per half tergum: 022/11111. Abd VI without anal spines. Ventral tube with 4 + 4 chaetae. Tita I, II and III with 18, 18 and 17 chaetae, respectively. Femora I, II and III with 11, 11,10 chaetae, respectively, trochanters with 3,4,4 chaetae each, coxae I, II and III with 3, 8 and 7 chaetae.
Remarks. Acherontiella sp. [3] is similar to Acherontiella candida (Delamare Deboutteville, 1952), A. colotlipana Palacios-Vargas and Thibaud, 1985 and A. kowalskiorum Weiner and Najt, 1998 by type of guard sensilla of the Ant III organ (ring-shaped) and number of sensilla in Ant IV (with 5 globular sensilla). Acherontiella sp. differs from these species by anterior labrum chaetae (spiniform versus thin and smooth). For comparisons between Acherontiela species recorded in Brazil, see Table 6.
Table 6.
Morphological characters of Acherontiella Absolon, 1913 species recorded from Brazil. S VI—spine on Abd VI; AL—anterior labral chaetae; Ten h—tenent hair; BD—body length in µm; NS—sensilla number of Ant IV; Tr—trochanter; TL—type locality; “?” —missing date
Distribution. Acherontiella sp. was found only on Fernando de Noronha Island in the SF region. The specimen was collected in soil litter. Additional material is required in order to properly describe this potentially new species. For other congeners records in Brazil, see Figure 14.
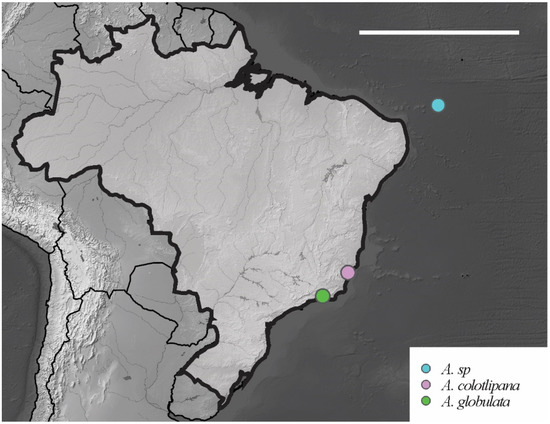
Figure 14.
Distribution map for Acherontiella Absolon, 1913 species recorded in Brazil. Scale bar: 3000 km.
Identification Key for the Species of Acherontiella Recorded in Brazil
1 Ant IV with 6 globular sensilla…………….. A. globulata Thibaud e Massoud, 1980
- Ant IV with 5 globular sensilla……………….………...……………………………….. 2
2 Labrum with 4 smooth anterior chaetae…………………………………………………. ………………………………………… A. colotiplana Palacios-Vargas and Thibaud, 1985
- Labrum with 4 spiniform anterior chaetae…….……. A. sp. Lima and Zeppelini, 2015
Paraxenylla zeliae sp. nov Lima and Zeppelini
Type material. Holotype male on slide, Brazil, Atol das Rocas, Cemitério Island, (3°51′47.15″ S; 33°48′54.14″ O), Oct 2015, E.C.A. Lima leg., deposited at the CRFS-UEPB # 14567. Paratypes: 2 females on slides, Brazil, Atol das Rocas, Farol Island, (3°51′28.14″ S; 33°48′44.87″ O), guano, Sept 2015, E.C.A. Lima leg., deposited at the CRFS-UEPB # 14565, 14566.
Description. Habitus typical of the genus. Color in ethanol blue. Body length average: 690 µm (n = 10); holotype measurements as in Table 2. Body with short acuminate chaetae (7–9 μm). Macrochaetae and S-chaetae (14–18 μm) about twice the length of ordinary chaetae.
Antenna (Figure 15A). Ant IV with 4 thick S-chaetae and a small S-microchaeta between the 3 S-chaetae of the external group. Subapical organ present (Figure 15A). Apical bulb simple. Sensory organ of Ant III with 5 S-chaetae (2 small, S3 and S4, behind a fold of the integument, 2 longer guard chaetae S2 and S5, and one external microchaeta S1). Ant II–I with 11 and 7 chaetae, respectively.
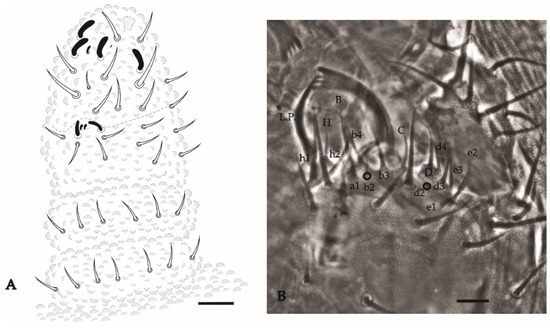
Figure 15.
Paraxenylla zeliae sp. nov. (A), dorsal view of antenna; (B), labial palp and mandible apex, nomenclature after [22]. Scale bars: 10 µm.
Head. Eyes 5 + 5 well pigmented. Labral formula: 27/554 chaetae. Mandible with 4 apical teeth (Figure 15B). Internal process of labium long, labial palps as in Figure 15B. Maxillae with 6 hypertrophied fringed lamellae.
Thorax I with 3 + 3 to 4 + 4 dorsal chaetae. Dorsal chaetotaxy of head and Th in Figure 16a; m1 absent, m2 present on Th II–III as macrochaetae, ms on Th II not visualized.
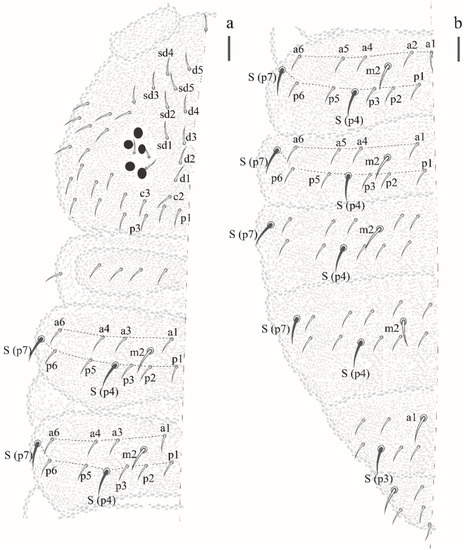
Figure 16.
Paraxenylla zeliae sp. nov. (a) dorsal view of head and Th I–III, scale bar: 10 µm. (b) dorsal view of Abd I–VI, scale bar: 10 µm.
Dorsal chaetotaxy of abdomen as in Figure 16b. Axial S chaetae 44/44,443; m2 on Abd I–III and a1 on Abd V as macrochaetae. Abd VI dorsally with 5 + 5 chaetae, one posterior chaetae and 3 An microchaetae on the supra-anal valve.
Leg. Subcoxa 1 I–III with 1,3,3 chaetae. Subcoxa 2 I–III with 0,1,1 chaetae. Coxae I–III with 3,6, 7 chaetae. Trochanters with 5 chaetae each, femora I, II, III with 12, 12, 11 chaetae, respectively. On each femur, 1 ventral chaeta longer than others. Tita I, II, III with 19, 19, 18 chaetae, respectively, each with 4 capitate tenent hairs, 2 internal longer than 2 external. Unguis without teeth, no empodial appendix (Figure 17a).
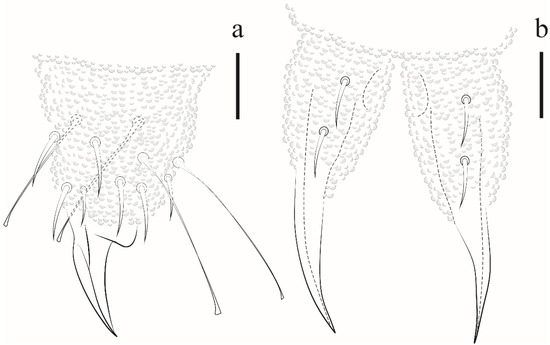
Figure 17.
Paraxenylla zeliae sp. nov. (a) Tita III, scale bar: 20 µm. (b) mucrodens, scale bar: 20 µm.
Ventral tube with 1 + 1 chaetae. Abd II with 1 + 1 ventral chaetae, Abd III with 5 + 5 chaetae. Tenaculum with 3 teeth on each ramus. Mucrodens with two chaetae (Figure 17b). Female with 3 + 3 pregenital chaetae, 4–6 circumgenital and 2 eugenital. Male with 3 + 3 pregenital chaetae, 8–10 circumgenital and 4 + 4 eugenital. Each anal valve with 11–13 chaetae. Anus terminal, no anal spines.
Etymology. The species is named after Head of the Biological Reserve Atol das Rocas Mrs. Maurizélia de Brito Silva, in honor of a lifetime dedication to conservation and assist scientific research on the Atol das Rocas.
Remarks. Paraxenylla zeliae sp. nov. is similar to P. affiniformis (Stach, 1930) (cosmopolitan), P. piloua Thibaud and Weiner, 1997 (New Caledonia), P. lapazana Palacios-Vargas and Vázquez, 1988 (Mexico), P. cubana Palacios-Vargas and Janssens, 2006 (Cuba), P. peruensis Palacios-Vargas and Janssens, 2006 (Peru) P. mahahualana Palacios-Vargas and Vázquez, 2018 (Mexico) by ventral tube with 1 + 1 chaetae, Th II–III m1 absent and m2 present, Abd I–III p2 and Abd V a1 as macrochaetae, Abd V S-chaetae in p4 position. The new species differ from P. cubana by tenaculum teeth (3 + 3 versus 2 + 2) and capitate tenent hair on Tita III (4 versus acuminate). Paraxenylla zeliae sp. nov. has 4,4,4 clavate tenent hair while P. lapazana 3,3,3 clavate or acuminate, P. affiniformis 5,5,5 acuminate and P. mahahualana 5,5,5 clavate. The new species share the same number of tenent hair with P. piloua and P. peruensis. However, P. zeliae sp. nov. differ from P. piloua by tenent hair morphology (clavate versus acuminate), body size (490–720 versus 360–400) and femora III (11 versus 10 chaetae); differ from P. peruensis by Th I dorsal chaetotaxy (4 + 4 versus 5 + 5), Abd II ventral (1 + 1 versus 3 + 3) and axial chaetae formula (44/44443 versus 44/44462). For other morphological comparisons between Paraxenylla species recorded in Brazil, see Table 7.
Table 7.
Morphological characters of Paraxenylla Murphy, 1965 species recorded from Brazil. VT—ventral tube chaetae; TT—tenaculum teeth; VC—ventral chaetae Abd II; Ten h—tenent hair; BD—body length in µm; NS—sensilla number of At. IV; TL—type locality. Ac—acuminate; Cl—clavate.
Distribution. The new species was found on Atol das Rocas Island in the SF sites. The specimen was collected in sandy soil near the psammophile vegetation and seabird guano on rocks. For the congeners registered in Brazil, see Figure 18.
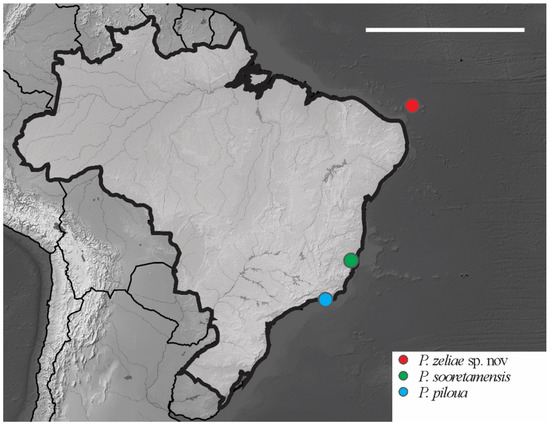
Figure 18.
Distribution map for Paraxenylla Murphy, 1965 species recorded in Brazil. Scale bar: 3000 km.
Identification Key for Paraxenylla Murphy, 1965 Species Recorded in Brazil
1 Ventral tube 1 + 1 chaetae.................................................................................................... 2
– Ventral tube 4 + 4 chaetae.......................... P. sooretamensis Queiroz and Deharveng, 2008
2 tenent hair clavate, femora III with 11 chaetae................................. P. zeliae sp. nov.
– tenent hair acuminate, femora III with 10 chaetae. P. piloua Thibaud and Weiner, 1997
Willemia insularum sp. nov. Lima and Zeppelini
Type material. Holotype female on the slide, Brazil, Rio Grande do Norte, Atol das Rocas, (03°51′50″ S, 33°48′48″ W), intertidal sand, 08–18 September 2015, E.C.A. Lima leg., deposited at the CRFS-UEPB # 12261. Paratypes: 2 males and 3 females on slides, same data as holotype, deposited at the CRFS-UEPB # 12262, 12263, 12264, 12265, 12266. Additional material: 2 females and 1 male on slides, Brazil, Pernambuco, Fernando de Noronha (3°50′58.48″ S; 32°26′3.83″ O and 3°50′36.76″ S; 32°25′12.64″ O), 25–26 July 2012, E.C.A. Lima and A.S. Ferreira leg., deposited at the CRFS-UEPB # 14562, 14563, 14564.
Description. Habitus typical of the genus. Color in ethanol white. Tegumental granulation fine and regular. Body length: 490 µm (n = 5); holotype measurements as in Table 2.
Body with short acuminate ordinary chaetae, some slightly longer. Sensory chaetae are lanceolate and longer than ordinary chaetae. Antennae are somewhat shorter than cephalic diagonal.
Antenna (Figure 19). Ant IV with 4 (S1, S2, S8, S9) subcylindrical sensilla, 2 globular sensilla S7 and S4, hidden each in a separate cavity. Microsensillum (ms) set in the cavity of S7 sensillum. Sensilla S2 and S9 hardly differentiated from ordinary chaetae, S1 and S8 subcylindrical. Apical bulb small and oval, subapical organelle set in a deep cavity, roundish. Sensorial organ of Ant III with 2 long guard chaetae, 2 long subcylindrical sensilla, 1 ventral microsensillum, and a large integumentary fold hiding 2 internal sensilla. Ant II and I with 11 and 6 chaetae, respectively.
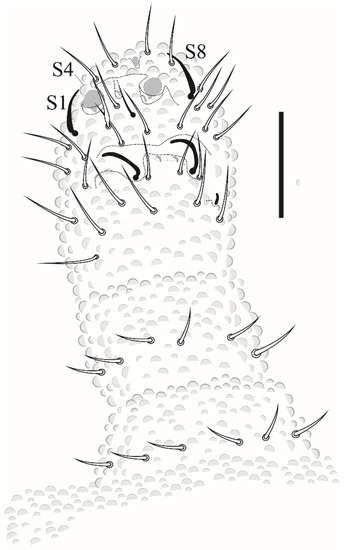
Figure 19.
Willemia insularum sp. nov. 33, dorsal view of antenna. Scale bar: 10 µm.
Eyes and a0 chaetae absent, PAO with 7–8 vesicles (Figure 20a). Head dorsal (Figure 20b). Prelabrum/labrum with 2/534 chaetae (Figure 21a). Along cephalic groove 3 + 3 chaetae (Figure 21b).
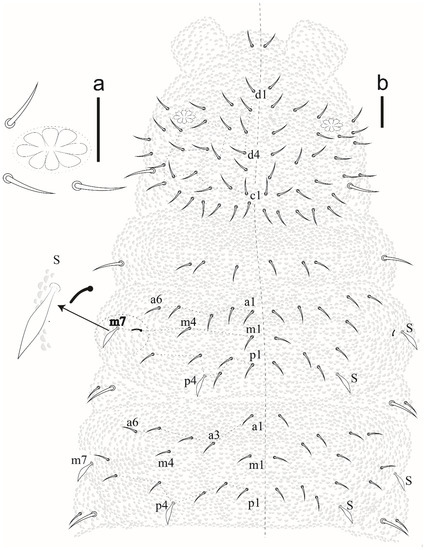
Figure 20.
Willemia insularum sp. nov. (a) postantennal organ, scale bar: 20 µm. (b) dorsal view of head and thorax I–III. Scale bar: 10 µm.
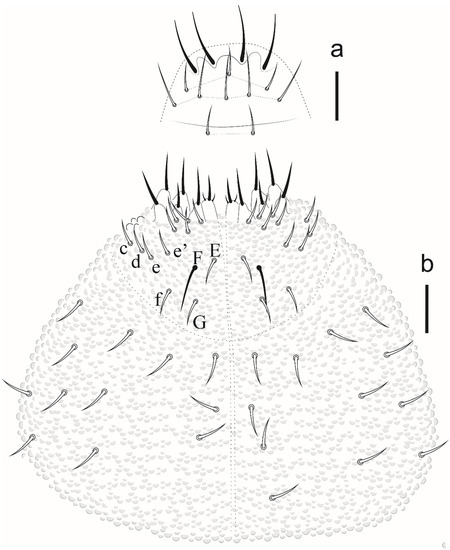
Figure 21.
Willemia insularum sp. nov. (a) prelabrum/labrum, scale bar: 10 µm. (b) ventral view of head, scale bar: 10 µm.
Dorsal chaetotaxy (Figure 20b and Figure 22). S-chaetae per half tergum formula: 22/11111, in m7 and p4 position on Th II and III, in p4 position on abdominal I to IV and p2 position on Abd V. Sensilla p4 on Th II–Abd I easily differentiated from ordinary chaetae. Abd IV without m-row. With two anal spines on m1 position.

Figure 22.
Willemia insularum sp. nov., dorsal view of Abd I–VI, anal spine details. Scale bars: 10 µm and anal spine detail, 20 µm.
Ventral abdominal chaetotaxy is presented in Figure 23a. Sternum of Abd IV with 12 + 12 chaetae. Sternum of Abd II with a-row with 2 chaetae on each side (a3 present). Anal lobes with 16 + 16 chaetae (2 i, 3 hr, e and b absent). Ventral tube with 4 + 4 chaetae (Figure 23b). Tita I, II and III with 12, 12, 11 chaetae. Unguis without teeth, empodial appendage vestigial (Figure 23c).
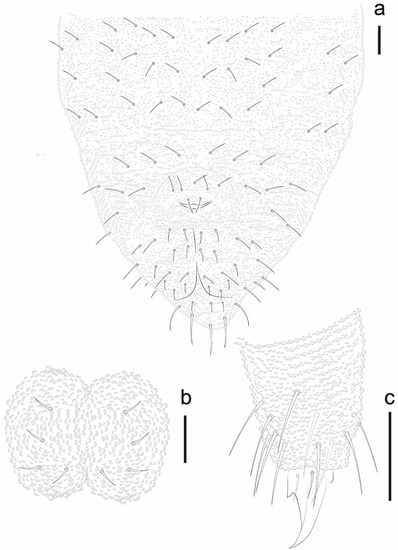
Figure 23.
Willemia insularum sp. nov. (a) ventral view of Abd II–VI, scale bar: 10 µm. (b) ventral tubes, scale bar: 20 µm. (c) Tita III, scale bar: 20 µm.
Etymology. An allusion for being the only new species of this study that occurs in two different island regions.
Remarks. Willemia insularum sp. nov. shares characteristics with the buddenbrocki-group species sensu D’Hese 2000 [23,24,25] by S-chaetae S4 and S7 of Ant IV both large, globular, placed in the cavity, and covered in part by tegumental fold and absence of a0 chaetae on the head. The new species presents apical vesicle, Abd V with S-chaetae in p2 position; 3 + 3 chaetae on Th I, presence of anal spines; Abd IV and V with S-chaetae lanceolate. The new species is similar to W. dhesei Bu, Potapov and Gao, 2012 (Shandong, Pacific coast of China), W. delamarei Prabhoo, 1971 (Indian region); W. antennomonstrum Bu, Potapov and Gao, 2012 (Hainan, South China). Willemia insularum sp. nov. differs from W. dhesei by chaetotaxy of Abd IV (without m-row versus m1 to m4 present) and postantennal organ (7–8 versus 5–6 vesicles). W. delamarei has a postantennal organ with 8–12 vesicles, S-chaetae absent in Th II and Abd I while the new species present postantennal organ with 7–8 vesicles and 2 S-chaetae in Th II (m7 and p4) and 1 in Abd I (p4). Willemia insularum sp. nov., can be easily distinguished from W. antennomonstrum by the sensory organ of Ant III usual for the genus versus 2 expanded and granulated guard sensilla. For other morphological comparisons between Willemia species recorded in Brazil, see Table 8.
Table 8.
Data matrix of 52 characters coded for Willemia species recorded in Brazil. Character numbers correspond to the characters listed in [26]. Polymorphic characters coded as follow [23], 0 or 1 = a; 0 or 2 = b; 0, 2 or 3 = c. “?”—missing date.
Distribution. The new species were found on Fernando de Noronha and Atol das Rocas Islands in the SF sites. The specimens were collected in sand near the psammophile vegetation, litter and seabird guano. For the congeners registered in Brazil, see Figure 24.
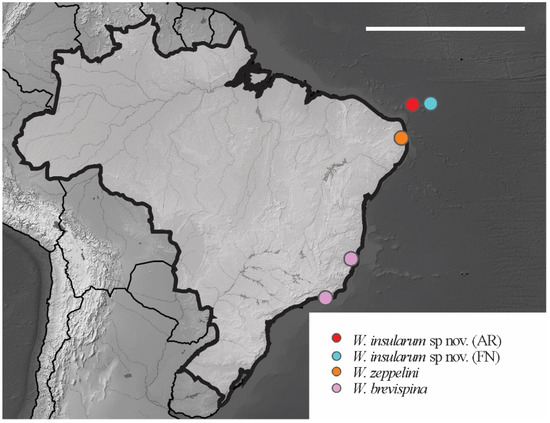
Figure 24.
Distribution map for Willemia Börner, 1901 species recorded in Brazil. Scale bar: 3000 km.
Identification Key for Willemia Species Recorded in Brazil
1 Anal spine absent……………………………... W. zeppelinii D’Hese and Thibaud, 2011
– Anal spine present……………………………………………………………………….. 2
2 Abd IV and V with S-chaetae subcylindrical and acuminate……...…………………. ………………………………………………………………….W. brevispina Hüther, 1962
– Abd IV and V with S-chaetae lanceolate…………………….... W. insularum sp. nov.
Xenylla yucatana Mills, 1938
Brazilian Oceanic Island Records. FN (3°50′43.00″ S; 32°25′34.04″ W), SF, 20 Jul 2012, E.C.A Lima and D. Zeppelini leg., CRFS# 14518, 14519, 14522, 14528. FN (3°50′54.21″ S; 32°25′45.56″ W), TF, 20 July 2012, E.C.A Lima and D. Zeppelini leg., CRFS# 14520, 14525, 14527, 14529, 14530, 14531, 14535, 14536, 14553, 14560, 14561. FN (3°50′31.85″ S; 32°24′14.74″ W), TF, 07 August 2012, E.C.A Lima and D.D. Silva leg, CRFS#14474, 14491, 14545, 14546, 14547, 14548, 14549, 14550. FN (3°50′18.66″ S; 32°24′3.09″ W), SF, 07 August 2012, E.C.A Lima and D.D. Silva leg, CRFS# 14479, 14487, 14489, 14492, 14503–14505, 14511–14517, 14521, 14523–14524, 14526, 14532–14537, 14538–14544, 14551–14552, 14555–14556, 14558–14559, 14734, 14735–14768. FN (3°51′3.73″ S; 32°26′0.38″ W), SF, 27 July 2012, E.C.A Lima and A.S. Ferreira leg., CRFS# 14478, 14481, 14484. FN (3°50′36.76″ S; 32°25′12.64″ W), SF, 26 July 2012, E.C.A Lima and A.S. Ferreira leg., CRFS# 9373, 9374–9384, 9423–9533. FN (3°50′48.92″ S; 32°25′13.61″ W), TF, 26 Jul 2012, E.C.A Lima and A.S. Ferreira leg., CRFS# 9532. FN (3°51′50.41″ S; 32°26′5.44″ W), TF, 01 August 2012, E.C.A Lima and A.S. Ferreira leg., CRFS# 14462–14472, 14476–14477, 14480, 14485, 14486, 14493–14502.
Brazilian Occurrence. In Brazil, this species had already been registered in the states of ES, RJ and the island of Fernando Noronha [13] (Figure 25). Cosmopolitan species.
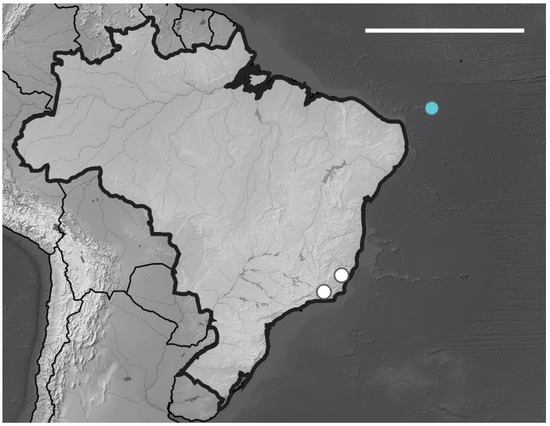
Figure 25.
Distribution map for Xenylla yucatana Mills, 1938 records in Brazil. Scale bar: 3000 km. Blue circle, Fernando de Noronha; white circles, Brazilian states. Scale bar: 3000 km.
Biogeographic Distribution. The type locality of this species in Mexico, Yucatan Peninsula, San Bulha Cenote Motul; specimens collected in bat feces [27]. Xenylla yucatana has circumtropical distribution distributed in the following biogeographic regions African Indian Desert (9), East African Steppe (13), South African (14), Madagascar (15), Ascension and St Helena (16), Continental South and East Asia (18), Malaysian (19), Hawaiian (20), New Caledonia (21), Melanesia and Micronesia (22), Caribbean Mainland (24a), Antillean and South Florida (24b), Venezuela and Guyana (25), Northeast and Central Brazilian (27), Andean (28), North and East Australia (32) according to [1].
Remarks. The specimens analyzed agree with descriptions by [28,29,30,31]. For other morphological comparisons between Xenylla species recorded in Brazil, see Table 9. In the oceanic islands, the species was found in great abundance on the island of Fernando de Noronha in the SF and TF sites.
Table 9.
Morphological characters of Xenylla Tullberg, 1869 species recorded from Brazil.; Ten h—tenent hair; OCM—ordinary chaetae morphology; TL—type locality; MD—mucrodens, MSD—mucro separate from dens; “?”—missing date.
Identification Key for Xenylla Tullberg, 1869 Species Recorded in Brazil
1 With 5 + 5 eyes……………………………………………………………………………… 2
– With 4 + 4 eyes……………………………………………………… X. yucatana Mills, 1938
2 Ant IV with 4–5 sensilla………………………………………………………………… 3
– Ant IV with 6 sensilla…………………………………………………………………… 8
3 With 1,1,1 tenent hair…………………………………… X. brasiliensis da Gama, 1978
– With 1,2,2 or 2,2,2 tenent hair…………………………………………………………… 4
4 With 1,2,2 tenent hair…………………………………………… X. welchi Folsom, 1916
– With 2,2,2 tenent hair……………………………………………………………………… 5
5 Manubrium with 28 chaetae……………………… X. hodori Neves and Mendonça, 2017
– Manubrium with more than 28 chaetae………………………………………………… 6
6 Mucro separate from dens……………………………………………………………… 7
– Mucro and dens fused …………………………………………………………………… 9
7 Creased cuticular structure and p6 chaetae present on Abd IV………………………… ……………………………………………………X. manuelae Queiroz and Mendonça, 2016
– Creased cuticular structure and p6 chaetae absent on Abd IV………………………………………………………………………………X. capixaba Fernandes and Mendonça, 2010
8 With 1,1,1 tenent hair………………………………… X. nirae Gama and Oliveira, 1994
– With 1,2,2 tenent hair………………………… X. wandae Queiroz and Mendonça, 2016
9 Dens with 2 posterior chaetae……………………………… X. maritima Tullberg, 1869
– Dens with 1 posterior chaetae…………………………… X. subcavernarum Gama, 1969
3.1.4. Isotogastruridae Thibaud and Najt, 1992
Isotogastrura mucrospatulata Palacios-Vargas, Lima and Zeppelini, 2013
Brazilian Oceanic Island Records. FN (3°50′40.61″ S, 32°25’35.41″ O), SB, 20 July 2012, E.C.A. Lima and A.S. Ferreira Leg., CRFS# 3680–3696. FN (3°50′13.36″ S, 32°24’0.28″ O), SB, 07 August 2012, E.C.A Lima and D. Zeppelini leg., CRFS# 3697–3701. FN (3°50′30.12″ S, 32°25’3.84” O), SB, 20 July 2012, E.C.A. Lima and A.S. Ferreira Leg., CRFS# 3702. FN (3°50′30.12″ S, 32°25′3.84″ O), SB, 20 Jul 2012, E.C.A. Lima and A.S. Ferreira Leg., CRFS# 3703–3708. FN (3°51′19.98″ S, 32°26′43.23″ O), SB, 20 July 2012, E.C.A. Lima and A.S. Ferreira Leg., CRFS# 3709–3710.
Distribution: This species is known only in the sand beaches of the Fernando de Noronha Islands. Small body size (350 µm) of I. mucrospatulata indicates that it inhabits narrow passages among the grains of sand [32]. Interstitial Collembola looks like typical euedaphic species but is flexible and slender enough to be able to move between sand grains of small size without changing the pore architecture [33]. All specimens in this study were found in the sand at the intertidal zone (SB sites).
Remark. The specimens analyzed agree with the original description by [34]. For others, morphological comparisons between Isotogastrura species recorded in Brazil, see Table 10.
Table 10.
Morphological characters of Isotogastrura species record from Brazil modified from [34]; 1, Abd V tegumentary tubercle; 2. Abd I sternite chaetae number; 3. Abd III tergite with a pair of tubercles; 4. Prelabral chaetae (presence vs. absence); 5. Sensillum of Ant III simple or bifid; 6. Dental chaetae; 7. Shape of mucro; 8. Genital chaetae; 9. Number of sensilla on Ant IV; 10. Tubercles on Abd VI; 11. Chaetae on labial triangle; 12. Chaetae on manubrium; 13. Chaetae on furcal subcoxa; 14. Basal manubrium Chaetae. Abbreviations: BI = bifid; H = hooked; MOD = modified; SP = spatulate; un = unmodified; “?”—missing date.
3.2. Species List of Brazilian Poduromorpha
In Brazil, Poduromorpha fauna is registered in 18 states and 3 oceanic islands (Table 11). The states of Acre, Alagoas, Goiás, Maranhão, Roraima, Rio Grande do Sul, Santa Catarina, Tocantins and the oceanic Archipelago of Trindade and Martim Vaz have no record of Poduromorpha. Including the new taxa presented in this study, 139 Brazilian species of Poduromopha distributed in the families Brachystomellidae Stach, 1949 (24 spp.); Hypogastruridae Börner, 1906 (31 spp.); Isotogastruridae Thibaud and Najt, 1992 (2 spp.); Neanuridae Börner, 1901 (71 spp.); Odontellidae Massoud, 1967 (1 sp.); Onychiuridae Lubbock, 1867 (4 spp.); Tullbergiidae Bagnall, 1935 (6 spp.), see Table 2. This number represents an increase of 41 new species records since the last synthesis by [2].
Table 11.
Species of Poduromorpha recorded from Brazilian states and oceanic islands–updated from [2]. *—New record after [2]. In parentheses, information obtained from non-original reference. “?” following state abbreviation—questionable state record. “un”—unspecified or unknown Brazilian collection habitats. World distribution—see Methods/Species list of Brazilian Poduromorpha. “?” following the distribution abbreviation—questionable distribution record. Habitat—representative type of habitat for the species. “un”—unspecified or unknown habitat (Brazilian distribution, locality abbreviations refer to Brazilian states and Insular locality as listed in Table 1). Family names in bold.
4. Discussion
The first Collembola species registered in Brazil was the Seira musarum Ridley, 1890 (Entomobryomorpha), whose type locality is Fernando de Noronha. However, after this pioneering study, the only works with Collembola in Brazilian oceanic islands were the description of Isotogastrura mucrospatulata Palacios, Lima and Zeppelini 2012 [36] and the species survey done by Lima and Zeppelini, 2015 [3] both in Fernando de Noronha.
Our results show that Poduromorpha fauna of the oceanic islands showed a high percentage of species known only from their type localities and with a great potential for endemisms. All of the Poduromorpha species found in the São Pedro and São Paulo archipelago and Atoll das Rocas are undescribed and with consistent morphological differences in relation to the species found on the continent. Fernando de Noronha, the only Brazilian oceanic island where tourist visitation is allowed, one endemic (I. mucruspatulata), four with wide continental distribution and two new species described here. These results may be useful in the management and environmental monitoring programs of oceanic islands in Brazil.
Although the increase in the number of Brazilian Poduromorpha species over the past decade has been relevant (about 35% since [2]) when we consider the variety of Brazilian continental biomes and the complex physiognomy of these areas, current knowledge of the Poduromorpha fauna can still be considered scarce and far from providing a reliable overview of its biogeographical panorama.
Author Contributions
Conceptualization, all authors; methodology, E.C.A.d.L.; formal analysis, all authors; investigation, E.C.A.d.L. and D.Z.; resources, all authors; data curation, D.Z.; writing—original draft preparation, E.C.A.d.L. and T.C.d.S.; writing—review and editing, E.C.A.d.L. and G.C.Q.; supervision, M.C.d.M. and D.Z.; project administration, E.C.A.d.L.; funding acquisition, all authors. All authors have read and agreed to the published version of the manuscript.
Funding
E.C.A.d. Lima is granted by CNPq (Protax, Process 440521/2015-7), M.C.d. Mendonça is granted by CNPq (Process 307644/2015-4), D Zeppelini is granted by CNPq (Process 301803/2012-9).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Institutional Review Board of Chico Mendes Institute for Biodiversity Conservation (ICMBio) (SISBIO 32469).
Informed Consent Statement
Not applicable.
Data Availability Statement
All the specimens collected and used in this research are deposited in the Coleção de Referência de Fauna de Solo at the State University of Paraiba (CRFS-UEPB) under tumble number as cited in the materials and methods.
Acknowledgments
The ICMBio provided the permits for collection; Marinha do Brasil provided the transportation to St Peter and St Paul rocks, the preparatory training, and support during the scientific expeditions; Aíla Soares Ferreira and Diego Dias da Silva gave support during the fieldwork in Fernando de Noronha; Luís Carlos Stievano helped in the ordenation of the samples and assistanced in all laboratory work; anonymous reviwers greatly improved the manuscript. A special thanks for the crew members of the boats Borandá and Alfa, for passages to the most incredible places I have yet had the opportunity to know: the Oceanic Islands.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 2 December 2019).
- Abrantes, E.A.; Bellini, B.C.; Bernardo, A.N.; Fernandes, L.H.; Mendonça, M.C.; Oliveira, E.P.; Queiroz, G.C.; Sautter, K.D.; Silveira, T.C.; Zeppelini, D. Errata Corrigenda and update for the “Synthesis of Brazilian Collembola: An update to the species list”. Zootaxa 2012, 3168, 1–21. [Google Scholar] [CrossRef]
- Lima, E.C.A.; Zeppelini, D. First survey of Collembola (Hexapoda: Entognatha) fauna in soil of Archipelago Fernando de Noronha, Brazil. Flor. Entomol. 2015, 98, 368–369. [Google Scholar] [CrossRef]
- Jordana, R.; Arbea, J.I.; Simón, C.; Luciáñez, M.J. Fauna Iberica, Collembola Poduromorpha; Museo Nacional de Ciencias Naturales: Madrid, Spain, 1997; Volume 8, pp. 1–807.
- D’Haese, C.A. Homology and morphology in Poduromorpha (Hexapoda, Collembola). Eur. J. Entomol. 2003, 100, 385–407. [Google Scholar] [CrossRef]
- Potapov, M.B.; Banasko, J.A.A. New species of springtail from Cuba with comments on the role of chaetotaxy in diagnostics of the Friesea (Collembola, Neanuridae) species. Zoolog. Zhur. 1985, 64, 1162–1167. [Google Scholar]
- Deharveng, L.; Bedos, A. Taxonomy and cladistics analysis of the Thai species of Friesea Dalla Torre 1895 (Collembola Ne-anuridae). Trop. Zool. 1991, 4, 287–315. [Google Scholar] [CrossRef]
- Weiner, W.M.; Bedos, A.; Deharveng, L. Species of the genus Friesea(Collembola, Neanuridae) from New Caledonia and Vanuatu. Zoosystema 2009, 31, 507–518. [Google Scholar] [CrossRef]
- Culik, M.P.; Filho, D.Z. Diversity and distribution of Collembola (Arthropoda: Hexapoda) of Brazil. Biodivers. Conserv. 2003, 12, 1119–1143. [Google Scholar] [CrossRef]
- Good, R. The Geography of Flowering Plants; Longman Group: London, UK, 1974; pp. 1–574. [Google Scholar]
- Christiansen, K.; Bellinger, P. The biogeography of Collembola. Bull. Entomol. Polog. 1995, 64, 279–294. [Google Scholar]
- Cassagnau, P. Les espèces européennes du genre Friesea [Collemboles Poduromorphes]. Bull. Soc. D’Hist. Nat. Toul. 1958, 93, 17–29. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Bellini, B.C. Collembola in Catálogo Taxonômico da Fauna do Brasil. Available online: http://fauna.jbrj.gov.br/fauna/faunadobrasil/379 (accessed on 8 August 2019).
- Folsom, J.W. Insects of the subclass Apterygota from Central America and the West Indies. Proc. U. S. Natl. Mus. 1927, 72, 1–16. [Google Scholar] [CrossRef][Green Version]
- Massoud, Z. Les Collemboles Poduromorphes du Surinam. Stud. Suriname Guyanas 1963, 20, 42–51. [Google Scholar]
- Mendonça, M.C.; Fernandes, L.H. Contribuição para o conhecimento do gênero Arlesia Handschin (Collembola: Neanuridae, Pseudachorutinae). Rev. Bras. Zool. 1999, 16, 1195–1201. [Google Scholar] [CrossRef]
- Wray, D.L. New Collembola from Puerto Rico. J. Agric. Univ. Puerto Rico 1953, 37, 140–150. [Google Scholar] [CrossRef]
- Massoud, Z.; Thibaud, J.M. Les Collemboles des Petites II—Neanuridae. Rev. D´Écól. Biol. Sol. 1980, 17, 591–605. [Google Scholar]
- Najt, J.; Thibaud, J.-M.; Weiner, W. Collemboles (Insecta) Poduromorphes de Guyanae Française. Mem. Mus. Nat. D´Hist. Natu. 1990, 4, 95–121. [Google Scholar]
- Mendonça, C.; Arlé, R. Variação quetotáxica em Brachystomella agrosa Wray, 1953 (Collembola, Neanuridae). Bol. Mus. Nac. 1992, 351, 1–9. [Google Scholar]
- Weiner, W.M.; Najt, J. Species of Brachystomella (Collembola: Brachystomellidae) from the Neotropical region. Eur. J. Entomol. 2001, 98, 387–413. [Google Scholar] [CrossRef]
- Fjellberg, A. The Labial Palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- D’Haese, C.; Thibaud, J.-M. Description and phylogenetic position of a new Willemia species (Collembola: Hypogas-truridae) from the littoral coast of Brazil. Zootaxa 2011, 2932, 33–40. [Google Scholar] [CrossRef]
- Bu, Y.; Potapov, M.B.; Gao, Y. Littoral Willemia (Collembola: Hypogastrurudae) of china with a description of two new species and a new case of convergente between Hypogastruridar and Onychiuridae. Flor. Entomol. 2015, 95, 580–586. [Google Scholar] [CrossRef]
- Zon, S.D.; Bedos, A.; D´Haese, C. Phylogeny of the genus Willemia (Collembola: Hypogastruridae) and biogeography of the W. buddenbrocki-group with description of a new species from Ivory Coast (western Africa). Zootaxa 2015, 3980, 230–240. [Google Scholar] [CrossRef]
- D´Haese, C. Is Psammophily and Evolutionary dead end? A phylogenetic test in the genus Willemia (Collembola: Hypogas-truriade). Cladistics 2000, 16, 255–273. [Google Scholar] [CrossRef]
- Mills, H.B. Collembola from Yucatan caves. Carn. Inst. Washing. Publ. 1938, 491, 183–190. [Google Scholar]
- Murphy, D.H. Collembola Poduromorpha from the Gambia (West Africa). J. Zool. 2010, 146, 388–411. [Google Scholar] [CrossRef]
- Gama, M.M. Notes taxonomiques et lignées généalogiques de quarante deux espèces et sous-espèces du genre Xenylla (Insecta: Collembola). Mem. Est. Mus. Zool. Univers. Coimbra. 1969, 308, 1–61. [Google Scholar]
- André, H.M. Redescription of Xenylla yucatana Mills, 1938 (Collembola, Hypogastruridae). Fol. Entomol. Mex. 1988, 74, 69–77. [Google Scholar]
- Christiansen, K.; Bellinger, P. Insects of Hawaii. A Manual of the Insects of the Hawaiian Islands, Including an Enumeration of the Species and Notes on Their Origin, Distribution, Hosts, Parasites; University of Hawaii Press: Honolulu, HI, USA, 1992; p. 445. [Google Scholar]
- Potapov, M.B.; Bu, Y.; Gao, Y. First record of the littoral family Isotogastruridade (Collembola) in Asia. Zookeys 2011, 136, 23–29. [Google Scholar]
- Thibaud, J.-M. Recent advances and synthesis in biodiversity and biogeography of arenicolous Collembola. Ann. Soc. Entomol. Fr. 2007, 43, 181–185. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.G.; Lima, E.C.A.; Zeppelini, D. A new species of Isotogastrura (Collembola: Isotogastruridae) from North-eastern Brazil. Flor. Entomol. 2013, 96, 1579–1587. [Google Scholar] [CrossRef]
- Cassagnau, P.; Rapoport, E.H. Collemboles d’Amérique du Sud, I Poduromorphes. Biol. Amér. Aust. 1962, 1, 139–184. [Google Scholar]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mitt. Natu. Mus. Hamb. 1906, 23, 147–188. [Google Scholar]
- Fernandes, L.H.; Mendonça, M.C. Collembola Poduromorpha do litoral de Maricá, Rio de Janeiro, Brasil. Rev. Bras. Zool. 2004, 21, 15–25. [Google Scholar] [CrossRef]
- Arlé, R. Sôbre Pseudachorutidae da região amazônica (Collembola). At. Soc. Biol. Rio J. 1962, 6, 1–3. [Google Scholar]
- Queiroz, G.C.; Weiner, W.M. A new species of Brachystomella (Collembola: Brachystomellidae) from the Atlantic Forest of southeast Brazil. Zootaxa 2011, 2885, 65–68. [Google Scholar] [CrossRef]
- Bellini, B.; Santos, N.; Souza, P.; Weiner, W. Two new species of Brazilian springtails (Hexapoda: Collembola) with comments on Neotropical Brachystomella Ågren and Seira (Lepidocyrtinus) Börner. Insect Syst. Evol. 2019, 50, 297–326. [Google Scholar] [CrossRef]
- Dos Santos-Rocha, I.M.; Andreazze, R.; Bellini, B.C. Registros de Collembola (Arthropoda, Hexapoda) no Estado do Rio Grande do Norte, Brasil. Biota Neotrop. 2011, 11, 167–170. [Google Scholar] [CrossRef][Green Version]
- Fernandes, L.H.; Mendonça, M.C. A new species of Xenylla (Collembola: Poduromorpha: Hypogastruridae) with new records for the state of Espirito Santo, southeastern Brazil. Zoologia 2010, 27, 123–127. [Google Scholar] [CrossRef]
- Da Silveira, T.C.; de Mendonça, M.C. Checklist of Poduromorpha (Collembola: Hexapoda) from Rio de Janeiro, Brazil. Zootaxa 2019, 4656, 572–590. [Google Scholar] [CrossRef] [PubMed]
- Brito, R.A.; Lima, E.C.A.; Ferreira, A.S.; Oliveira, J.V.L.C.; Amorim, T.P.L.; Machado, T.G.; Lopes, B.C.H.; Costa, I.A.; Alves, J.L.S.; Brito, N.P.; et al. A Catalogue of Species of Collembola (Hexapoda: Ellipura) Deposited in Coleção de Referência de Fauna de Solo of the Universidade Estadual da Paraíba, Brazil. Fla. Entomol. 2017, 100, 9–14. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H.; Abrantes, E.A.; Queiroz, G.C.; Bernardo, A.N.; Silveira, T.C. Fauna colembológica do Estado do Rio de Janeiro, Brasil. Arq. Mus. Nac. 2009, 67, 265–274. [Google Scholar]
- Arlé, R. Collembola Arthropleona do Brasil oriental e central. Arq. Mus. Nac. 1959, 49, 155–211. [Google Scholar]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of this Group of Insects. Families: Neogastruridae and Brachystomellidae. Acta Monogr. Musei Hist. Nat. Kraków 1949, 2, 1–341. [Google Scholar]
- Arlé, R. Collemboles nouveaux de Rio de Janeiro. Ann. Acad. Bras. Ciênc. 1939, 11, 25–32. [Google Scholar]
- De Mendonça, M.C.; Fernandes, L.H.; Abrantes, E.A. Two new species of Folsomiella Bonet and redescription of F. albida (Arlé, 1959) (Collembola, Brachystomellidae). Zootaxa 2005, 1006, 11. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H. A new genus of Brachystomellinae from Brazil (Collembola: Neanuridae). Bol. Mus. Nac. 1997, 379, 1–7. [Google Scholar]
- Queiroz, G.C.; Mendonca, M.C. New species of Brachystomellidae and characterization of Micronella porcus (Denis, 1933) from Brazil. ZooKeys 2013, 316, 81–98. [Google Scholar] [CrossRef]
- Mendonça, M.C.; Fernandes, L.H. Nova espécie de Rapoportella Ellis & Bellinger, 1973 do Brasil (Collembola, Neanuridae). Bol. Mus. Nac. 1995, 360, 1–6. [Google Scholar]
- Palacios-Vargas, J.G.; Gnaspini-Netto, P. A new Brazilian species of Acherontides (Collembola: Hypogastruridae), with notes on its ecology. Jour. Kan. Entom. Soc. 1992, 65, 443–447. [Google Scholar]
- De Lima, E.C.A.; Stievano, L.C.; Zeppelini, D. A new species of Acherontides (Collembola: Hypogastruridae) from Brazil, with an identification key. Zootaxa 2019, 4623, 141–150. [Google Scholar] [CrossRef]
- Culik, M.P.; Martins, D.D.S.; Ventura, J.A. Collembola (Arthropoda: Hexapoda) communities in the soil of papaya orchards managed with conventional and integrated production in Espírito Santo, Brazil. Biota Neotrop. 2006, 6, 1–8. [Google Scholar] [CrossRef]
- Fernandes, L.H.; Bellini, B.C.; de Mendonça, M.C. Revision of the genus Austrogastrura Thibaud & Palacios-Vargas, 1999 (Collembola: Poduromorpha: Hypogastruridae). Zootaxa 2010, 2542, 61–68. [Google Scholar] [CrossRef]
- Arlé, R. Collembola, anexo N. 2, ao relatório da excursão científica do Instituto Oswaldo Cruz realizada na zona da E. F. N. O. B., em outubro de 1938. Boletim Biol. 1939, 4, 295–300. [Google Scholar]
- Arlé, R. Uma nova espécie de Onychiurus (Collembola-Onychiuridae) de ocorrência periódica em Belém (Pará). Bol. Mus. Para. Emilio Goeldi 1970, 72, 1–11. [Google Scholar]
- Thibaud, J.-M.; Palacios-Vargas, J.G. Brazilian Collembola from littoral sand with description of Austrogastrura gen. n. and Isotomodes carioca sp. n. [Hypogastruridae; Isotomidae]. Rev. Franç. Entom. 1999, 21, 25–31. [Google Scholar]
- Moniez, R. Isotoma pallida, collembole nouveau de Brésil. Rev. Biol. Nord Fr. 1984, 6, 354. [Google Scholar]
- Queiroz, G.; Mendonça, M. New Hypogastruridae (Collembola) and first record of Cephalachorutes (Collembola: Neanuridae) for the Neotropical Region. Insect Syst. Evol. 2016, 47, 33–52. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.; Cipola, N.; Bellini, B. Two new species of Ceratophysella (Collembola: Hypogastruridae) from Brazil with a key to South American species of the genus. Insect Syst. Evol. 2018, 49, 207–230. [Google Scholar] [CrossRef]
- Denis, J.R. Sur Isotoma pallida Moniez et un autre collembole nouveau du Brésil: Hypogastrura moniezi n. sp. Bull. Soc. Zool. Fr. 1923, 48, 95–98. [Google Scholar]
- Fernandes, L.H.; Mendonça, M.C. Collembola Poduromorpha de áreas preservadas e impactadas do litoral sudeste do Brasil. Rev. Bras. Zool. 2007, 24, 777–785. [Google Scholar] [CrossRef]
- Queiroz, G.C.; Deharveng, L. Description of a new species of Paraxenylla (Collembola, Hypogastruridae) from Brazil, with comments on the genus. Zootaxa 2008, 1858, 1–8. [Google Scholar] [CrossRef]
- Fernandes, L.H.; Mendonça, C. Uma nova espécie de Schoettella Schaeffer (Collembola, Hypogastruridae). Rev. Bras. Entom. 1998, 41, 453–457. [Google Scholar]
- Oliveira, E.P.; Thibaud, J.-M. Un nouveau genre d’insecte collembole Hypogastruridae d’Amazonie. Amazoniana 1988, 10, 299–302. [Google Scholar]
- Gama, M.M. Systématique évolutive des Xenylla. X. Espèces provenant du Brésil, du Vietnam et de Madagscar (Insecta: Collembola). Ciênc. Biol. 1978, 4, 45–48. [Google Scholar]
- Neves, A.C.R.; Mendonça, M.C. A new species of Xenylla Tullberg, 1869 (Collembola: Hypogastruridae) from Central Amazon, Brazil. Zootaxa 2017, 4347, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gama, M.M.; Oliveira, E.P. Systématique évolutive des Xenylla. XVI. Description d’une nouvelle espèce provenant d’Amazonie (Insecta: Collembola). Amazoniana 1994, 13, 205–208. [Google Scholar]
- Da Silveira, T.C.; de Mendonça, M.C.; Da-Silva, E.R. A second new species of Isotogastrura Thibaud & Najt (Collembola: Isotogastruridae) from Brazil. J. Insect Biodivers. 2014, 2, 1–6. [Google Scholar] [CrossRef][Green Version]
- Arlé, R. Conspecto das espécies brasileiras de Pseudachorutinae, com descrição de uma espécie nova da Colômbia (Insecta, Collembola). Acta Amaz. 1981, 11, 583–593. [Google Scholar] [CrossRef]
- Fernandes, L.H.; Mendonça, M.C. Duas novas espécies de Pseudachorutinae (Collembola, Neanuridae) do Brasil. Bol. Mus. Nac. 2002, 496, 1–8. [Google Scholar]
- Da Paz, R.V.; Queiroz, G.C.; Bellini, B.C. A new species of Aethiopella Handschin, 1942 (Collembola, Poduromorpha, Neanuridae) from Neotropical Region, with comments on the genus. Zootaxa 2019, 4629, 39–50. [Google Scholar] [CrossRef]
- Arlé, R. Generalidades e importância ecológica da ordem Collembola (Apterygota). At. Soc. Biol. Rio J. 1959, 3, 4–7. [Google Scholar]
- Schuster, R. Über die Ökologie und Artengliederung der thalassobionten Collembolenfauna Brasiliens1. Beitr. Neotropischen Fauna 1965, 4, 191–208. [Google Scholar] [CrossRef]
- Arlé, R. Un nouveau collembole marin du littoral brésilien. Rev. Bras. Biol. 1966, 26, 367–372. [Google Scholar]
- Zeppelini, D.; Bellini, B.C.; Creão-Duarte, A.J.; Hernández, M.I.M. Collembola as bioindicators of restoration in mined sand dunes of Northeastern Brazil. Biodivers. Conserv. 2008, 18, 1161–1170. [Google Scholar] [CrossRef]
- Arlé, R. Collemboles d’Amazonie, I. Poduromorphes nouveaux ou peu connus et notes biologiques sur Neotropiella carli (Denis). Bol. Mus. Para. Emilio Goeldi 1966, 60, 1–19. [Google Scholar]
- Queiroz, G.; Deharveng, L. New Australonura Cassagnau 1980 (Neanuridae: Neanurinae) from Brazil. J. Nat. Hist. 2014, 48, 2293–2305. [Google Scholar] [CrossRef]
- Arlé, R. Novas espécies de Pseudachorutini (Collembola) do Rio de Janeiro e arredores. Bol. Biol. 1939, 4, 67–72. [Google Scholar]
- Zeppelini, D.; Queiroz, G.C.; Lopes, N.P.; Mendonça-Junior, F.J.B. Chemical analysis of Brasilimeria Stach, 1949 (Hexapoda, Collembola, Neanuridae) hemolymphatic secretion, and description of a new species. PLoS ONE 2019, 14, e0212451. [Google Scholar] [CrossRef]
- Arlé, R.; Rufino, E. Contribução ao conhecimento dos Pseudachorutinae da Amazônia (Collembola). Acta Amaz. 1976, 6, 99–107. [Google Scholar] [CrossRef]
- Arlé, R. Deux nouveaux collemboles du Brésil. Rev. Bras. Biol. 1943, 3, 109–112. [Google Scholar]
- Queiroz, G.C.; Deharveng, L. New genus, new species and new record of Neanurinae (Collembola, Neanuridae) for the Neotropics. Zootaxa 2015, 4020, 134–152. [Google Scholar] [CrossRef]
- Neves, A.C.; Mendonça, M.C.; Queiroz, G.C. Two new species and new records of Neanuridae (Hexapoda: Collembola) from Brazilian central Amazonia. Zoologi 2019, 36, 1–8. [Google Scholar] [CrossRef]
- Queiroz, G.C.; de Mendonça, M.C. Two new Brazilian species of Friesea Dalla Torre, 1895 (Collembola: Neanuridae). Austral. Entomol. 2014, 54, 110–116. [Google Scholar] [CrossRef]
- Silveira, T.C.; Mendonça, M.C. Two new species of Neanuridae (Collembola: Poduromorpha) from littoral of Rio de Janeiro, Brazil. Neotrop. Entomol. 2018, 47, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, G.C.; Fernandes, L.H. New Brazilian species of Furculanurida Massoud, 1967 (Collembola: Neanuridae). Zootaxa 2011, 2805, 57–64. [Google Scholar] [CrossRef]
- Queiroz, G.C.; Mendonça, M.C. Revalidation of Handschinia Stach, 1949 (Collembola, Neanuridae) with description of a new species from Brazil. Soil Org. 2018, 86, 169–173. [Google Scholar]
- Zeppelini, D.; Palacios-Vargas, J.G. A New Brazilian Species of Hylaeanura(Collembola: Neanuridae). Fla. Entomol. 2013, 96, 1401–1405. [Google Scholar] [CrossRef]
- Arlé, R. Notas sôbre a família Oncopoduridae, com descrição de duas espécies novas do Brasil (Collembola). Arq. Mus. Nac. 1960, 50, 9–23. [Google Scholar]
- Oliveira, E.P.; Deharveng, L. Response of soil Collembola (Insecta) communities to forest disturbance in central Amazonia (Brazil). In Functioning and Dynamics of Natural and Perturbed Ecosystems; Bellan, D., Bonin, G., Emig, C., Eds.; Technique et Documentation, Lavoisier, Intercept Ltd.: Paris, France, 1995; pp. 361–376. [Google Scholar]
- Queiroz, G.C.; Silveira, T.C.; Mendonça, M.C. New species of Neotropiella Handschin, 1942 (Collembola: Neanuridae) from Brazil. Soil Org. 2013, 85, 41–49. [Google Scholar]
- Thibaud, J.M.; Oliveira, E.P. Note sur les Collemboles de L’Amazonie Brezilienne II—Neanuridae: Pseudachorutinae ad. p. avec la description de deux espèces nouvelles. Rev. Franç. d’Entom. 2011, 32, 135–140. [Google Scholar]
- Cassagnau, P.; Oliveira, E.P. Les collemboles Neanurinae d’Amérique du Sud. Bull. Soc. d’Hist. Natur. Toulouse 1990, 126, 19–23. [Google Scholar]
- Oliveira, E.P.; Deharveng, L. Deux nouvelles especes de Pseudachorutes d’Amazonie (Collemboles, Neanuridae). Bull. Soc. Entom. Fr. 1994, 99, 389–395. [Google Scholar]
- Börner, C. Voläufige Mitteilung über einige neue Aphorurinen und zur Systematik der Collembola. Zool. Anz. 1901, 633, 1–15. [Google Scholar]
- Mendonça, M.C.; Silveira, T.C. A new species of Tijucameria from Brazil (Collembola: Neanuridae: Pseudachorutinae). Soil Org. 2012, 84, 563–568. [Google Scholar]
- Mendonça, M.C.; Fernandes, L.H. Novo gênero e nova espécie de Pseudachorutini do sudeste do Brasil (Collembola, Neanuridae, Pseudachorutinae). Rev. Bras. Zool. 2005, 22, 699–701. [Google Scholar] [CrossRef][Green Version]
- Arlé, R. Collemboles d’Amazonie, II. Pseudostachia folsomi gen. n. sp. n., intéressante espèce euédaphique. Bol. Mus. Para. Emilio Goeldi 1968, 68, 1–7. [Google Scholar]
- Oliveira, E.; Thibaud, J.-M. Notes sur les Collemboles de l’Amazonie, Brésil. 1. Hypogastruridae et Onychiuridae, avec la desciption de deux espèces nouvelles (Collembola). Opu. Zool. Flum. 1992, 95, 1–8. [Google Scholar]
- Rufino, E.O.; Schubart, H.O.R. Uma experiência sobre a nutrição de Onychiurus cunhai Arlé, 1970 (Insecta, Collembola), com notas sobre sua ecologia. Acta Amaz. 1974, 4, 53–55. [Google Scholar] [CrossRef][Green Version]
- Palacios-Vargas, J.G.; Díaz, M. Survey of the Onychiuridae (Collembola) from the Neotropical region. Ana. Inst. Biol. Univ. Nac. Aut. Mexico 1996, 66, 165–180. [Google Scholar]


























Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).