
The Impact of Climate Change on Agricultural Insect Pests




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Climate under Change
2.1. Impact of Climate Change on Crop Production
2.1.1. Impact of Temperature Increase
2.1.2. Impact of Elevated CO2 Concentration
2.1.3. Impact of Changeable Precipitation Pattern
2.2. Impact of Climate Change on Insect Pests
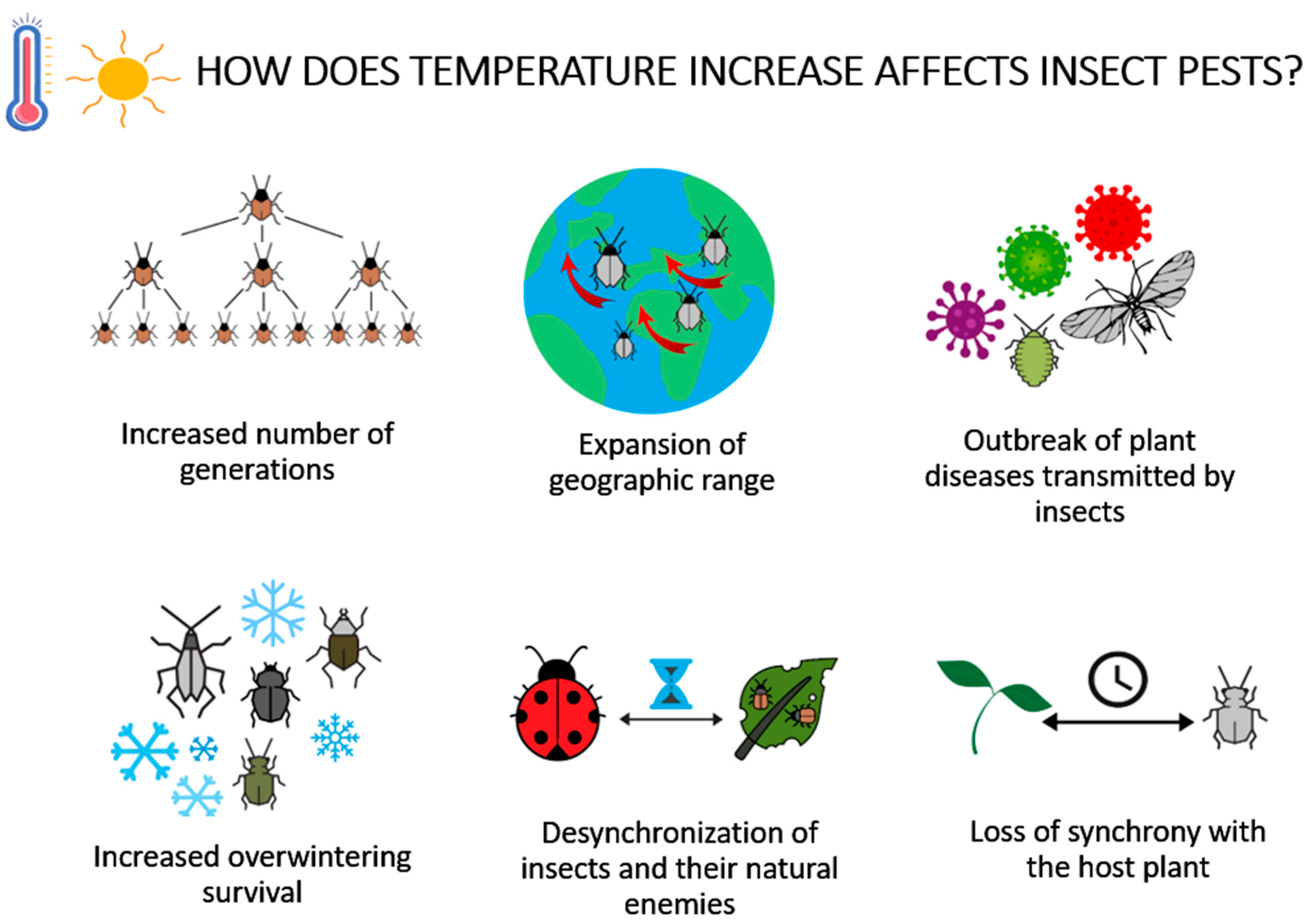
2.2.1. Response of Insect Pests to Increased Temperature
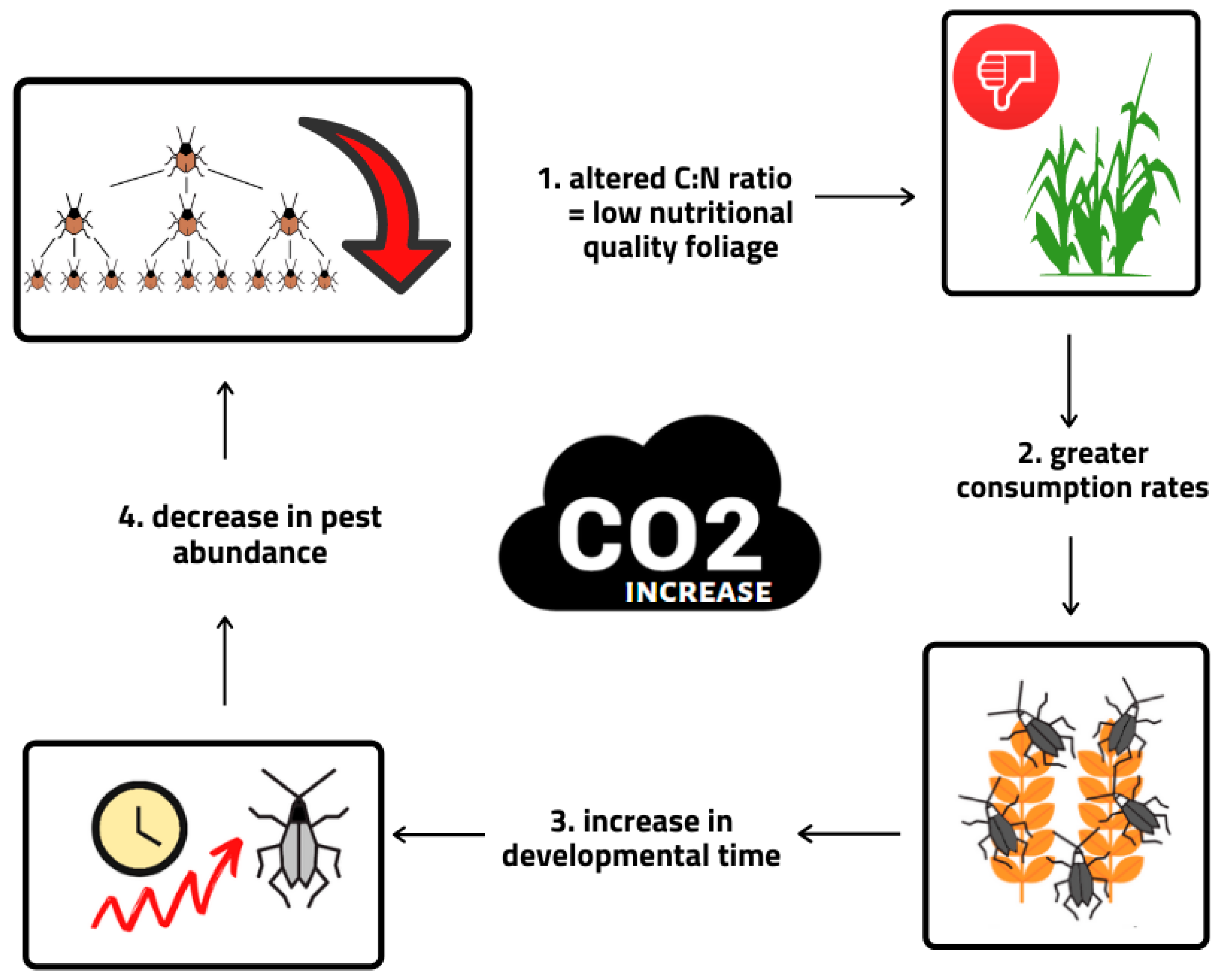
2.2.2. Response of Insect Pests to Increased CO2 Concentration
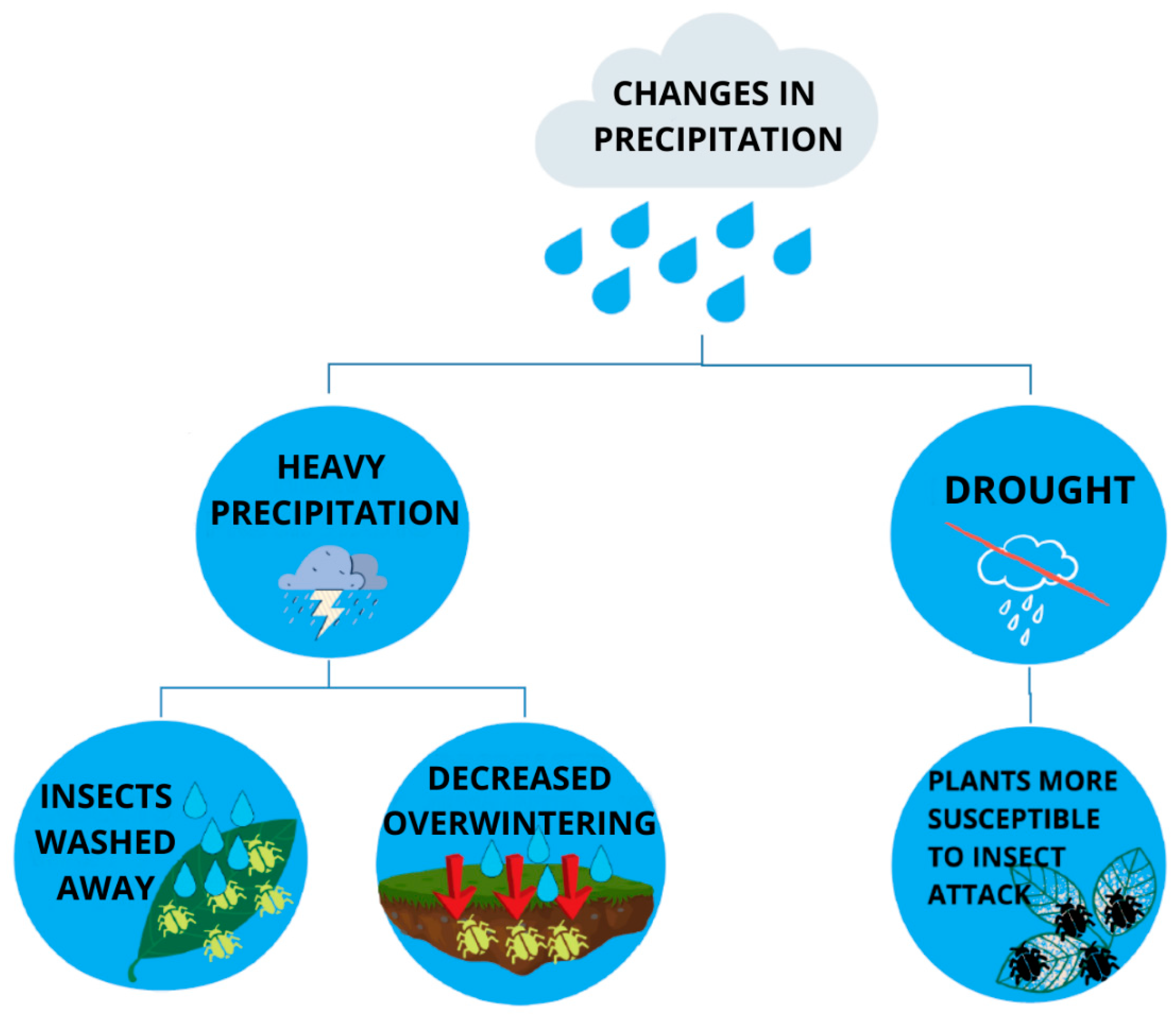
2.2.3. Response of Insect Pests to Changeable Precipitation Pattern
2.2.4. Expansion of Insects’ Distribution
2.2.5. Increased Overwintering Survival
2.2.6. Increased Number of Generations
2.2.7. Increased Risk of Invasive Alien Insect Species
2.2.8. Reduced Effectiveness of Biological Control Agents—Natural Enemies
2.2.9. Increased Incidence of Plant Diseases Transmitted by Insect Vectors
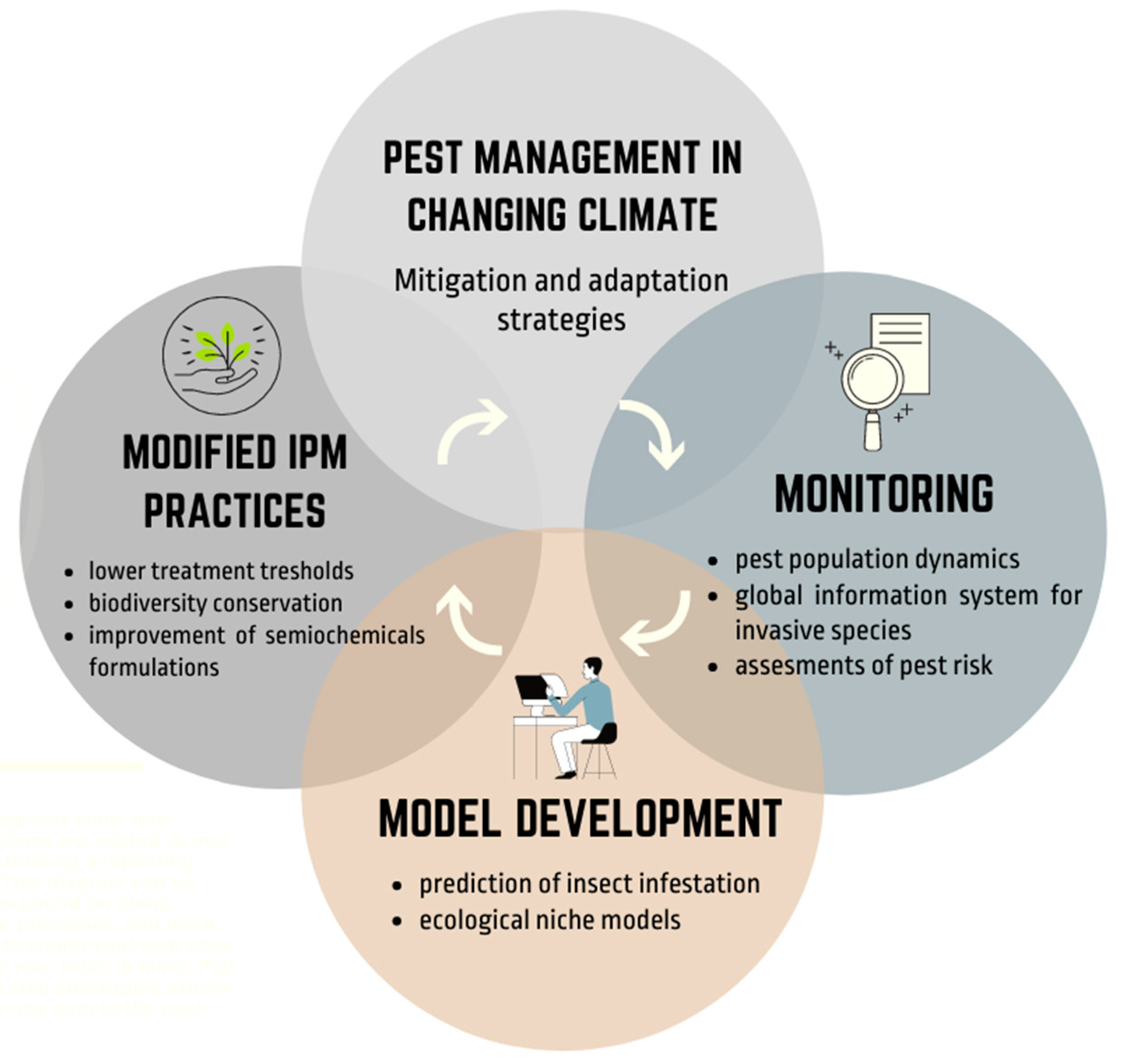
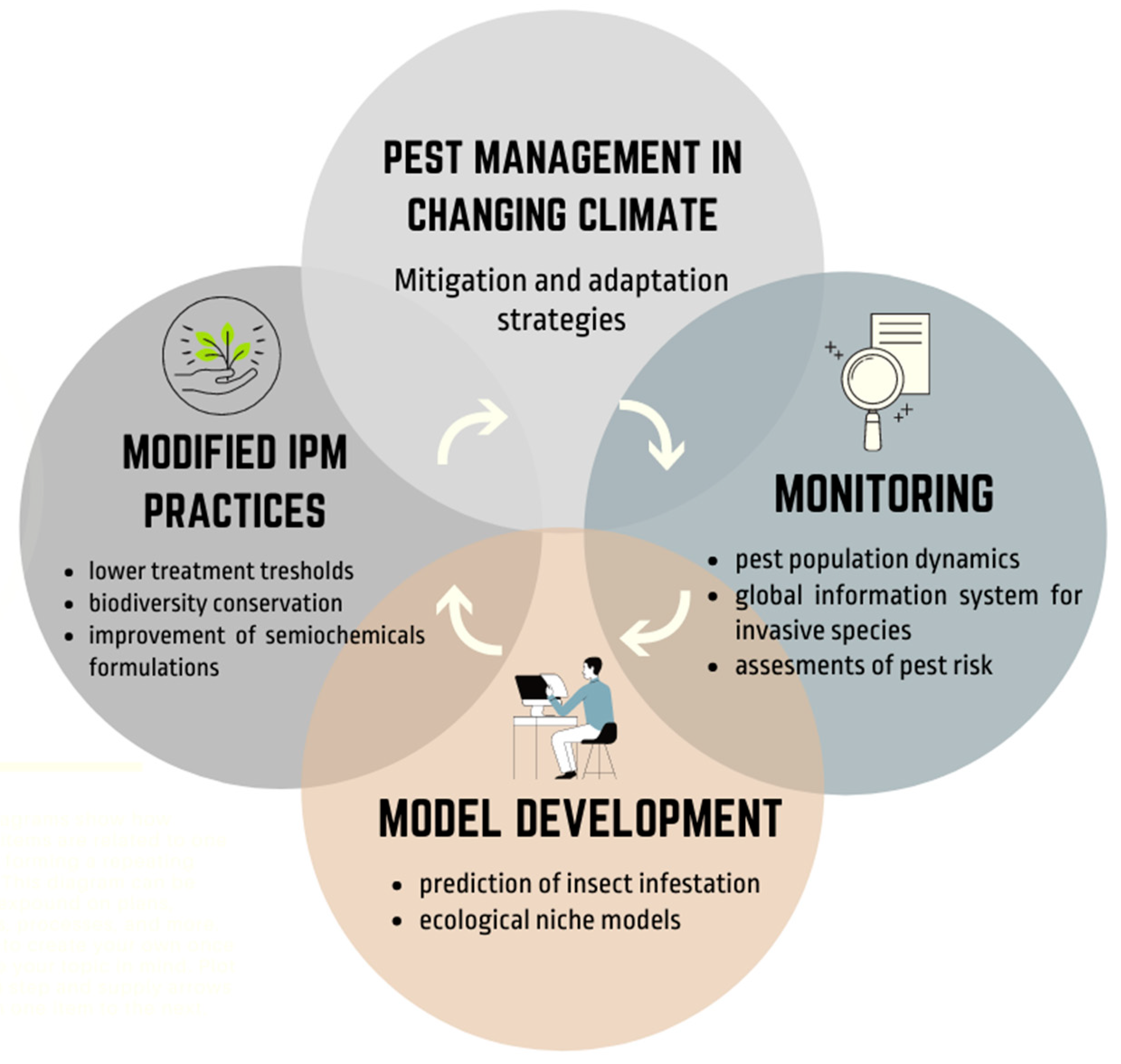
3. Adaptation and Mitigation Strategies for Pest Management in a Changing Climate
3.1. Modified Integrated Pest Management (IPM) Practices
3.2. Monitoring Abundance and Distribution
3.3. Climate Forecasting and Model Development
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Parry, M. The potential impact on agriculture of the greenhouse effect. Land Use Policy 1990, 7, 109–123. [Google Scholar] [CrossRef]
- Prakash, A.; Rao, J.; Mukherjee, A.K.; Berliner, J.; Pokhare, S.S.; Adak, T.; Munda, S.; Shashank, P.R. Climate Change: Impact on Crop Pests; Applied Zoologists Research Association (AZRA), Central Rice Research Institute: Odisha, India, 2014; ISBN 81-900947-2-7. [Google Scholar]
- Rosenzweig, C.; Major, D.C.; Demong, K.; Stanton, C.; Horton, R.; Stults, M. Managing climate change risks in New York City’s water system: Assessment and adaptation planning. Mitig. Adapt. Strat. Glob. Chang. 2007, 12, 1391–1409. [Google Scholar] [CrossRef]
- Shrestha, S. Effects of climate change in agricultural insect pest. Acta Sci. Agric. 2019, 3, 74–80. [Google Scholar] [CrossRef]
- Field, C.B.; Barros, V.R.; Dokken, D.J.; Mach, K.J.; Mastrandrea, M.D.; Bilir, T.E.; Chatterjee, M.; Ebi, K.L.; Estrada, Y.O.; Genova, R.C.; et al. IPCC Summary for policymakers. In Climate Change 2014: Impacts, Adaptation, and Vulnerability, Part A: Global and Sectoral Aspects; Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1–32. ISBN 978-92-9169-143-2. [Google Scholar]
- Pachauari, R.K.; Reisinger, A. Climate Change 2007: Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment Report on Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2007; ISBN 92-9169-122-4. [Google Scholar]
- Yoro, K.O.; Daramola, M.O. Chapter 1—CO2 emission sources, greenhouse gases, and the global warming effect. In Advances in Carbon Capture, 1st ed.; Rahimpour, M.R., Farsi, M., Makarem, M.A., Eds.; Woodhead Publishing: Sawston, UK, 2020; pp. 3–28. ISBN 9780128196571. [Google Scholar]
- Rogelj, J.D.; Shindell, K.; Jiang, S.; Fifita, P.; Forster, V.; Ginzburg, C.; Handa, H.; Kheshgi, S.; Kobayashi, E.; Kriegler, E.; et al. Mitigation pathways compatible with 1.5 °C in the context of sustainable development. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable De-velopment, and Efforts to Eradicate Poverty; Masson-Delmotte, V.P., Zhai, H.-O., Pörtner, D., Roberts, J., Skea, P.R., Shukla, A., Pirani, W., Moufouma-Okia, C., Péan, R., Pidcock, S., et al., Eds.; WMO: Geneva, Switzerland, 2018; ISBN 978-92-9169-151-7. [Google Scholar]
- Rosenzweig, C. Global climate change: Predictions and observations. Am. J. Agric. Econ. 1989, 71, 1265–1271. [Google Scholar] [CrossRef]
- Prentice, I.C.; Farquhar, G.D.; Fasham, M.J.R.; Goulden, M.L.; Heimann, M.; Jaramillo, V.J.; Kheshgi, H.S.; Le Quéré, C.; Scholes, R.J.; Wallace, D.W.R. The carbon cycle and atmospheric carbon dioxide. In Climate Change 2001: The Scientific Basis; Houghton, J.T., Ding, Y., Griggs, D.J., Noguer, M., van der Linden, P.J., Dai, X., Maskell, K., Johnson, C.A., Eds.; Cambridge University Press: New York, NY, USA, 2001; pp. 185–237. ISBN 0521807670. [Google Scholar]
- Global Monitioring Division, Earth System Research Laboratory. Trends in Atmospheric Carbon Dioxide. Available online: http://www.esrl.noaa.gov/gmd/ccgg/trends/ (accessed on 1 July 2011).
- Mahlman, J.D. Uncertainties in projections of human-caused climate warming. Science 1997, 278, 1416–1417. [Google Scholar] [CrossRef] [Green Version]
- Streck, N.A. Climate change and agroecosystems: The effect of elevated atmospheric CO2 and temperature on crop growth, development, and yield. Ciênc. Rural 2005, 35, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Deshar, R.; Koirala, M. Climate Change and Gender Policy. In Global Climate Change and Environmental Policy: Agriculture Perspectives, 1st ed.; Venkatramanan, V., Shah, S., Prasad, R., Eds.; Springer Nature Pte Ltd.: Singapore, 2019; pp. 411–422. [Google Scholar]
- Müller, C. African lessons on climate change risks for agriculture. Annu. Rev. Nutr. 2013, 33, 395–411. [Google Scholar] [CrossRef]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of nitrogen deposition on the species richness of grasslands. Science 2004, 303, 1876–1879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nat. Cell Biol. 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Rehman, M.U.; Rather, G.H.; Gull, Y.; Mir, M.R.; Mir, M.M.; Waida, U.I.; Hakeem, K.R. Effect of climate change on horticultural crops. In Crop Production and Global Environmental Issues; Hakeem, K.R., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 211–239. [Google Scholar]
- Luo, Q. Temperature thresholds and crop production: A review. Clim. Chang. 2011, 109, 583–598. [Google Scholar] [CrossRef]
- Tubiello, F.; Rosenzweig, C.; Goldberg, R.; Jagtap, S.; Jones, J. Effects of climate change on US crop production: Simulation results using two different GCM scenarios. Part I: Wheat, potato, maize, and citrus. Clim. Res. 2002, 20, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Rötter, R.; Van De Geijn, S. Climate change effects on plant growth, crop yield and livestock. Clim. Chang. 1999, 43, 651–681. [Google Scholar] [CrossRef]
- Woodward, F.I. Global change: Translating plant ecophysiological responses to ecosystems. Trends Ecol. Evol. 1990, 5, 308–311. [Google Scholar] [CrossRef]
- Drake, B.G.; Gonzàlez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Annu. Rev. Plant Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenzweig, C.; Hillel, D. Climate Change and the Global Harvest. Potential Impacts of the Greenhouse Effect on Agriculture; Oxford University Press Inc.: New York, NY, USA, 1998; ISBN 0195088891. [Google Scholar]
- Bloom, A.J.; Asensio, J.S.R.; Randall, L.; Rachmilevitch, S.; Cousins, A.B.; Carlisle, E.A. CO2enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 2012, 93, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R.; Cerling, T.E.; Dearing, M.D. Atmospheric CO2 as a global change driver influencing plant-animal interactions. Integr. Comp. Biol. 2002, 42, 424–430. [Google Scholar] [CrossRef]
- Leegood, R.C. Photosynthesis. In Encyclopedia of Biological Chemistry, 1st ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press: London, UK, 2004; pp. 330–335. ISBN 9780124437104. [Google Scholar]
- Wang, C.; Guo, L.; Chuanli, W.; Wang, Z. Systematic comparison of C3 and C4 plants based on metabolic network analysis. BMC Syst. Biol. 2012, 6, S9. [Google Scholar] [CrossRef] [Green Version]
- Majeran, W.; Friso, G.; Ponnala, L.; Connolly, B.; Huang, M.; Reidel, E.; Zhang, C.; Asakura, Y.; Bhuiyan, N.H.; Sun, Q.; et al. Structural and metabolic transitions of C4 leaf development and differentiation defined by microscopy and quantitative proteomics in maize. Plant Cell 2010, 22, 3509–3542. [Google Scholar] [CrossRef] [Green Version]
- Cure, J.D.; Israel, D.W.; Rufty, T.W. Nitrogen stress effects on growth and seed yield of nonnodulated soybean exposed to elevated carbon dioxide. Crop Sci. 1988, 28, 671–677. [Google Scholar] [CrossRef]
- Olesen, J.E.; Bindi, M. Consequences of climate change for European agricultural productivity, land use and policy. Eur. J. Agron. 2002, 16, 239–262. [Google Scholar] [CrossRef]
- Huntington, T.G. Climate warming-induced intensification of the hydrologic cycle: An assessment of the published record and potential impacts on agriculture. Adv. Agron. 2010, 109, 1–53. [Google Scholar] [CrossRef]
- Lickley, M.; Solomon, S. Drivers, timing and some impacts of global aridity change. Environ. Res. Lett. 2018, 13, 104010. [Google Scholar] [CrossRef]
- Kirda, C.; Topcu, S.; Cetin, M.; Dasgan, H.; Kaman, H.; Topaloglu, F.; Derici, M.; Ekici, B. Prospects of partial root zone irrigation for increasing irrigation water use efficiency of major crops in the Mediterranean region. Ann. Appl. Biol. 2007, 150, 281–291. [Google Scholar] [CrossRef]
- Zayan, S.A. Impact of climate change on plant diseases and IPM strategies. In Plant Pathology and Management of Plant Diseases; Topolovec-Pintarić, S., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-78985-116-8. [Google Scholar]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Tank, A.M.G.K.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, F.; et al. Global observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. Phys. 2006, 111, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Min, S.-K.; Zhang, X.; Zwiers, F.W.; Hegerl, G.C. Human contribution to more-intense precipitation extremes. Nature 2011, 470, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Brignall, A.P.; Rounsevel, M.D.A. Land evaluation modelling to assess the effects of climate change on winter wheat potential in England and Wales. J. Agric. Sci. 1995, 124, 159–172. [Google Scholar] [CrossRef]
- Kocmánková, E.; Trnka, M.; Juroch, J.; Dubrovský, M.; Semerádová, D.; Možný, M.; Žalud, Z.; Pokorný, R.; Lebeda, A. Impact of climate change on the occurrence and activity of harmful organisms. Plant Prot. Sci. 2010, 45, S48–S52. [Google Scholar] [CrossRef] [Green Version]
- Fand, B.B.; Kamble, A.L.; Kumar, M. Will climate change pose serious threat to crop pest management: A critical review. Int. J. Sci. Res. 2012, 2, 1–14. [Google Scholar]
- FAO. Climate Related Transboundary Pests and Diseases. Available online: http://www.fao.org/3/a-ai785e.pdf (accessed on 19 December 2020).
- Hare, J.D. Effects of plant variation on herbivore-natural enemy interactions. In Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and Genetics, 1st ed.; Fritz, R.S., Simms, E.L., Eds.; University of Chicago Press: Chicago, IL, USA, 1992; pp. 278–298. ISBN 9780226265544. [Google Scholar]
- Caulfield, F.; Bunce, J.A. Elevated atmospheric carbon dioxide concentration affects interactions between Spodoptera exigua (Lepidoptera: Noctuidae) larvae and two host plant species outdoors. Environ. Entomol. 1994, 23, 999–1005. [Google Scholar] [CrossRef]
- Roth, S.K.; Lindroth, R.L. Elevated atmospheric CO2: Effects on phytochemistry, insect performance and insect-parasitoid interactions. Glob. Chang. Biol. 1995, 1, 173–182. [Google Scholar] [CrossRef]
- Dukes, J.S.D.S.; Pontius, J.; Orwig, D.; Garnas, J.R.G.R.; Rodgers, V.L.; Brazee, N.; Cooke, B.; Theoharides, K.A.T.A.; Stange, E.E.S.E.; Harrington, R.; et al. Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: What can we predict? This article is one of a selection of papers from NE Forests 2100: A Synthesis of Climate Change Impacts on Forests of the Northeastern US and Eastern Canada. Can. J. For. Res. 2009, 39, 231–248. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- DeLucia, E.H.; Casteel, C.L.; Nabity, P.D.; O’Neill, B.F. Insects take a bigger bite out of plants in a warmer, higher carbon dioxide world. Proc. Natl. Acad. Sci. USA 2008, 105, 1781–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [Green Version]
- Awmack, C.; Woodcock, C.; Harrington, R. Climate change may increase vulnerability of aphids to natural enemies. Ecol. Entomol. 1997, 22, 366–368. [Google Scholar] [CrossRef]
- Pathania, M.; Verma, A.; Singh, M.; Arora, P.K.; Kaur, N. Influence of abiotic factors on the infestation dynamics of whitefly, Bemisia tabaci (Gennadius 1889) in cotton and its management strategies in North-Western India. Int. J. Trop. Insect Sci. 2020, 40, 969–981. [Google Scholar] [CrossRef]
- Johansen, B.E. The Global Warming Desk Reference; Greenwood Publishing Group: Westport, CT, USA, 2002; ISBN 0-313-31679-1. [Google Scholar]
- Karl, T.R.; Trenberth, K.E. Modern global climate change. Science 2003, 302, 1719–1723. [Google Scholar] [CrossRef] [Green Version]
- Collins, W.; Colman, R.; Haywood, J.; Manning, M.R.; Mote, P. the physical science behind climate change. Sci. Am. 2007, 297, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; De Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Hance, T.; Van Baaren, J.; Vernon, P.; Boivin, G. Impact of extreme temperatures on parasitoids in a climate change perspective. Annu. Rev. Entomol. 2007, 52, 107–126. [Google Scholar] [CrossRef]
- Fuhrer, J. Agroecosystem responses to combinations of elevated CO2, ozone, and global climate change. Agric. Ecosyst. Environ. 2003, 97, 1–20. [Google Scholar] [CrossRef]
- Coviella, C.E.; Trumble, J.T. Effects of elevated atmospheric carbon dioxide on insect-plant interactions. Conserv. Biol. 1999, 13, 700–712. [Google Scholar] [CrossRef]
- Lincoln, D.E.; Sionit, N.; Strain, B.R. Growth and feeding response of Pseudoplusia includens (Lepidoptera: Noctuidae) to host plants grown in controlled carbon dioxide atmospheres. Environ. Entomol. 1984, 13, 1527–1530. [Google Scholar] [CrossRef]
- Lincoln, D.E. The influence of plant carbon dioxide and nutrient supply on susceptibility to insect herbivores. Vegetatio 1993, 104, 273–280. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ineson, P.; Scott, A. Elevated CO2 reduces the nitrogen concentration of plant tissues. Glob. Chang. Biol. 1998, 4, 43–54. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Jones, T.H.; Knight, K.J. Long-term effects of elevated CO2 and temperature on populations of the peach potato aphid Myzus persicae and its parasitoid Aphidius matricariae. Oecologia 1998, 116, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.G.; Dermody, O.; Aldea, M.; Zangerl, A.R.; Rogers, A.; Berenbaum, M.R.; DeLucia, E.H. Anthropogenic changes in tropospheric composition increase susceptibility of soybean to insect herbivory. Environ. Entomol. 2005, 34, 479–485. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Kinney, K.K.; Platz, C.L. Responses of diciduous trees to elevated atmospheric CO2: Productivity, phytochemistry, and insect performance. Ecology 1993, 74, 763–777. [Google Scholar] [CrossRef]
- Thomson, L.J.; Macfadyen, S.; Hoffmann, A.A. Predicting the effects of climate change on natural enemies of agricultural pests. Biol. Control 2010, 52, 296–306. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Kozlov, M.V. Consequences of simultaneous elevation of carbon dioxide and temperature for plant-herbivore interactions: A metaanalysis. Glob. Chang. Biol. 2005, 12, 27–41. [Google Scholar] [CrossRef]
- Sutherst, R.W.; Constable, F.; Finlay, K.J.; Harrington, R.; Luck, J.; Zalucki, M.P. Adapting to crop pest and pathogen risks under a changing climate. Wiley Interdiscip. Rev. Clim. Chang. 2011, 2, 220–237. [Google Scholar] [CrossRef]
- Stiling, P.; Cornelissen, T. How does elevated carbon dioxide (CO2) affect plant–herbivore interactions? A field experiment and meta-analysis of CO2-mediated changes on plant chemistry and herbivore performance. Glob. Chang. Biol. 2007, 13, 1823–1842. [Google Scholar] [CrossRef]
- Hullé, M.; D’Acier, A.C.; Bankhead-Dronnet, S.; Harrington, R. Aphids in the face of global changes. C. R. Biol. 2010, 333, 497–503. [Google Scholar] [CrossRef]
- Sun, Y.; Ge, F. How do aphids respond to elevated CO2? J. Asia-Pac. Entomol. 2011, 14, 217–220. [Google Scholar] [CrossRef]
- Pathak, H.; Aggarwal, P.K.; Singh, S.D. Climate Change Impact, Adaptation and Mitigation in Agriculture: Methodology for Assessment and Applications; Indian Agricultural Research Institute: New Delhi, India, 2012; ISBN 978-81-88708-82-6. [Google Scholar]
- Staley, J.T.; Hodgson, C.J.; Mortimer, S.R.; Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Taylor, M.E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. Eur. J. Soil Biol. 2007, 43, 189–198. [Google Scholar] [CrossRef]
- Johnson, S.N.; Anderson, E.A.; Dawson, G.; Griffiths, D.W. Varietal susceptibility of potatoes to wireworm herbivory. Agric. For. Entomol. 2008, 10, 167–174. [Google Scholar] [CrossRef]
- Gregory, P.J.; Johnson, S.N.; Newton, A.C.; Ingram, J.S.I. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 2009, 60, 2827–2838. [Google Scholar] [CrossRef]
- Yihdego, Y.; Salem, H.S.; Muhammed, H.H. Agricultural pest management policies during drought: Case studies in Australia and the state of Palestine. Nat. Hazards Rev. 2019, 20, 05018010. [Google Scholar] [CrossRef]
- Ezcurra, E.; Rapoport, E.H.; Marino, C.R. The geographical distribution of insect pests. J. Biogeogr. 1978, 5, 149. [Google Scholar] [CrossRef]
- Hill, D.S. Agricultural Insect Pests of Temperate Regions and Their Control; Cambridge University Press: New York, NY, USA, 1987; ISBN 0521240131. [Google Scholar]
- Meynard, C.N.; Migeon, A.; Navajas, M. Uncertainties in predicting species distributions under climate change: A case study using Tetranychus evansi (Acari: Tetranychidae), a widespread agricultural pest. PLoS ONE 2013, 8, e66445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laštůvka, Z. Climate change and its possible influence on the occurrence and importance of insect pests. Plant Prot. Sci. 2010, 45, S53–S62. [Google Scholar] [CrossRef] [Green Version]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Porter, J.; Parry, M.; Carter, T. The potential effects of climatic change on agricultural insect pests. Agric. For. Meteorol. 1991, 57, 221–240. [Google Scholar] [CrossRef]
- Lopez-Vaamonde, C.; Agassiz, D.; Augustin, S.; De Prins, J.; De Prins, W.; Gomboc, S.; Ivinskis, P.; Karsholt, O.; Koutroumpas, A.; Koutroumpa, F.; et al. Lepidoptera. Chapter 11. BioRisk 2010, 4, 603–668. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nat. Cell Biol. 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Coulson, S.J.; Hodkinson, I.D.; Webb, N.R.; Mikkola, K.; Harrison, J.A.; Pedgley, D.E. Aerial colonization of high Arctic islands by invertebrates: The diamondback moth Plutella xylostella (Lepidoptera: Yponomeutidae) as a potential indicator species. Divers. Distrib. 2002, 8, 327–334. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; D’Oultremont, T.; Ellis, C.; Ponti, L. Climatic limits of pink bollworm in Arizona and California: Effects of climate warming. Acta Oecol. 2006, 30, 353–364. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Cossu, Q.A. Prospective comparative analysis of global warming effects on olive and olive fly. (Bactrocera oleae (Gmelin)) in Arizona-California and Italy. Clim. Chang. 2009, 95, 195–217. [Google Scholar] [CrossRef]
- Fleming, R.A.; Volney, W.J.A. Effects of climate change on insect defoliator population processes in Canada’s boreal forest: Some plausible scenarios. Water Air Soil Pollut. 1995, 82, 445–454. [Google Scholar] [CrossRef]
- Raza, M.M.; Khan, M.A.; Arshad, M.; Sagheer, M.; Sattar, Z.; Shafi, J.; Haq, E.U.; Ali, A.; Aslam, U.; Mushtaq, A.; et al. Impact of global warming on insects. Arch. Phytopathol. Plant Prot. 2014, 48, 84–94. [Google Scholar] [CrossRef]
- Patterson, D.T.; Westbrook, J.K.; Joyce, R.; Lingren, P.D.; Rogasik, J. Weeds, insects, and diseases. Clim. Chang. 1999, 43, 711–727. [Google Scholar] [CrossRef]
- Pareek, A.; Meena, B.M.; Sharma, S.; Tetarwal, M.L.; Kalyan, R.K.; Meena, B.L. Impact of climate change on insect pests and their management strategies. In Climate Change and Sustainable Agriculture; Kumar, P.S., Kanwat, M., Meena, P.D., Kumar, V., Alone, R.A., Eds.; New India Publishing Agency: New Delhi, India, 2017; pp. 253–286. ISBN 9789385516726. [Google Scholar]
- González-Tokman, D.; Córdoba-Aguilar, A.; Dáttilo, W.; Lira-Noriega, A.; Sánchez-Guillén, R.A.; Villalobos, F. Insect responses to heat: Physiological mechanisms, evolution and ecological implications in a warming world. Biol. Rev. 2020, 95, 802–821. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A.L. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef] [Green Version]
- Taylor, L.R.; Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: Oxford, UK, 1986. [Google Scholar]
- Gill, H.K.; Goyal, G.; Chahil, G. Insect diapause: A review. J. Agric. Sci. Technol. A 2017, 7, 454–473. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects: Structure and Function, 4th ed.; Cambridge University Press: Cambridge, UK, 1998; ISBN 9780521113892. [Google Scholar]
- Pullin, A.; Bale, J. Influence of diapause and temperature on cryoprotectant synthesis and cold hardiness in pupae of Pieris brassicae. Comp. Biochem. Physiol. Part A Physiol. 1989, 94, 499–503. [Google Scholar] [CrossRef]
- Bradshaw, W.E.; Holzapfel, C.M. Light, time, and the physiology of biotic response to rapid climate change in animals. Annu Rev. Plant Biol. 2010, 72, 147–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McWatters, H.G.; Saunders, D.S. Maternal temperature has different effects on the photoperiodic response and duration of larval diapause in blow fly (Calliphora vicina) strains collected at two latitudes. Physiol. Entomol. 1998, 23, 369–375. [Google Scholar] [CrossRef]
- Musolin, D.L. Insects in a warmer world: Ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob. Chang. Biol. 2007, 13, 1565–1585. [Google Scholar] [CrossRef]
- Tougou, D.; Musolin, D.L.; Fujisaki, K. Some like it hot! Rapid climate change promotes changes in distribution ranges of Nezara viridula and Nezara antennata in Japan. Entomol. Exp. Appl. 2009, 130, 249–258. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Energetics of insect diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Glob. Chang. Biol. 2007, 13, 1860–1872. [Google Scholar] [CrossRef]
- Harrington, R.; Woiwod, I.; Sparks, T. Climate change and trophic interactions. Trends Ecol. Evol. 1999, 14, 146–150. [Google Scholar] [CrossRef]
- Harrington, R.; Fleming, R.A.; Woiwod, I.P. Climate change impacts on insect management and conservation in temperate regions: Can they be predicted? Agric. For. Entomol. 2001, 3, 233–240. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A. Direct impacts of recent climate warming on insect populations. Integr. Zool. 2010, 5, 132–142. [Google Scholar] [CrossRef]
- Bale, J.S. Classes of insect cold hardiness. Funct. Ecol. 1993, 7, 751–753. [Google Scholar]
- Yamamura, K.; Yokozawa, M. Prediction of a geographical shift in the prevalence of rice stripe virus disease transmitted by the small brown planthopper, Laodelphax striatellus (Fallen) (Hemiptera: Delphacidae), under global warming. Appl. Entomol. Zool. 2002, 37, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Kocmánková, E.; Trnka, M.; Eitzinger, J.; Dubrovský, M.; Štěpánek, P.; Semeradova, D.; Balek, J.; Skalak, P.; Farda, A.; Juroch, J.; et al. Estimating the impact of climate change on the occurrence of selected pests at a high spatial resolution: A novel approach. J. Agric. Sci. 2011, 149, 185–195. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Krupke, C.H.; White, M.A.; Alexander, C.E. Global warming presents new challenges for maize pest management. Environ. Res. Lett. 2008, 3, 044007. [Google Scholar] [CrossRef]
- Cannon, R.J. The implications of predicted climate change for insect pests in the UK, with emphasis on non-indigenous species. Glob. Chang. Biol. 1998, 4, 785–796. [Google Scholar] [CrossRef]
- Bale, J.S. Implications of cold hardiness for pest management. In Insects at Low Temperature; Lee, R.E., Ed.; Springer: New York, NY, USA, 1991; pp. 461–498. ISBN 978-1-4757-0190-6. [Google Scholar]
- Harrington, R.; Bale, J.S.; Tatchell, G.M. Aphids in a changing climate. In Insects in a Changing Environment; Harrington, R., Stork, N.E., Eds.; Academic Press: London, UK, 1995; pp. 125–155. [Google Scholar]
- Zhou, X.-L.; Harrington, R.; Woiwod, I.P.; Perry, J.N.; Bale, J.S.; Clark, S.J. Effects of temperature on aphid phenology. Glob. Chang. Biol. 1995, 1, 303–313. [Google Scholar] [CrossRef]
- Baker, R.-A.; Cannon, R.-C.; Walters, K.-A. An assessment of the risks posed by selected non-indigenous pests to UK crops under climate change. Asp. Appl. Biol. 1996, 45, 323–330. [Google Scholar]
- Menéndez, R.; González-Megías, A.; Collingham, Y.; Fox, R.; Roy, D.B.; Ohlemüller, R.; Thomas, C.D. Direct and indirect effects of climate and habitat factors on butterfly diversity. Ecology 2007, 88, 605–611. [Google Scholar] [CrossRef]
- Menéndez, R. How are insects responding to global warming? Tijdschr. Entomol. 2007, 150, 355. [Google Scholar]
- Yamamura, K.; Kiritani, K. A simple method to estimate the potential increase in the number of generations under global warming in temperate zones. Appl. Entomol. Zool. 1998, 33, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Cayton, H.L.; Haddad, N.M.; Gross, K.; Diamond, S.E.; Ries, L. Do growing degree days predict phenology across butterfly species? Ecology 2015, 96, 1473–1479. [Google Scholar] [CrossRef]
- Pollard, E.; Yates, T.J. Monitoring Butterflies for Ecology and Conservation: The British Butterfly Monitoring Scheme; Springer: Dordrecht, The Netherlands, 1993; ISBN 978-0-412-40220-3. [Google Scholar]
- Bernays, E.A. Feeding by lepidopteran larvae is dangerous. Ecol. Entomol. 1997, 22, 121–123. [Google Scholar] [CrossRef]
- Altermatt, F. Climatic warming increases voltinism in European butterflies and moths. Proc. R. Soc. B 2010, 277, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Harrington, R.; Clark, S.J.; Welham, S.J.; Verrier, P.J.; Denholm, C.H.; Hulle, M. Environmental change and the phenology of European aphids. Glob. Chang. Biol. 2007, 13, 1550–1564. [Google Scholar] [CrossRef]
- Roy, D.B.; Sparks, T.H. Phenology of British butterflies and climate change. Glob. Chang. Biol. 2000, 6, 407–416. [Google Scholar] [CrossRef]
- Stefanescu, C.; Penuelas, J.; Filella, I. Effects of climatic change on the phenology of butterflies in the northwest Mediterranean Basin. Glob. Chang. Biol. 2003, 9, 1494–1506. [Google Scholar] [CrossRef]
- Martín-Vertedor, D.; Ferrero-García, J.J.; Torres-Vila, L.M. Global warming affects phenology and voltinism of Lobesia botrana in Spain. Agric. For. Entomol. 2010, 12, 169–176. [Google Scholar] [CrossRef]
- Roff, D. Optimizing development time in a seasonal environment: The ‘ups and downs’ of clinal variation. Oecologia 1980, 45, 202–208. [Google Scholar] [CrossRef]
- Tobin, P.C.; Nagarkatti, S.; Loeb, G.; Saunders, M.C. Historical and projected interactions between climate change and insect voltinism in a multivoltine species. Glob. Chang. Biol. 2008, 14, 951–957. [Google Scholar] [CrossRef]
- Shine, C.; Williams, N.; Gündling, L. A Guide to Designing Legal and Institutional Frameworks on Alien Invasive Species, Environmental Policy and Law Paper No. 40; IUCN: Gland, Switzerland; Cambridge, UK; Bonn, Germany, 2000; ISBN 2-8317-0548-7. [Google Scholar]
- Ward, N.L.; Masters, G.J. Linking climate change and species invasion: An illustration using insect herbivores. Glob. Chang. Biol. 2007, 13, 1605–1615. [Google Scholar] [CrossRef]
- Ricciardi, A. Invasive species. In Ecological Systems, 1st ed.; Leemans, R., Ed.; Springer: New York, NY, USA, 2013; pp. 161–178. [Google Scholar] [CrossRef]
- CBD. Convention on Biological Diversity. Invasive alien species: The application of classical biological control for the management of established invasive alien species causing environmental impacts. In Proceedings of the Conference of the Parties to the Convention on Biological Diversity, 14th Meeting, Sharm El-Sheikh, Egypt, 17–29 November 2018. [Google Scholar]
- Vander Zanden, M.J. The success of animal invaders. Proc. Natl. Acad. Sci. USA 2005, 102, 7055–7056. [Google Scholar] [CrossRef] [Green Version]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, M.P.; Bertelsmeier, C.; Clusella-Trullas, S.; Garnas, J.; Robertson, M.P.; Terblanche, J.S. Predicted decrease in global climate suitability masks regional complexity of invasive fruit fly species response to climate change. Biol. Invasions 2016, 18, 1105–1119. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Blumenthal, D.M.; Runion, G.B.; Raymond Hunt, E., Jr.; Diaz-Soltero, H. Invasive species and climate change: An agronomic perspective. Clim. Chang. 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Masters, G.; Norgrove, L. Climate Change and Invasive Alien Species; CABI Working Paper 1; CABI: Wallingford, UK, 2010; p. 30. [Google Scholar]
- Vanhanen, H. Invasive insects in Europe—The role of climate change and global trade. Diss. For. 2008, 57, 33. [Google Scholar] [CrossRef] [Green Version]
- Tobin, P.C.; Parry, D.; Aukema, B.H. The influence of climate change on insect invasions in temperate forest ecosystems. For. Sci. 2014, 81, 267–293. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization Plant Pests and Diseases in the Context of Climate Change and Climate Variability, Food Security and Biodiversity Risks. Available online: http://www.fao.org/3/nb088/nb088.pdf (accessed on 12 January 2020).
- Hill, M.P.; Mafokoane, L.D.; Zimmermann, H.G. Development of Cactoblastis cactorum (Berg) (Lepidoptera: Pyralidae) on six north American Opuntia species. Afr. Entomol. 2007, 15, 295–299. [Google Scholar] [CrossRef]
- Burgiel, S.W.; Muir, A.A. Invasive Species, Climate Change and Ecosystem-Based Adaptation: Addressing Multiple Drivers of Global Change; Global Invasive Species Program (GISP): Washington, DC, USA, 2010; ISBN 9789290592877. [Google Scholar]
- Kiritani, K.; Yamamura, K. Exotic insects and their pathways for invasion. In Invasive Species: Vectors and Management Strategies; Ruiz, G.M., Carlton, J.T., Eds.; Island Press: Washington, DC, USA, 2003; pp. 44–67. ISBN 1559639032. [Google Scholar]
- Lockwood, J.L.; Cassey, P.; Blackburn, T.M. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Bacon, S.J.; Aebi, A.; Calanca, P.; Bacher, S. Quarantine arthropod invasions in Europe: The role of climate, hosts and propagule pressure. Divers. Distrib. 2014, 20, 84–94. [Google Scholar] [CrossRef]
- Rota-Stabelli, O.; Blaxter, M.; Anfora, G. Drosophila suzukii. Curr. Biol. 2013, 23, R8–R9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cini, A.; Anfora, G.; Escudero-Colomar, L.A.; Grassi, A.; Santosuosso, U.; Seljak, G.; Papini, A. Tracking the invasion of the alien fruit pest Drosophila suzukii in Europe. J. Pest. Sci. 2014, 87, 559–566. [Google Scholar] [CrossRef]
- Vermeij, G.J. An agenda for invasion biology. Biol. Conserv. 1996, 78, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Snell-Rood, E.C.; Kobiela, M.E.; Sikkink, K.L.; Shephard, A.M. Mechanisms of plastic rescue in novel environments. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 331–354. [Google Scholar] [CrossRef]
- Fox, R.J.; Donelson, J.M.; Schunter, C.; Ravasi, T.; Gaitán-Espitia, J.D. Beyond buying time: The role of plasticity in phenotypic adaptation to rapid environmental change. Phil. Trans. R. Soc. B 2019, 374, 20180174. [Google Scholar] [CrossRef] [PubMed]
- Abram, P.K.; Boivin, G.; Moiroux, J.; Brodeur, J. Behavioural effects of temperature on ectothermic animals: Unifying thermal physiology and behavioural plasticity. Biol. Rev. 2017, 92, 1859–1876. [Google Scholar] [CrossRef] [PubMed]
- Sgro, C.M.; Terblanche, J.S.; Hoffmann, A.A. What can plasticity contribute to insect responses to climate change? Annu. Rev. Entomol. 2016, 61, 433–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the Asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [Green Version]
- Chevin, L.M.; Lande, R.; Mace, G.M. Adaptation, plasticity, and extinction in a changing environment: Towards a predictive theory. PLoS Biol. 2010, 8, e1000357. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, M.A.; Trowbridge, A.M.; Raffa, K.F.; Lindroth, R.L. Consequences of climate warming and altered precipitation patterns for plant-insect and multitrophic interactions. Plant Physiol. 2012, 160, 1719–1727. [Google Scholar] [CrossRef] [Green Version]
- Boullis, A.; Francis, F.; Verheggen, F.J. Climate change and tritrophic interactions: Will modifications to greenhouse gas emissions increase the vulnerability of herbivorous insects to natural enemies? Environ. Entomol. 2015, 44, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Furlong, M.J.; Zalucki, M.P. Climate change and biological control: The consequences of increasing temperatures on host–parasitoid interactions. Curr. Opin. Insect Sci. 2017, 20, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Welch, K.D.; Harwood, J.D. Temporal dynamics of natural enemy–pest interactions in a changing environment. Biol. Control 2014, 75, 18–27. [Google Scholar] [CrossRef]
- Evans, E.W.; Carlile, N.R.; Innes, M.B.; Pitigala, N. Warm springs reduce parasitism of the cereal leaf beetle through phenological mismatch. J. Appl. Entomol. 2013, 137, 383–391. [Google Scholar] [CrossRef]
- Crowder, D.W.; Harwood, J.D. Promoting biological control in a rapidly changing world. Biol. Control 2014, 75, 1–7. [Google Scholar] [CrossRef]
- Gilman, S.E.; Urban, M.C.; Tewksbury, J.; Gilchrist, G.W.; Holt, R.D. A framework for community interactions under climate change. Trends Ecol. Evol. 2010, 25, 325–331. [Google Scholar] [CrossRef]
- Chen, F.; Wu, G.; Parajulee, M.N.; Ge, F. Impact of elevated CO2 on the third trophic level: A predator Harmonia axyridis and a parasitoid Aphidius picipes. Biocontrol Sci. Technol. 2007, 17, 313–324. [Google Scholar] [CrossRef]
- Dyer, L.A.; Richards, L.A.; Short, S.A.; Dodson, C.D. Effects of CO2 and temperature on tritrophic interactions. PLoS ONE 2013, 8, e62528. [Google Scholar] [CrossRef]
- Chen, F.; Ge, F.; Parajulee, M.N. Impact of elevated CO2 on tri-trophic interaction of Gossypium hirsutum, Aphis gossypii, and Leis axyridis. Environ. Entomol. 2005, 34, 37–46. [Google Scholar] [CrossRef]
- Boland, G.J.; Melzer, M.S.; Hopkin, A.; Higgins, V.; Nassuth, A. Climate change and plant diseases in Ontario. Can. J. Plant Pathol. 2004, 26, 335–350. [Google Scholar] [CrossRef]
- Sastry, K.S.; Zitter, T.A. Plant Virus and Viroid Diseases in the Tropics. Volume 2: Epidemiology and Management, 1st ed.; Springer: Dordrecht, The Netherlands, 2014; ISBN 978-94-007-7820-7. [Google Scholar]
- Hull, R. Plant Virology; Elsevier/Academic Press: London, UK, 2014. [Google Scholar]
- Trębicki, P. Climate change and plant virus epidemiology. Virus Res. 2020, 286, 198059. [Google Scholar] [CrossRef]
- Trębicki, P.; Vandegeer, R.K.; Bosque-Pérez, N.A.; Powell, K.S.; Dader, B.; Freeman, A.J.; Yen, A.L.; Fitzgerald, G.J.; Luck, J.E. Virus infection mediates the effects of elevated CO2 on plants and vectors. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canto, T.; Aranda, M.A.; Fereres, A. Climate change effects on physiology and population processes of hosts and vectors that influence the spread of hemipteran-borne plant viruses. Glob. Chang. Biol. 2009, 15, 1884–1894. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.C.; Dhillon, M.K.; Kibuka, J.; Mukuru, S.Z. Plant defense responses to sorghum spotted stem borer, Chilo partellus under irrigated and drought conditions. Int. Sorghum Millets Newsl. 2005, 46, 49–52. [Google Scholar]
- Sharma, H.C. Climate change effects on insects: Implications for crop protection and food security. J. Crop Improv. 2014, 28, 229–259. [Google Scholar] [CrossRef]
- Nault, L.R. Arthropod transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Fereres, A.; Irwin, M.E.; Kampmeier, G.E. Aphid movement: Process and Consequences. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI Publishing: Wallingford, UK, 2017; pp. 196–224. ISBN 9781780647098. [Google Scholar]
- Zeyen, R.J.; Stromberg, E.L.; Kuehnast, E.L. Long-range aphid transport hypothesis for maize dwarf mosaic virus: History and distribution in Minnesota, USA. Ann. Appl. Biol. 1987, 111, 325–336. [Google Scholar] [CrossRef]
- Robert, Y.; Woodford, J.T.; Ducray-Bourdin, D.G. Some epidemiological approaches to the control of aphid-borne virus diseases in seed potato crops in northern Europe. Virus Res. 2000, 71, 33–47. [Google Scholar] [CrossRef]
- Irwin, M.E.; Ruesink, W.G.; Isard, S.A.; Kampmeier, G.E. Mitigating epidemics caused by non-persistently transmitted aphid-borne viruses: The role of the pliant environment. Virus Res. 2000, 71, 185–211. [Google Scholar] [CrossRef]
- Alonso-Prados, J.L.; Luis-Arteaga, M.; Alvarez, J.M.; Moriones, E.; Batlle, A.; Laviña, A.; García-Arenal, F.; Fraile, A. Epidemics of aphid-transmitted viruses in melon crops in Spain. Eur. J. Plant Pathol. 2003, 109, 129–138. [Google Scholar] [CrossRef]
- Jarošová, J.; Żelazny, W.R.; Kundu, J.K. Patterns and predictions of barley yellow dwarf virus vector migrations in Central Europe. Plant Dis. 2019, 103, 2057–2064. [Google Scholar] [CrossRef]
- Roos, J.; Hopkins, R.; Kvarnheden, A.; Dixelius, C. The impact of global warming on plant diseases and insect vectors in Sweden. Eur. J. Plant Pathol. 2011, 129, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Parry, H.R.; Macfadyen, S.; Kriticos, D.J. The geographical distribution of Yellow dwarf viruses and their aphid vectors in Australian grasslands and wheat. Australas. Plant Pathol. 2012, 41, 375–387. [Google Scholar] [CrossRef]
- Sseruwagi, P.; Sserubombwe, W.S.; Legg, J.P.; Ndunguru, J.; Thresh, J.M. Methods of surveying the incidence and severity of cassava mosaic disease and whitefly vector populations on cassava in Africa: A review. Virus Res. 2004, 100, 129–142. [Google Scholar] [CrossRef]
- Ramos, R.S.; Kumar, L.; Shabani, F.; da Silva, R.S.; de Araújo, T.A.; Picanço, M.C. Climate model for seasonal variation in Bemisia tabaci using CLIMEX in tomato crops. Int. J. Biometeorol. 2019, 63, 281–291. [Google Scholar] [CrossRef]
- Boudon-Padieu, E. Phytoplasmas associated to Grapevine yellows and potential vectors. Bull. O.I.V. 2005, 78, 299–320. [Google Scholar]
- Boudon-Padieu, E. The situation of grapevine yellows and current research directions: Distribution, diversity, vectors, diffusion and control. In Proceedings of the Extended Abstracts, 14th Meeting of the ICVG, Locorotondo, Italy, 12–17 September 2003; pp. 47–53. [Google Scholar]
- Falzoi, S.; Lessio, F.; Spanna, F.; Alma, A. Influence of temperature on the embryonic and post-embryonic development of Scaphoideus titanus (Hemiptera: Cicadellidae), vector of grapevine Flavescence dorée. Int. J. Pest. Manag. 2014, 60, 246–257. [Google Scholar] [CrossRef]
- Boudon-Padieu, É.; Maixner, M. Potential effects of climate change on distribution and activity of insect vectors of grapevine pathogens. In Proceedings of the International and Multi-Disciplinary “Global Warming, Which Potential Impacts on the Vineyards?”, Beaune, France, 28–30 March 2007; p. 23. [Google Scholar]
- Boudon-Padieu, E. Cicadelle vectrice de la flavescence dorée, Scaphoideus titanus Ball, 1932. In Ravageurs de la Vigne; Stockel, J., Ed.; Féret: Bordeaux, France, 2000; pp. 110–120. [Google Scholar]
- Mirutenko, V.; Janský, V.; Margitay, V. First records of Scaphoideus titanus (Hemiptera, Cicadellidae) in Ukraine. EPPO Bull. 2018, 48, 167–168. [Google Scholar] [CrossRef]
- Howden, S.M.; Soussana, J.F.; Tubiello, F.N.; Chhetri, N.; Dunlop, M.; Meinke, H. Adapting agriculture to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19691–19696. [Google Scholar] [CrossRef] [Green Version]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Boller, E.F.; Avilla, J.; Joerg, E.; Malavolta, C.; Wijnands, F.G.; Esbjerg, P. Integrated Production: Principles and Technical Guidelines; IOBC/WPRS: Dijon, France, 2004; Available online: https://www.iobc-wprs.org/ip_ipm/01_IOBC_Principles_and_Tech_Guidelines_2004.pdf (accessed on 13 March 2015).
- Gomez-Zavaglia, A.; Mejuto, J.C.; Simal-Gandara, J. Mitigation of emerging implications of climate change on food production systems. Food Res. Int. 2020, 134, 109256. [Google Scholar] [CrossRef]
- FAO. How to Practice Integrated Pest Management. Available online: http://www.fao.org/agriculture/crops/thematic-sitemap/theme/compendium/tools-guidelines/how-to-ipm/en/ (accessed on 15 February 2021).
- Lamichhane, J.R.; Barzman, M.; Booij, K.; Boonekamp, P.; Desneux, N.; Huber, L.; Kudsk, P.; Langrell, S.R.H.; Ratnadass, A.; Ricci, P.; et al. Robust cropping systems to tackle pests under climate change. A review. Agron. Sustain. Dev. 2015, 35, 443–459. [Google Scholar] [CrossRef]
- Trumble, J.; Butler, C. Climate change will exacerbate California’s insect pest problems. Calif. Agric. 2009, 63, 73–78. [Google Scholar] [CrossRef]
- Andrew, N.R.; Hill, S.J. Effect of climate change on insect pest management. In Environmental Pest Management: Challenges for Agronomists, Ecologists, Economists and Policymakers, 1st ed.; Coll, M., Wajnberg, E., Eds.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 197–215. ISBN 978-1-119-25555-0. [Google Scholar]
- Heuskin, S.; Verheggen, F.J.; Haubruge, E.; Wathelet, J.P.; Lognay, G. The use of semiochemical slow-release devices in integrated pest management strategies. Biotechnol. Agron. Soc. Environ. 2011, 15, 459–470. [Google Scholar]
- Nihal, R. Global Climate change and its impact on integrated pest management. Agro Econ. Int. J. 2020, 7, 133–137. [Google Scholar]
- Wenda-Piesik, A.; Piesik, D.; Nowak, A.; Wawrzyniak, M. Tribolium confusum responses to blends of cereal kernels and plant volatiles. J. Appl. Entomol. 2015, 140, 558–563. [Google Scholar] [CrossRef]
- Vadez, V.; Berger, J.D.; Warkentin, T.; Asseng, S.; Ratnakumar, P.; Poorna Chandra Rao, K.; Gaur, P.M.; Munier-Jolain, N.; Larmure, A.; Voisin, A.-S.; et al. Adaptation of grain legumes to climate change: A review. Agron. Sustain. Dev. 2012, 32, 31–44. [Google Scholar] [CrossRef]
- Yamamura, K.; Yokozawa, M.; Nishimori, M.; Ueda, Y.; Yokosuka, T. How to analyze long-term insect population dynamics under climate change: 50-year data of three insect pests in paddy fields. Popul. Ecol. 2006, 48, 31–48. [Google Scholar] [CrossRef]
- Heeb, L.; Jenner, E.; Cock, M.J. Climate-smart pest management: Building resilience of farms and landscapes to changing pest threats. J. Pest. Sci. 2019, 92, 951–969. [Google Scholar] [CrossRef]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef]
- Perrings, C.; Burgiel, S.; Lonsdale, M.; Mooney, H.; Williamson, M. International cooperation in the solution to trade-related invasive species risks. Ann. N. Y. Acad. Sci. 2010, 1195, 198–212. [Google Scholar] [CrossRef]
- Joyce, L.A.; Briske, D.D.; Brown, J.R.; Polley, H.W.; McCarl, B.A.; Bailey, D.W. Climate change and North American rangelands: Assessment of mitigation and adaptation strategies. Rangel. Ecol. Manag. 2013, 66, 512–528. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Neven, L.G.; Yee, W.L. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Evans, T.G.; Diamond, S.E.; Kelly, M.W.X. Mechanistic species distribution modelling as a link between physiology and conservation. Conserv. Physiol. 2015, 3, cov056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillson, L.; Dawson, T.P.; Jack, S.; McGeoch, M.A. Accommodating climate change contingencies in conservation strategy. Trends Ecol. Evol. 2013, 28, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Porfirio, L.L.; Harris, R.M.; Lefroy, E.C.; Hugh, S.; Gould, S.F.; Lee, G.; Bindoff, N.B.; Mackey, B. Improving the use of species distribution models in conservation planning and management under climate change. PLoS ONE 2014, 9, e113749. [Google Scholar] [CrossRef] [Green Version]
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.M.; Butchart, S.H.M.; Kovacs, K.M.; Scheffers, B.R.; Hole, D.G.; Martin, T.G.; Akcakaya, H.R.; et al. Assessing species vulnerability to climate change. Nat. Clim. Chang. 2015, 5, 215–224. [Google Scholar] [CrossRef]
- Kearney, M.R.; Wintle, B.A.; Porter, W.P. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conserv. Lett. 2010, 3, 203–213. [Google Scholar] [CrossRef]
- Cuddington, K.; Fortin, M.J.; Gerber, L.R.; Hastings, A.; Liebhold, A.; O’Connor, M.; Ray, C. Process-based models are required to manage ecological systems in a changing world. Ecosphere 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Maywald, G.F.; Yonow, T.; Zurcher, E.J.; Herrmann, N.I.; Sutherst, R. Exploring the effects of climate on plants, animals and diseases. CLIMEX Version 2015, 4, 184. [Google Scholar]
- Yonow, T.; Kriticos, D.J.; Kirichenko, N.; Ota, N. Considering biology when inferring range-limiting stress mechanisms for agricultural pests: A case study of the beet armyworm. J. Pest. Sci. 2018, 91, 523–538. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. https://doi.org/10.3390/insects12050440
Skendžić S, Zovko M, Živković IP, Lešić V, Lemić D. The Impact of Climate Change on Agricultural Insect Pests. Insects. 2021; 12(5):440. https://doi.org/10.3390/insects12050440
Chicago/Turabian StyleSkendžić, Sandra, Monika Zovko, Ivana Pajač Živković, Vinko Lešić, and Darija Lemić. 2021. "The Impact of Climate Change on Agricultural Insect Pests" Insects 12, no. 5: 440. https://doi.org/10.3390/insects12050440
APA StyleSkendžić, S., Zovko, M., Živković, I. P., Lešić, V., & Lemić, D. (2021). The Impact of Climate Change on Agricultural Insect Pests. Insects, 12(5), 440. https://doi.org/10.3390/insects12050440