
A Multi-Scale Model of Disease Transfer in Honey Bee Colonies


Abstract
:Simple Summary
Abstract
1. Introduction
2. Model
3. Results
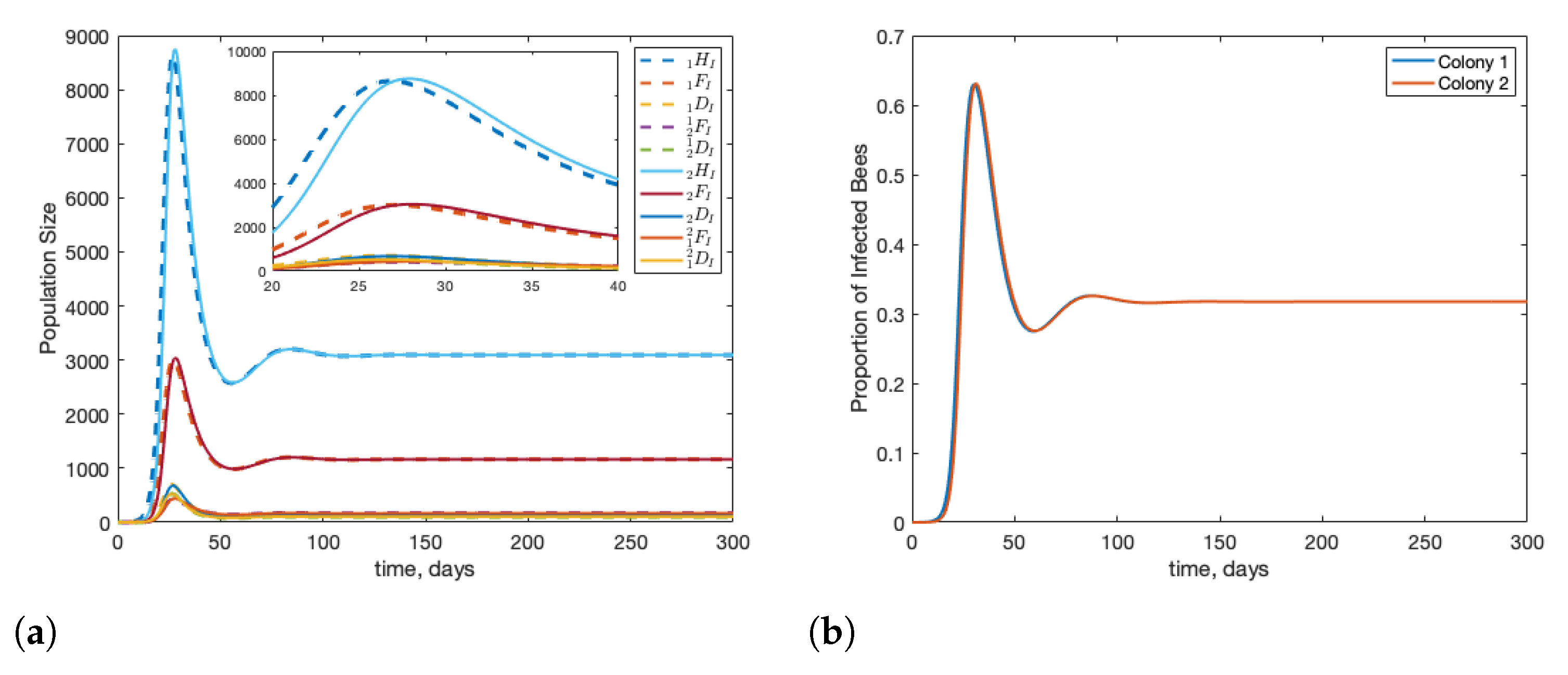
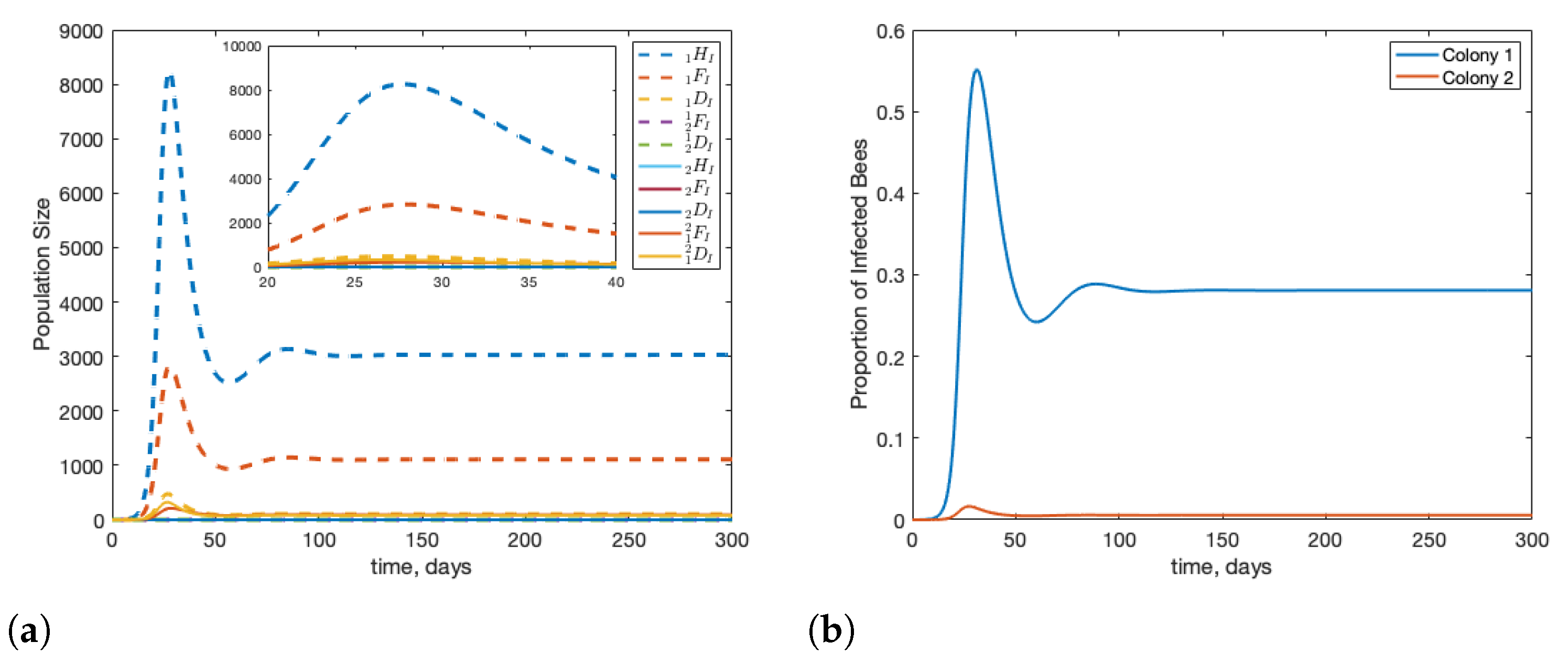
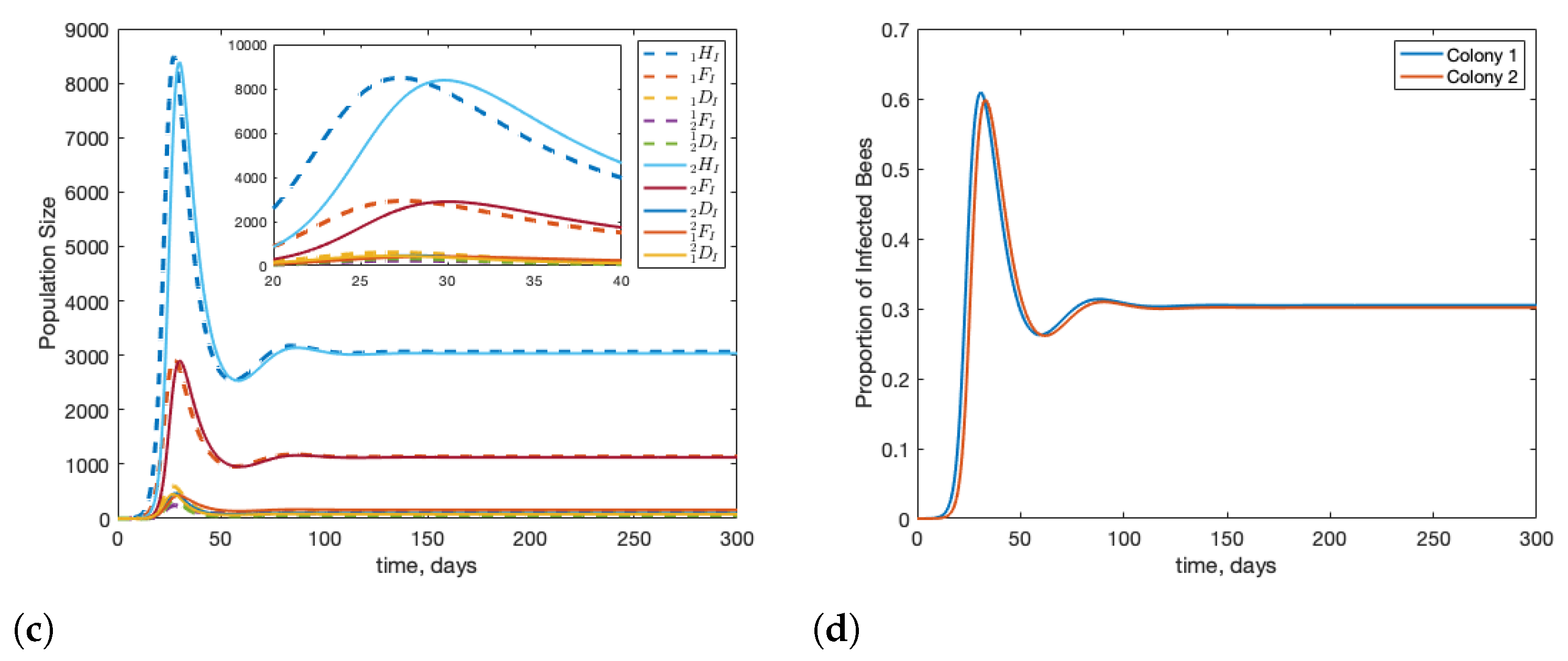
3.1. Scenario: Drone Drift vs. Forager Drift
3.2. Scenario: Filtering Infected Bees
3.3. Scenario: Robbing Behaviour
3.4. Basic Reproduction Number
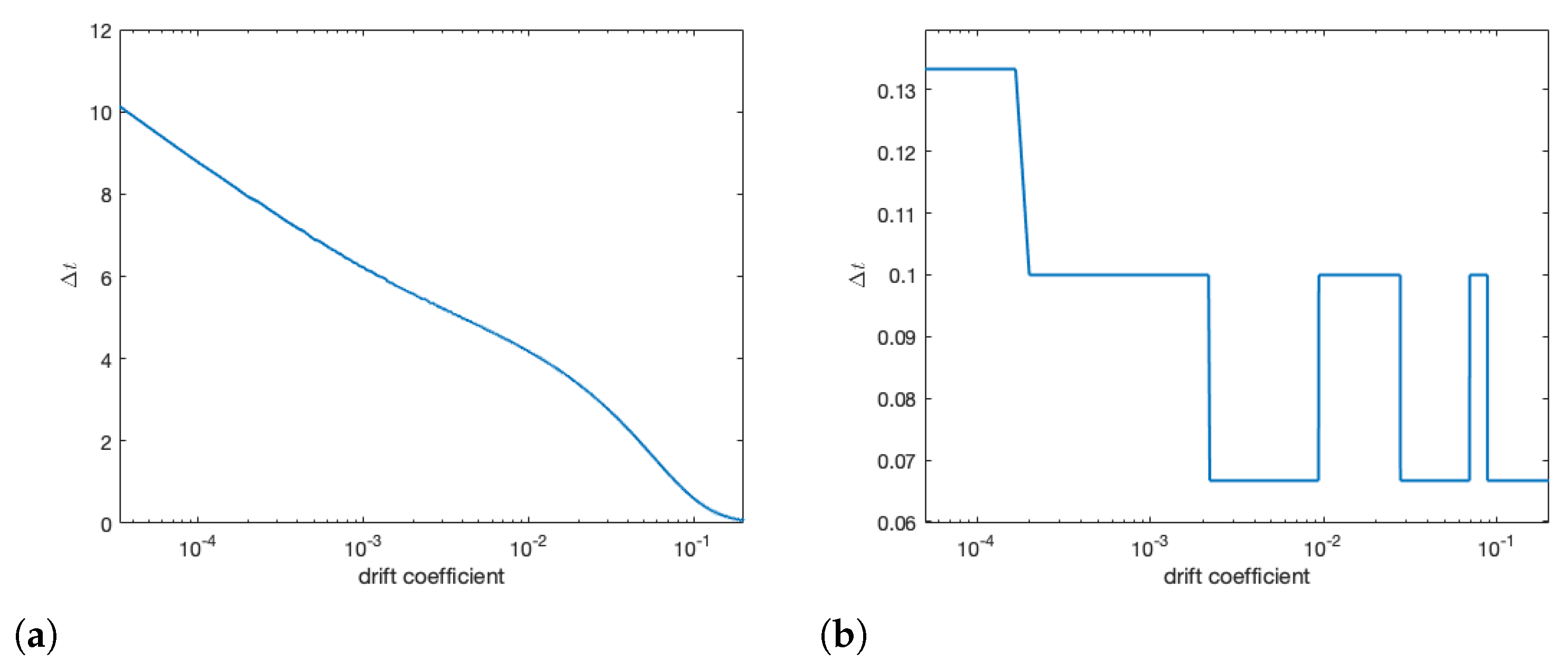
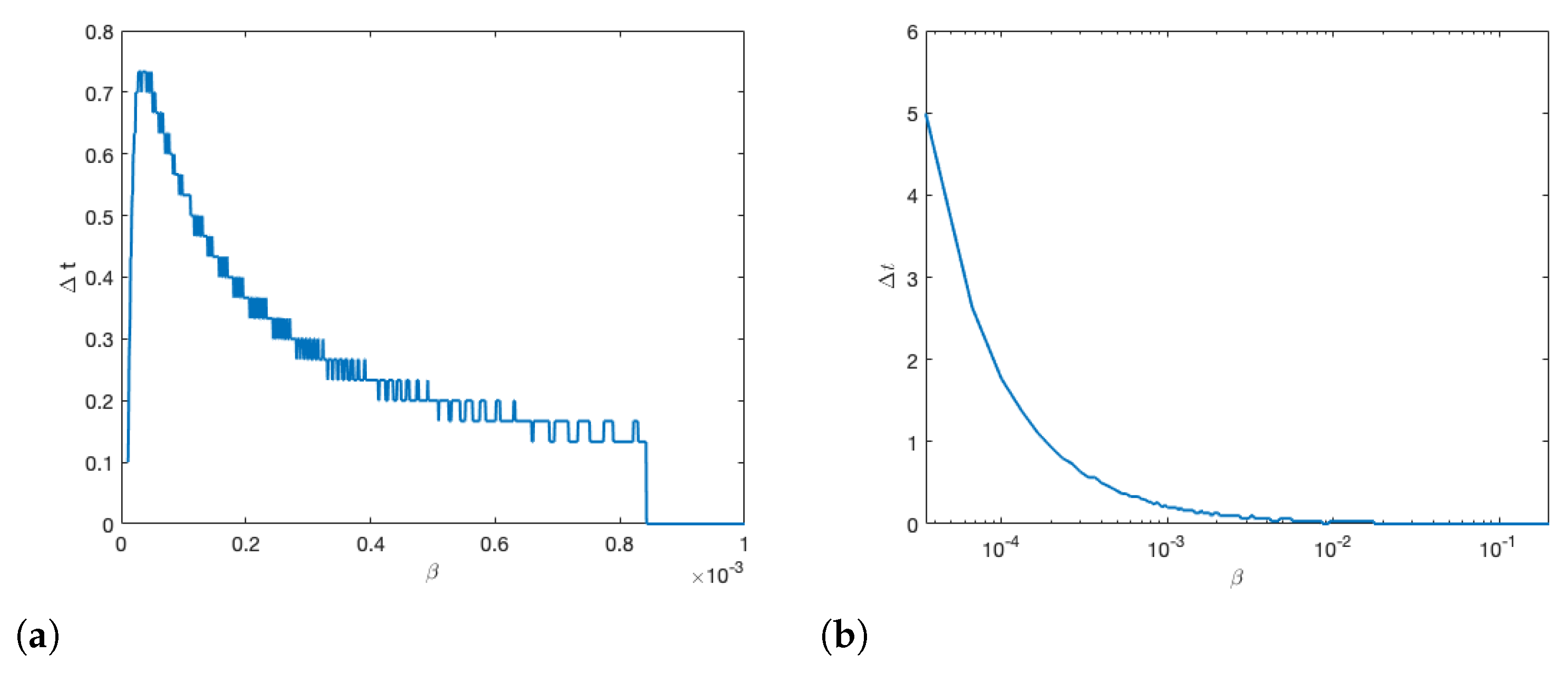
3.5. Varying Drift & Reproduction Number
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Owen, R. Role of human action in the spread of honey bee (Hymenoptera: Apidae) pathogens. J. Econ. Entomol. 2017, 110, 797–801. [Google Scholar] [CrossRef]
- Coulon, M.; Dalmon, A.; Di Prisco, G.; Prado, A.; Arban, F.; Dubois, E.; Ribière-Chabert, M.; Alaux, C.; Thiéry, R.; Le Conte, Y. Interactions between thiamethoxam and Deformed wing virus can drastically impair flight behavior of honey bees. Front. Microbiol. 2020, 11, 766. [Google Scholar] [CrossRef]
- Flores, J.M.; Gil-Lebrero, S.; Gámiz, V.; Rodríguez, M.I.; Ortiz, M.A.; Quiles, F.J. Effect of the climate change on honey bee colonies in a temperate Mediterranean zone assessed through remote hive weight monitoring system in conjunction with exhaustive colonies assessment. Sci. Total Environ. 2019, 653, 1111–1119. [Google Scholar] [CrossRef]
- Colin, T.; Meikle, W.G.; Paten, A.M.; Barron, A.B. Long-term dynamics of honey bee colonies following exposure to chemical stress. Sci. Total Environ. 2019, 677, 660–670. [Google Scholar] [CrossRef]
- De Jong, D. Mites: Varroa and other parasites of brood. In Honey Bee Pests, Predators and Diseases; Cornell University Press: Ithaca, NY, USA, 1997; Volume 1, pp. 279–327. [Google Scholar]
- Wilfert, L.; Long, G.; Leggett, H.; Schmid-Hempel, P.; Butlin, R.; Martin, S.; Boots, M. Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [Green Version]
- Di Prisco, G.; Pennacchio, F.; Caprio, E.; Boncristiani, H.F., Jr.; Evans, J.D.; Chen, Y. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera. J. Gen. Virol. 2011, 92, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Carreck, N.L.; Ball, B.V.; Martin, S.J. The epidemiology of cloudy wing virus infections in honey bee colonies in the UK. J. Apic. Res. 2010, 49, 66–71. [Google Scholar] [CrossRef]
- Ravoet, J.; De Smet, L.; Wenseleers, T.; De Graaf, D.C. Genome sequence heterogeneity of Lake Sinai Virus found in honey bees and Orf1/RdRP-based polymorphisms in a single host. Virus Res. 2015, 201, 67–72. [Google Scholar] [CrossRef]
- Spivak, M.; Gilliam, M. Hygienic behaviour of honey bees and its application for control of brood diseases and Varroa: Part I. Hygienic behaviour and resistance to American foulbrood. Bee World 1998, 79, 124–134. [Google Scholar] [CrossRef]
- Spivak, M. Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie 1996, 27, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Fefferman, N.H.; Traniello, J.F.; Rosengaus, R.B.; Calleri, D.V., II. Disease prevention and resistance in social insects: Modeling the survival consequences of immunity, hygienic behavior, and colony organization. Behav. Ecol. Sociobiol. 2007, 61, 565–577. [Google Scholar] [CrossRef]
- Moore, A.J.; Breed, M.D.; Moor, M.J. The guard honey bee: Ontogeny and behavioural variability of workers performing a specialized task. Anim. Behav. 1987, 35, 1159–1167. [Google Scholar] [CrossRef]
- Free, J. The drifting of honey-bees. J. Agric. Sci. 1958, 51, 294–306. [Google Scholar] [CrossRef]
- Goodwin, R.; Taylor, M.; Mcbrydie, H.; Cox, H. Drift of Varroa destructor-infested worker honey bees to neighbouring colonies. J. Apic. Res. Bee World 2006, 45, 155. [Google Scholar] [CrossRef]
- Dynes, T.L.; Berry, J.A.; Delaplane, K.S.; Brosi, B.J.; de Roode, J.C. Reduced density and visually complex apiaries reduce parasite load and promote honey production and overwintering survival in honey bees. PLoS ONE 2019, 14, e0216286. [Google Scholar]
- Rittschof, C.C.; Nieh, J.C. Honey robbing: Could human alterations to the environment change a rare foraging tactic into a maladaptive behavior? Curr. Opin. Insect Sci. 2021, 45, 84–90. [Google Scholar] [CrossRef]
- Peck, D.T.; Seeley, T.D. Mite bombs or robber lures? The roles of drifting and robbing in Varroa destructor transmission from collapsing honey bee colonies to their neighbors. PLoS ONE 2019, 14, e0218392. [Google Scholar] [CrossRef]
- Graystock, P.; Goulson, D.; Hughes, W.O. Parasites in bloom: Flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. R. Soc. B Biol. Sci. 2015, 282, 20151371. [Google Scholar] [CrossRef] [Green Version]
- Purkiss, T.; Lach, L. Pathogen spillover from Apis mellifera to a stingless bee. Proc. R. Soc. B 2019, 286, 20191071. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Brown, M.J.; Stevenson, P.C. The role of disease in bee foraging ecology. Curr. Opin. Insect Sci. 2017, 21, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Betti, M.I.; Wahl, L.M.; Zamir, M. Effects of Infection on Honey Bee Population Dynamics: A Model. PLoS ONE 2014, 9, e110237. [Google Scholar] [CrossRef] [PubMed]
- Betti, M.I.; Wahl, L.M.; Zamir, M. Reproduction Number and Asymptotic Stability for a Model with Continuous Age Structure: An Application to Honey Bee Dynamics. Bull. Math. Biol. 2016. submitted. [Google Scholar]
- Betti, M.; Wahl, L.; Zamir, M. Age structure is critical to the population dynamics and survival of honeybee colonies. R. Soc. Open Sci. 2016, 3, 160444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, N.; Eberl, H.J. Two routes of transmission for Nosema infections in a honeybee population model with polyethism and time-periodic parameters can lead to drastically different qualitative model behavior. Commun. Nonlinear Sci. Numer. Simul. 2020, 84, 105207. [Google Scholar] [CrossRef]
- Ratti, V.; Kevan, P.G.; Eberl, H.J. A mathematical model for population dynamics in honeybee colonies infested with Varroa destructor and the Acute Bee Paralysis Virus. Can. Appl. Math. Q. 2013, 21, 63–93. [Google Scholar]
- Eberl, H.J.; Frederick, M.R.; Kevan, P.G. Importance of brood maintenance terms in simple models of the honeybee—Varroa destructor—Acute Bee Paralysis Virus complex. Electron. J. Differ. Equ. 2010, 2010, 85–98. [Google Scholar]
- Petric, A.; Guzman-Novoa, E.; Eberl, H.J. A mathematical model for the interplay of Nosema infection and forager losses in honey bee colonies. J. Biol. Dyn. 2017, 11, 348–378. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, N.; Eberl, H.J. A simple model of between-hive transmission of Nosemosis. In International Conference on Applied Mathematics, Modeling and Computational Science; Springer: Berlin/Heidelberg, Germany, 2017; pp. 385–395. [Google Scholar]
- Bartlett, L.J.; Rozins, C.; Brosi, B.J.; Delaplane, K.S.; de Roode, J.C.; White, A.; Wilfert, L.; Boots, M. Industrial bees: The impact of apicultural intensification on local disease prevalence. J. Appl. Ecol. 2019, 56, 2195–2205. [Google Scholar] [CrossRef] [Green Version]
- Le Conte, Y.; Ellis, M.; Ritter, W. Varroa mites and honey bee health: Can Varroa explain part of the colony losses? Apidologie 2010, 41, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Betti, M.; LeClair, J.; Wahl, L.M.; Zamir, M. Bee++: An Object-Oriented, Agent-Based Simulator for Honey Bee Colonies. Insects 2017, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Fries, I. Nosema apis—A parasite in the honey bee colony. Bee World 1993, 74, 5–19. [Google Scholar] [CrossRef]
- Khoury, D.S.; Myerscough, M.R.; Barron, A.B. A Quantitative Model of Honey Bee Colony Population Dynamics. PLoS ONE 2011, 6, e18491. [Google Scholar] [CrossRef] [Green Version]
- Khoury, D.S.; Barron, A.B.; Myerscough, M.R. Modelling Food and Population Dynamics in Honey Bee Colonies. PLoS ONE 2013, 8, e59084. [Google Scholar] [CrossRef] [Green Version]
- Heidinger, I.M.M.; Meixner, M.D.; Berg, S.; Büchler, R. Observation of the mating behavior of honey bee (Apis mellifera L.) queens using radio-frequency identification (RFID): Factors influencing the duration and frequency of nuptial flights. Insects 2014, 5, 513–527. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.; Mikheyev, A. Reproductive decisions by honey bee colonies: Tuning investment in male production in relation to success in energy acquisition. Insectes Sociaux 2003, 50, 134–138. [Google Scholar] [CrossRef]
- Fukuda, H.; Ohtani, T. Survival and life span of drone honeybees. Popul. Ecol. 1977, 19, 51–68. [Google Scholar] [CrossRef]
- Russell, S.; Barron, A.B.; Harris, D. Dynamic modelling of honey bee (Apis mellifera) colony growth and failure. Ecol. Model. 2013, 265, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Hrassnigg, N.; Crailsheim, K. Differences in drone and worker physiology in honeybees (Apis mellifera). Apidologie 2005, 36, 255–277. [Google Scholar] [CrossRef] [Green Version]
- Jay, S. Seasonal development of honeybee colonies started from package bees. J. Apic. Res. 1974, 13, 149–152. [Google Scholar] [CrossRef]
- Neumann, P.; Moritz, R.F.; Mautz, D. Colony evaluation is not affected by drifting of drone and worker honeybees (Apis mellifera L.) at a performance testing apiary. Apidologie 2000, 31, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Free, J.; Racey, P. The effect of the size of honeybee colonies on food consumption, brood rearing and the longevity of the bees during winter. Entomol. Exp. Appl. 1968, 11, 241–249. [Google Scholar] [CrossRef]
- Downs, S.G.; Ratnieks, F.L. Adaptive shifts in honey bee (Apis mellifera L.) guarding behavior support predictions of the acceptance threshold model. Behav. Ecol. 2000, 11, 326–333. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, K.; Crailsheim, K. Drifting of honeybees. Insectes Sociaux 1998, 45, 151–167. [Google Scholar] [CrossRef]
- Wolf, S.; Nicholls, E.; Reynolds, A.M.; Wells, P.; Lim, K.S.; Paxton, R.J.; Osborne, J.L. Optimal search patterns in honeybee orientation flights are robust against emerging infectious diseases. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Milbrath, M.O.; van Tran, T.; Huang, W.F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Mattila, H.; Otis, G. Effects of pollen availability and Nosema infection during the spring on division of labor and survival of worker honey bees (Hymenoptera: Apidae). Environ. Entomol. 2006, 35, 708–717. [Google Scholar] [CrossRef]
- Botías, C.; Martín-Hernández, R.; Días, J.; García-Palencia, P.; Matabuena, M.; Juarranz, Á.; Barrios, L.; Meana, A.; Nanetti, A.; Higes, M. The effect of induced queen replacement on Nosema spp. infection in honey bee (Apis mellifera iberiensis) colonies. Environ. Microbiol. 2012, 14, 845–859. [Google Scholar]
- Liess, M.; Foit, K.; Knillmann, S.; Schäfer, R.B.; Liess, H.D. Predicting the synergy of multiple stress effects. Sci. Rep. 2016, 6, 1–8. [Google Scholar]
- Comper, J.R.; Eberl, H.J. Mathematical modelling of population and food storage dynamics in a honey bee colony infected with Nosema ceranae. Heliyon 2020, 6, e04599. [Google Scholar] [CrossRef] [PubMed]
- Geffre, A.C.; Gernat, T.; Harwood, G.P.; Jones, B.M.; Gysi, D.M.; Hamilton, A.R.; Bonning, B.C.; Toth, A.L.; Robinson, G.E.; Dolezal, A.G. Honey bee virus causes context-dependent changes in host social behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 10406–10413. [Google Scholar] [CrossRef]
- Bromenshenk, J.J.; DeGrandi-Hoffman, G. PC BEEPOP: A Microcomputer Model of the Population Dynamics and Economics of Honey Bee Colonies. Am. Entomol. 1993, 39, 231–237. [Google Scholar] [CrossRef]
- Fürst, M.; McMahon, D.P.; Osborne, J.; Paxton, R.; Brown, M. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Manley, R.; Boots, M.; Wilfert, L. Emerging viral disease risk to pollinating insects: Ecological, evolutionary and anthropogenic factors. J. Appl. Ecol. 2015, 52, 331–340. [Google Scholar] [CrossRef]
- Dalmon, A.; Diévart, V.; Thomasson, M.; Fouque, R.; Vaissière, B.E.; Guilbaud, L.; Le Conte, Y.; Henry, M. Possible spillover of pathogens between bee communities foraging on the same floral resource. Insects 2021, 12, 122. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Value | Ref. |
|---|---|---|---|
| L | maximum rate of egg laying | 2000 eggs/day | [35] |
| w | number of hive bees for 50% egg survival | 5000 bees | [35] |
| b | mass of food stored for 50% egg survival | 500 g | [36] |
| k | increased food requirements for drone brood survival | variable | |
| maximum rate of recruitment | /day | [35] | |
| maximum fraction of colony that can be foraging | [35] | ||
| natural death rate of foragers (summer) | [35] | ||
| natural death rate of drones | [39] | ||
| c | food gathered per day per forager | 0.1 g/day/bee | [40] |
| daily food requirement per adult hive bee | 0.008 g/day/bee | [41] | |
| daily food requirement per adult forager | 0.008 g/day/bee | [41] | |
| daily food requirement per adult drone | 0.016 g/day/bee | calculated | |
| daily food requirement per worker brood | 0.016 g/day/bee | [42] | |
| daily food requirement per drone brood | 0.049 g/day/bee | [42] | |
| p | proportion of worker eggs | [43] | |
| rate of transmission of infection | [23] | ||
| drift rate of foragers | [44] | ||
| drift rate of drones | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Betti, M.; Shaw, K. A Multi-Scale Model of Disease Transfer in Honey Bee Colonies. Insects 2021, 12, 700. https://doi.org/10.3390/insects12080700
Betti M, Shaw K. A Multi-Scale Model of Disease Transfer in Honey Bee Colonies. Insects. 2021; 12(8):700. https://doi.org/10.3390/insects12080700
Chicago/Turabian StyleBetti, Matthew, and Karalyne Shaw. 2021. "A Multi-Scale Model of Disease Transfer in Honey Bee Colonies" Insects 12, no. 8: 700. https://doi.org/10.3390/insects12080700
APA StyleBetti, M., & Shaw, K. (2021). A Multi-Scale Model of Disease Transfer in Honey Bee Colonies. Insects, 12(8), 700. https://doi.org/10.3390/insects12080700