
Enhancement of the Diversity of Pollinators and Beneficial Insects in Intensively Managed Vineyards

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Areas of Study
2.2. Mixture Plant Selection
2.3. Experimental Design and Sampling
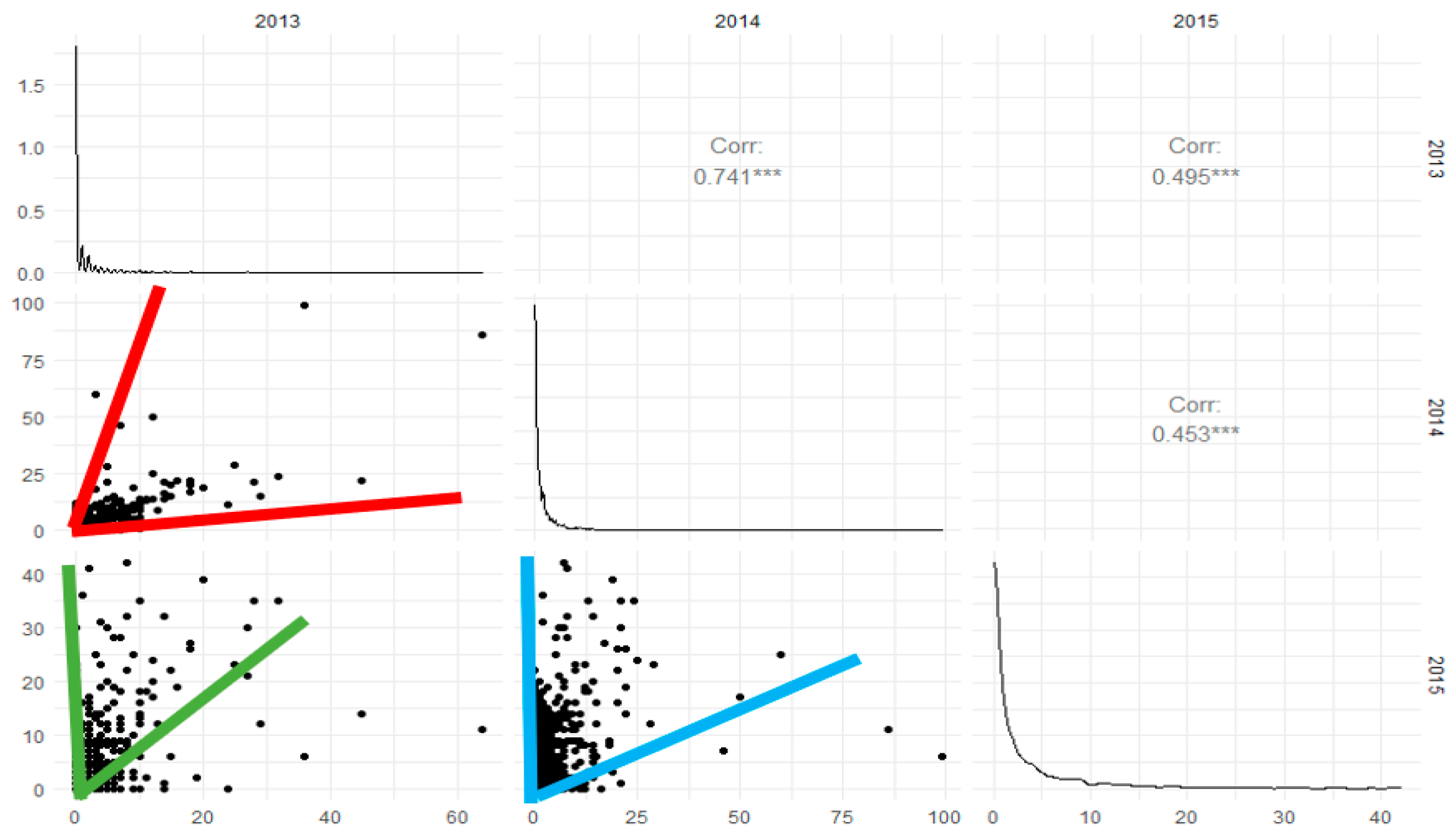
2.4. Data Analysis
3. Results
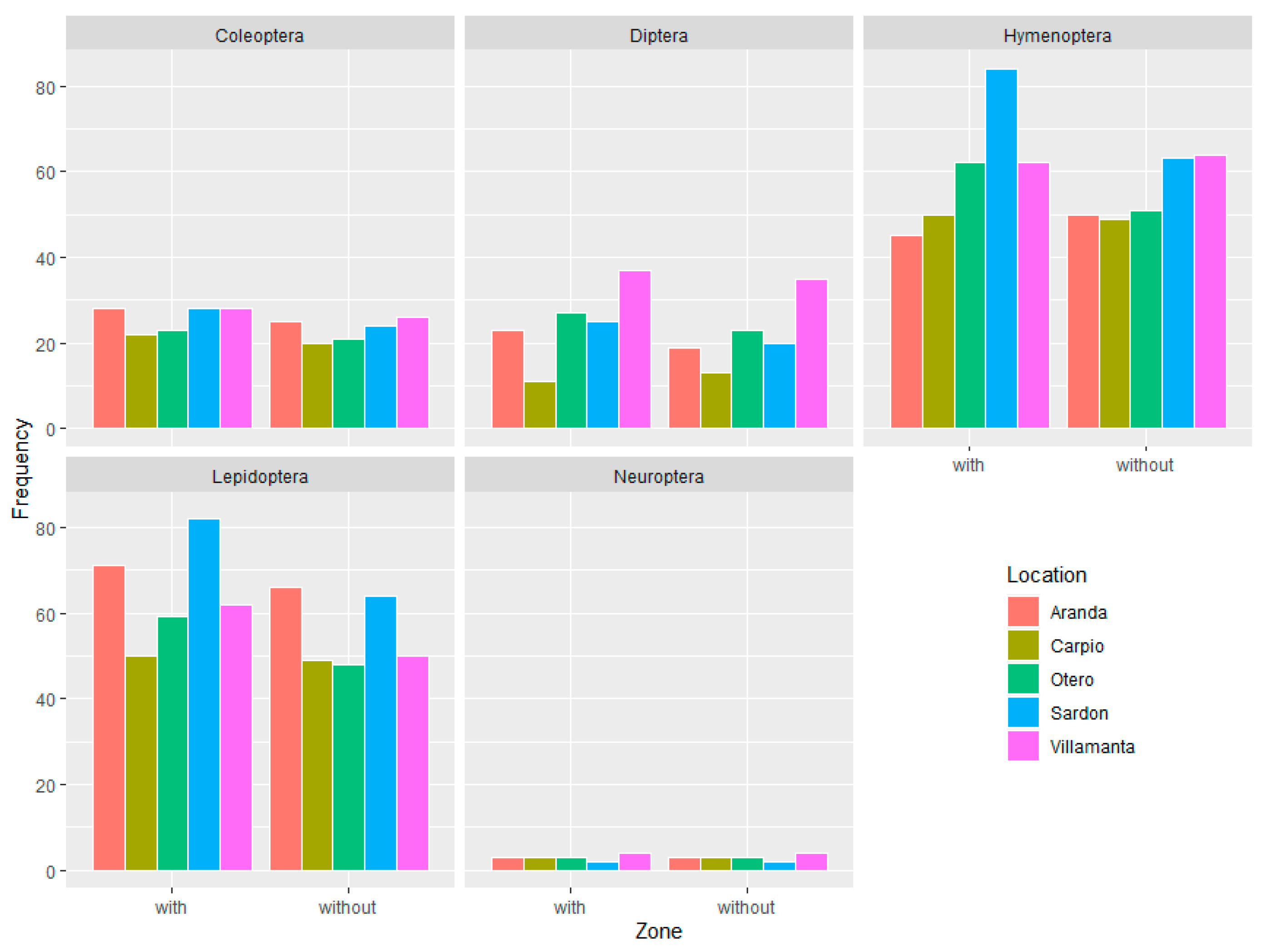
3.1. Diversity of Insects
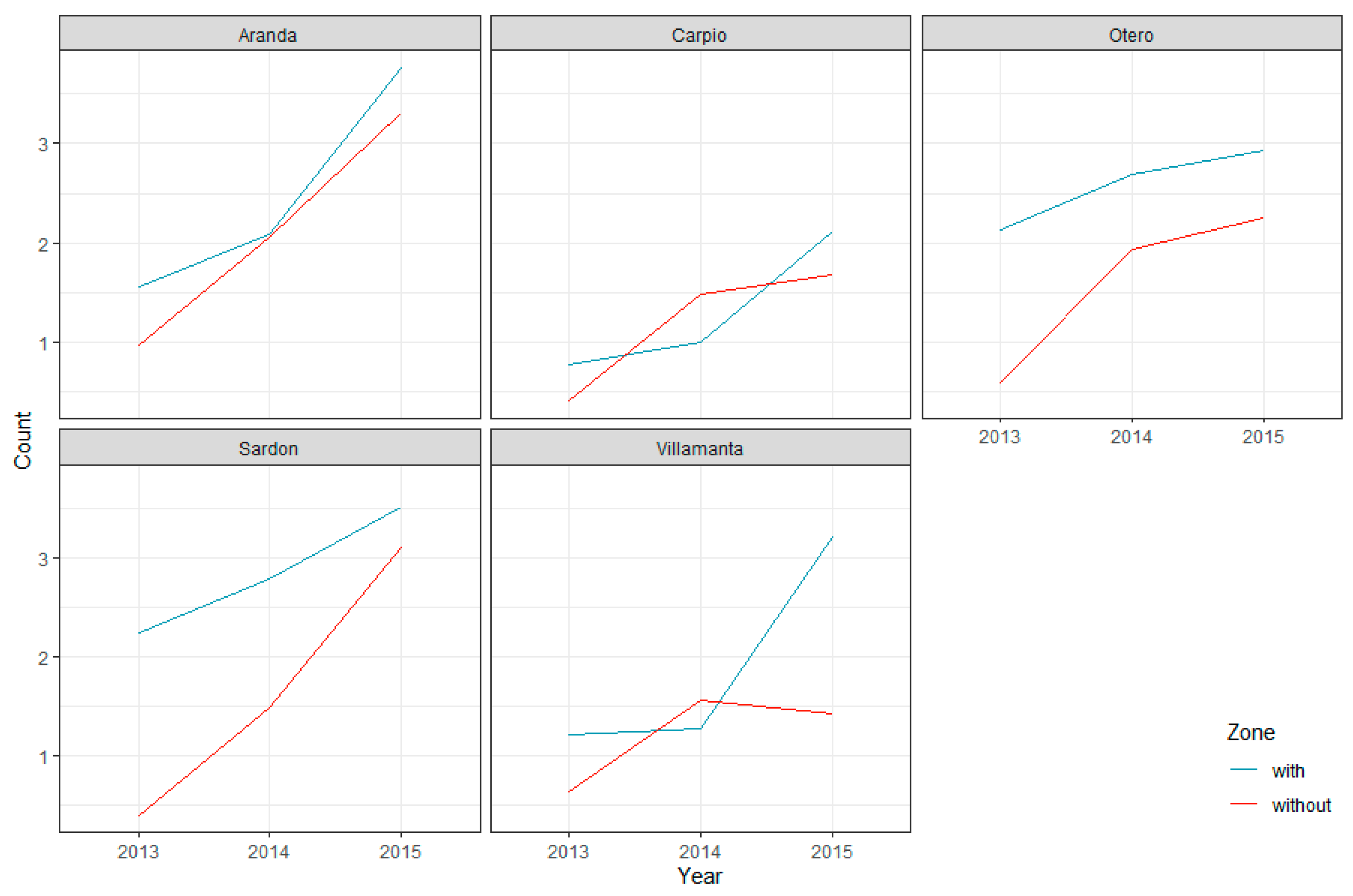
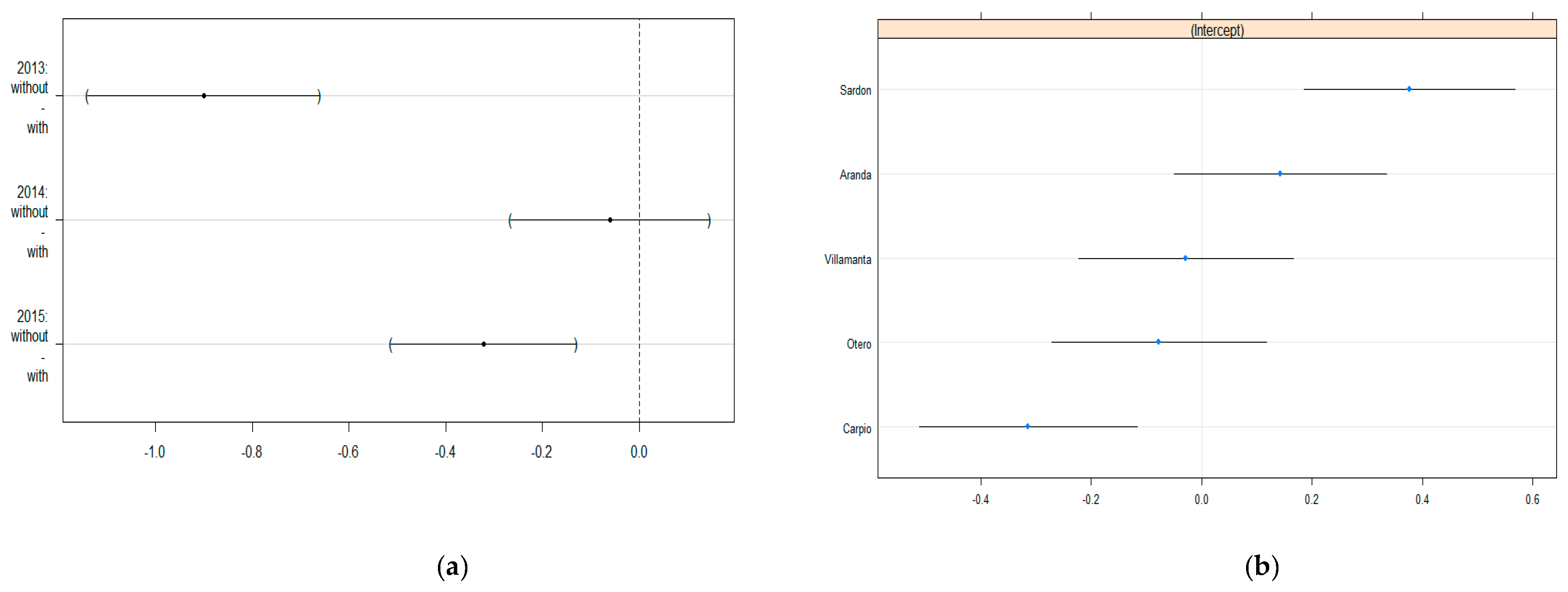
3.2. GLMM Modelling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stoate, C.; Boatman, N.; Borralho, R.; Carvalho, C.R.; De Snoo, G.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Robinson, R.A.; Sutherland, W.J. Post-war changes in arable farming and biodiversity in Great Britain. J. Appl. Ecol. 2020, 39, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.; Potts, S.G.; Woodcock, B.A.; Eggleton, P. Can arable field margins be managed to enhance their biodiversity, conservation and functional value for soil macrofauna? J. Appl. Ecol. 2008, 45, 269–278. [Google Scholar] [CrossRef]
- Wesche, K.; Krause, B.; Culmsee, H.; Leuschner, C. Fifty years of change in Central European grassland vegetation: Large losses in species richness and animal-pollinated plants. Biol. Conserv. 2012, 150, 76–85. [Google Scholar] [CrossRef]
- Nowakowski, M.; Pywell, R. Habitat Creation and Management for Pollinators; Centre for Ecology & Hydrology: Wallingford, UK, 2016. [Google Scholar]
- Klein, A.; Vaissière, B.E.; Cane, J.H.; Stean-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Memmott, J.; Craze, P.G.; Waser, N.M.; Price, M.V. Global warming and the disruption of plant–pollinator interactions. Ecol. Lett. 2007, 10, 710–717. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Morrison, J.; Izquierdo, J.; Plaza, E.H.; González-Andújar, J.L.; Hernández, E.; González-Andújar, J.L. The role of field margins in supporting wild bees in Mediterranean cereal agroecosystems: Which biotic and abiotic factors are important? Agric. Ecosyst. Environ. 2017, 247, 216–224. [Google Scholar] [CrossRef]
- Sánchez, J.A.; Carrasco, A.; La Spina, M.; Pérez-Marcos, M.; Ortíz-Sanchez, F.J. How bees respond differently to field margins of shrubby and herbaceous plants in intensive agricultural crops of the Mediterranean area. Insects 2020, 11, 26. [Google Scholar] [CrossRef] [Green Version]
- New, T.R. Invertebrate Conservation in Agricultural Ecosystems; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Haddaway, N.R.; Brown, C.; Eggers, S.; Josefsson, J.; Kronvang, B.; Randall, N.; Uusi-Kämppä, J. The multifunctional roles of vegetated strips around and within agricultural fields. A systematic map protocol. Environ. Evid. 2016, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Castle, D.; Grass, I.; Westphal, C. Fruit quantity of strawberries benefit from enhanced pollinator abundance at hedgerows in agricultural landscapes. Agric. Ecosyst. Environ. 2019, 275, 14–22. [Google Scholar] [CrossRef]
- Holden, J.; Grayson, R.P.; Berdeni, D.; Bird, S.; Chpaman, P.J.; Edmondson, J.L.; Firbank, L.G.; Helgason, T.; Hodons, M.E.; Hunt, S.F.P.; et al. The role of hedgerows in soil functioning within agricultural landscapes. Agric. Ecosyst. Environ. 2019, 273, 1–12. [Google Scholar] [CrossRef]
- Richards, A.J. Does low biodiversity resulting from modern agricultural practice affect crop pollination and yield? Ann. Bot. 2001, 88, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D. Parallel declines in pollinators and insect-pollinated plants in britain and the netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Stean-Dewenter, I.; Klein, A.M.; Alfert, T.; Gaebele, V.; Tscharntke, T. Bee Diversity and Plant–Pollinator Interactions in Fragmented Landscapes. Specialization and Generalization in PLANT–Pollinator Interactions; Waser, M., Ollerton, N.J., Eds.; Chicago Press: Chicago, IL, USA, 2006; pp. 387–408. [Google Scholar]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape effects on crop pollination services: Are there general patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Montero-Castaño, A.; Vilà, M. Impact of landscape alteration and invasions on pollinators: A meta-analysis. J. Ecol. 2012, 100, 884–893. [Google Scholar] [CrossRef]
- McKechnie, I.M.; Thomsen, C.J.M.; Sargent, R.D. Forested field edges support a greater diversity of wild pollinators in lowbush blueberry (Vaccinium angustifolium). Agric. Ecosyst. Environ. 2017, 237, 154–161. [Google Scholar] [CrossRef]
- Purvis, E.; Meehan, M.; Lindo, Z. Agricultural field margins provide food and nesting resources to bumble bees (Bombus spp., Hymenoptera: Apidae) in Southwestern Ontario, Canada. Insect Conserv. Diver. 2020, 13, 219–228. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Koh, I.; Lonsdorf, E.V.; Williams, N.M.; Brittain, C.; Isaacs, R.; Gibbs, J.; Ricketts, T.H. Modelling the status, trends, and impacts of wild bee abundance in the United States. Proc. Natl. Acad. Sci. USA 2016, 113, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar]
- Schumacher, W. Gefährdete Ackerwildkräuter können auf ungespritzten Feldrändern erhalten werden. Mitt. LÖLF 1984, 9, 14–20. [Google Scholar]
- Jörg, E. Field Margin-strip programmes. In Proceedings of the Technical Seminar of the Landesanstalt für Pflanzenbau und Pflanzschutz, Mainz, Germany, 25–27 May 1994. Pretty Print. [Google Scholar]
- De Snoo, G.R. Unsprayed field margins: Effects on environment, biodiversity and agricultural practice. Landsc. Urban Plan. 1999, 46, 151–160. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Jacot, K.; Eggenschwiler, L.; Junge, X.; Luka, H.; Bosshard, A. Improved field margins for a higher biodiversity in agricultural landscapes. Asp. Appl. Biol. 2007, 87, 277–283. [Google Scholar]
- Hines, H.M.; Hendrix, S.D. Bumble bee (Hymenoptera: Apidae) diversity and abundance in tallgrass prairie patches: Effects of local and landscape floral resources. Environ. Entomol. 2005, 34, 1477–1484. [Google Scholar] [CrossRef]
- Colla, S.R.; Taylor-Pindar, A. Recovery Strategy for the Rusty-Patched Bumble Bee (Bombus Affinis) in Ontario; Ontario Recovery Strategy Series; Queens Printer for Ontario: Peterborough, ON, Canada, 2011. [Google Scholar]
- Morandin, L.A.; Kremen, C. Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 2013, 23, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.A.; Carrasco, A.; La-Spina, M.; Ibáñez, H.; Canomanuel, G.; Ortiz-Sánchez, F.J.; López, E.; Lacasa, A. Edges of natural vegetation to increase the diversity of wild bees in agricultural field margins. IOBC-WPRS Bull. 2014, 100, 117–121. [Google Scholar]
- Meek, B.; Loxton, D.; Sparks, T.; Pywell, R.; Pickett, H.; Nowakowski, M. The effect of arable field margin composition on invertebrate biodiversity. Biol. Conserv. 2002, 106, 259–271. [Google Scholar] [CrossRef]
- Aguado, L.O.; Viñuelas, E.; Ferreres, A. Guía de Polinizadores de la Península Ibérica y de Los Archipiélagos Balear y Canario; Ediciones Mundiprensa & Syngenta: Madrid, Spain, 2016. [Google Scholar]
- Amiet, F.; Herrmann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 3: Halictus, Lasioglossum; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2001. [Google Scholar]
- Amiet, F.; Hermann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 5: Ammobates, Ammobatoides, Anthophora, Biastes, Ceratina, Dasypoda, Epeoloides, Epeolus, Eucera, Macropis, Melecta, Melitta, Nomada, Pasites, Tetralonia, Thyreus, Xylocopa; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2007. [Google Scholar]
- Amiet, F.; Hermann, M.; Müller, A.; Neumeyer, R. Fauna Helvetica APIDAE 6: Andrena, Meliturga, Panurginus, Panurgus; Centre suisse de cartographie de la faune (CSCF Info Fauna) Schweizerische Entomological Gesellschaft (SEG/SES): Bern, Switzerland, 2010. [Google Scholar]
- Cobos, A. Fauna Ibérica de Coleópteros Buprestidae; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1986. [Google Scholar]
- Eizaguirre, S. Coleóptera, coccinellidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales (CSIC): Madrid, Spain, 2004; Volume 40. [Google Scholar]
- Fernández-Rubio, F. Guía de las Mariposas Diurnas de la Península Ibérica, Baleares, Canarias, Azores y Madeira-1. Libytheidae, Nymphalidae, Riodinidae y Lycaenidae; Ediciones Pirámide, S.A.: Madrid, Spain, 1991. [Google Scholar]
- Fernández-Rubio, F. Guía de las Mariposas Diurnas de la Península Ibérica, Baleares, Canarias, Azores y Madeira. 2. Papilionidae, Pieridae, Danaidae, Satyridae y Hesperiidae; Ediciones Pirámide, S.A.: Madrid, Spain, 1991. [Google Scholar]
- Fernández-Rubio, F. Clave para la determinación de las especies españolas del género Zygaena Fabricius, 1775 (Insecta: Lepidoptera). Graellsia 2006, 62, 3–12. [Google Scholar] [CrossRef]
- Martín-Piera, F.; López-Colón, J.I. Coleóptera Scarabaeoidea I. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales. CSIC: Madrid, Spain, 2000; Volume 14. [Google Scholar]
- Oosterbroek, P. The European Families of the Diptera. Identification, Diagnosis, Biology; KNNV Publishing: Utrecht, The Netherlands, 2006. [Google Scholar]
- Scheuchl, E. Illustrierte Bestimmungstabellen der Wildbienen. Band I: Anthophoridae; Preisinger KG: Landhut, Germany, 2000. [Google Scholar]
- Scheuchl, E.; Willner, W. Taschenlexikon der Wildbienen Mitteleuropas: Alle Arten im Portrait; Quelle & Meyer GmbH: Wiebelsheim, Germany, 2016. [Google Scholar]
- Séméria, Y.; Berland, L. Atlas des Néuroptères de France et d´Europe. Mégaloptères, Raphidioptères, Néuroptères Planipennes, Mécoptères; Société Nouvelle des Éditions Boubée: Paris, France, 1988. [Google Scholar]
- Vives, E. Coleóptera, cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2000; Volume 12. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed effects models and extensions in ecology with R. In Statistics for Biology and Health; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hilbe, J.M. Log negative binomial regression as a generalized linear model. Grad. Coll. Comm. Stat. 1993, 1024, 1–16. [Google Scholar]
- Cameron, A.C.; Trivedi, P.K. Econometric models based on count data. Comparisons and applications on some estimators and tests. J. Appl. Econom. 1986, 1, 29–53. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Siemann, E.; Tilman, D.; Haarstad, J.; Ritchie, M. Experimental tests of the dependence of arthropod diversity on plant diversity. Am. Nat. 1998, 152, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Duan, M.; Zhang, X.; Zhang, X.; Yu, Z.; Axmacher, J.C.; Stewart, A.; Ewers, R. Effects of plant diversity, habitat and agricultural landscape structure on the functional diversity of carabid assemblages in the North China Plain. Insect Conserv. Diver. 2015, 8, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Danne, A.; Thomson, L.J.; Sharley, D.J.; Penfold, C.M.; Hoffmann, A.A. Effects on native grass cover crops on beneficial and pest invertebrates in Australian vineyards. Pest Manag. 2010, 39, 970–978. [Google Scholar] [CrossRef]
- Eckert, M.; Mathulwe, L.L.; Gaigher, R.; Joubert-van der Merwe, L.; Pryke, J.S. Native cover crops enhance arthropod diversity in vineyards of the Cape Floristic Region. J. Insect Conserv. 2020, 24, 133–149. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Pérez-Moreno, I.; Marco-Mancebón, V.S. Ground cover management in a Mediterranean vineyard: Impact on insect abundance and diversity. Agric. Ecosyst. Environ. 2019, 283, 106571. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Effects of ground cover management on insect predators and pests in a Mediterranean vineyard. Insects 2019, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agric. Ecosyst. Environ. 2021, 307, 107222. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Veldtman, R.; Shenkute, A.G.; Tesfay, G.B.; Pirk, C.W.W.; Donaldson, J.S.; Nicolson, S.W. Natural and within farmland biodiversity enhances crop productivity. Ecol. Lett. 2011, 14, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Miranda-Barroso, L.; Aguado, O.; Falcó-Garí, J.V.; Lopez, D.; Schade, M.; Vasileiadis, V.; Peris-Felipo, F.J. Multifunctional areas as a tool to enhance biodiversity and promote conservation in alfalfa fields. J. Insect Biodiver. Syst. 2021, 7, 251–261. [Google Scholar]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706, 135839. [Google Scholar] [CrossRef] [PubMed]
- Dassou, A.G.; Dépigny, S.; Canard, E.; Vinatier, F.; Carval, D.; Tixier, P. Contrasting effects of plant diversity across arthropod trophic groups in plantain-based agroecosystems. Basic Appl. Ecol. 2016, 17, 11–20. [Google Scholar] [CrossRef]
- Vasileiadis, V.P.; Sattin, M.; Otto, S.; Veres, A.; Palinkas, Z.; Pons, X.; Kudsk, P.; van der Weide, R.; Czembor, E.; Moonen, C.; et al. Crop protection in European maize-based cropping systems: Current practices and recommendations for innovative integrated pest management. Agric. Syst. 2011, 104, 533–540. [Google Scholar] [CrossRef]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. Agric. Ecosyst. Environ. 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground cover management with mixtures of flowering plants to enhance insect pollinators and natural enemies of pests in olive groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Möller, G.; Keasar, T.; Shapira, I.; Möller, D.; Ferrante, M.; Segoli, M. Effect of weed management on the parasitoid community in Mediterranean vineyards. Biology 2021, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Reiff, J.M.; Kolb, S.; Entling, M.H.; Herndl, T.; Möth, S.; Walzer, A.; Kropf, M.; Hoffmann, C.; Winter, S. Organic farming and cover-crop management reduce pest predation in Austrian vineyards. Insects 2021, 12, 220. [Google Scholar] [CrossRef]
- Zanettin, G.; Bullo, A.; Pozzebon, A.; Burgio, G.; Duso, C. Influence of vineyard inter-row groundcover vegetation management on arthropod assembalges in the vineyards of North-Eastern Italy. Insects 2021, 12, 349. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province | Place | GPS Coordinates | Grape Variety | Field Area (ha) |
|---|---|---|---|---|
| Burgos | Aranda de Duero | 41°37′22.51″ N, 3°41′17.44″ W | Tempranillo | 5 |
| Madrid | Villamanta | 40°18′00.61″ N, 4°07′17.83″ W | Garnacha | 4 |
| Toledo | El Carpio de Tajo | 39°49′34.70″ N, 4°26′38.80″ W | Tempranillo | 5 |
| Toledo | Otero | 39°59′36.00″ N, 4°31′42.60″ W | Tempranillo | 4 |
| Valladolid | Sardón de Duero | 41°37′01.00″ N, 4°24′39.00″ W | Tempranillo | 4.5 |
| Aranda de Duero | El Carpio de Tajo | Otero | Sardón de Duero | Villamanta | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| with Cover | without Cover | with Cover | without Cover | with Cover | without Cover | with Cover | without Cover | with Cover | without Cover | |
| Genera | 86 | 78 | 75 | 72 | 82 | 74 | 106 | 91 | 101 | 85 |
| Individuals | 1053 | 899 | 552 | 508 | 1100 | 679 | 1211 | 707 | 726 | 515 |
| With Cover Plants | Without Cover Plants | ||||
|---|---|---|---|---|---|
| Genus | Individuals | % | Genus | Individuals | % |
| Coccinella | 301 | 6.35 | Coccinella | 235 | 7.10 |
| Apis | 285 | 6.01 | Eristalis | 153 | 4.63 |
| Eristalis | 268 | 5.65 | Sphaerophoria | 147 | 4.44 |
| Andrena | 223 | 4.70 | Apis | 141 | 4.26 |
| Chrysoperla | 212 | 4.47 | Chrysoperla | 122 | 3.69 |
| Total | 1289 | 27.18 | 798 | 24.12 | |
| Test | Poisson | Negative Binomial |
|---|---|---|
| Likelihood ratio (LR) | 2.29 | 0.75 *** |
| Deviance (D) | 15,453.7 *** | 11,598.6 *** |
| AIC | 15,469.7 | 11,616.6 |
| BIC | 15,520.5 | 11,673.8 |
| Source | Chisq | Df | p-Value | |
|---|---|---|---|---|
| Zone | 56.04 | 1 | 7.117 × 10−14 | *** |
| Year | 412.57 | 1 | <2.2 × 10−16 | *** |
| Zone:Year | 42 | 2 | 7.553 × 10−10 | *** |
| Random Effects | |||
| Groups | Name | Variance | Std. Dev. |
| Genus | (Intercept) | 2.40 | 1.55 |
| Farm | (Intercept) | 0.06 | 0.25 |
| Fixed Effects | |||
| Parameter | Estimate | Std. Error | |
| (Intercept) | −0.78 | 0.19 | *** |
| Zone:without | −0.90 | 0.10 | *** |
| Year:2014 | 0.32 | 0.09 | *** |
| Year:2015 | 1.07 | 0.08 | *** |
| Zone:without-Year: 2014 | 0.84 | 0.13 | *** |
| Zone:without-Year: 2015 | 0.58 | 0.13 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peris-Felipo, F.J.; Santa, F.; Aguado, O.; Falcó-Garí, J.V.; Iborra, A.; Schade, M.; Brittain, C.; Vasileiadis, V.; Miranda-Barroso, L. Enhancement of the Diversity of Pollinators and Beneficial Insects in Intensively Managed Vineyards. Insects 2021, 12, 740. https://doi.org/10.3390/insects12080740
Peris-Felipo FJ, Santa F, Aguado O, Falcó-Garí JV, Iborra A, Schade M, Brittain C, Vasileiadis V, Miranda-Barroso L. Enhancement of the Diversity of Pollinators and Beneficial Insects in Intensively Managed Vineyards. Insects. 2021; 12(8):740. https://doi.org/10.3390/insects12080740
Chicago/Turabian StylePeris-Felipo, Francisco Javier, Fernando Santa, Oscar Aguado, José Vicente Falcó-Garí, Alicia Iborra, Michael Schade, Claire Brittain, Vasileios Vasileiadis, and Luis Miranda-Barroso. 2021. "Enhancement of the Diversity of Pollinators and Beneficial Insects in Intensively Managed Vineyards" Insects 12, no. 8: 740. https://doi.org/10.3390/insects12080740
APA StylePeris-Felipo, F. J., Santa, F., Aguado, O., Falcó-Garí, J. V., Iborra, A., Schade, M., Brittain, C., Vasileiadis, V., & Miranda-Barroso, L. (2021). Enhancement of the Diversity of Pollinators and Beneficial Insects in Intensively Managed Vineyards. Insects, 12(8), 740. https://doi.org/10.3390/insects12080740