
Microhabitat Selection for Overwintering: Overwintering Conditions of Three Jumping Spiders (Pellenes tripunctatus, P. nigrociliatus, and Attulus penicillatus) Living in Terrestrial Shells in the Czech Republic


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
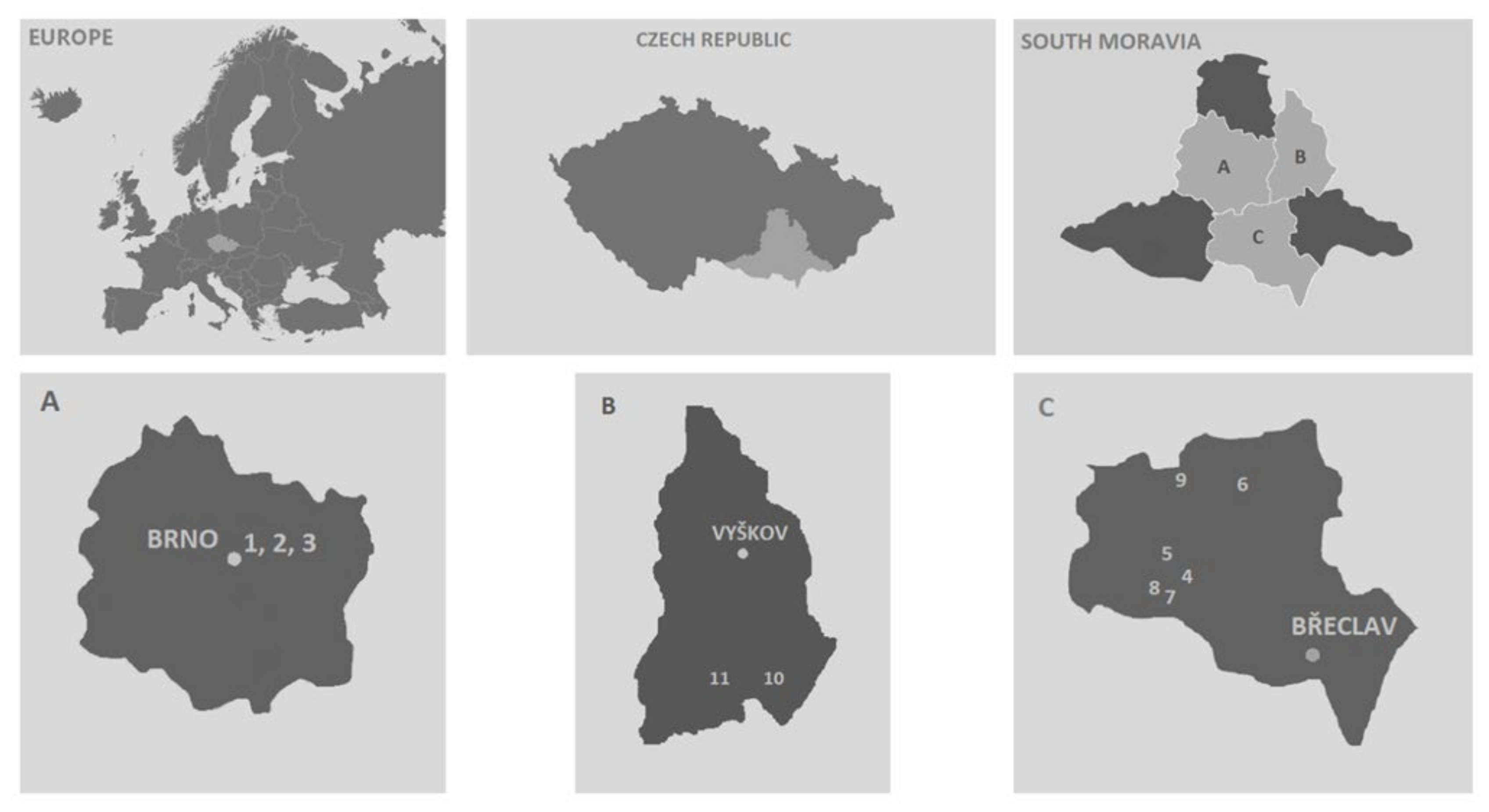
2.1. Localities
2.2. Field Survey
2.3. Analyses
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vertainen, L.; Alatalo, R.V.; Mappes, J.; Parri, S. Sexual differences in growth strategies of the wolf spider Hygrolycosa rubrofasciata. Evol. Ecol. 2000, 14, 595–610. [Google Scholar] [CrossRef]
- Krause, R.H.; Buse, J.; Matern, A.; Schröder, B.; Härdtle, W.; Assmann, T. Eresus kollari (Araneae: Eresidae) calls for heathland management. J. Arachnol. 2011, 39, 384–392. [Google Scholar] [CrossRef]
- Liedtke, J.; Schneider, J.M. Social makes smart: Rearing conditions affect learning and social behaviour in jumping spiders. Anim. Cogn. 2017, 20, 1093–1106. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J. Spatial distribution and microhabitat preference of Psecas chapoda (Peckham & Peckham) (Araneae, Salticidae). J. Arachnol. 2005, 33, 124–134. [Google Scholar]
- Rossa-Feres, D.; Romero, G.Q.; Gonçalves-de-Freitas, E.; Feres, R.J.F. Reproductive behavior and seasonal occurrence of Psecas viridipurpureus (Salticidae, Araneae). Braz. J. Biol. 2000, 60, 221–228. [Google Scholar]
- De Omena, P.M.; Romero, G.Q. Using visual cues of microhabitat traits to find home: The case study of a bromeliad-living jumping spider (Salticidae). Behav. Ecol. 2010, 21, 690–695. [Google Scholar] [CrossRef]
- Jackson, R.R. Nests of Phidippus johnsoni (Araneae: Salticidae): Characteristics, pattern of occupation, and function. J. Arachnol. 1979, 7, 47–58. [Google Scholar]
- Jackson, R.R.; Griswold, C.E. Nest associates of Phidippus johnsoni (Araneae, Salticidae). J. Arachnol. 1979, 7, 59–67. [Google Scholar]
- Jennings, D.T.; Cutler, B. Intrafloral partitioning of nesting sites by Eris militaris and Synageles canadensis (Araneae: Salticidae). Entomol. News 2019, 128, 314–319. [Google Scholar] [CrossRef]
- Mooney, K.A.; Haloin, J.R. Nest site fidelity of Paraphidippus aurantia (Salticidae). J. Arachnol. 2005, 34, 241–243. [Google Scholar] [CrossRef]
- Hoefler, C.D.; Jakob, E.M. Jumping spiders in space: Movement patterns, nest site fidelity and the use of beacons. Anim. Behav. 2006, 71, 109–116. [Google Scholar] [CrossRef]
- Eubanks, M.D.; Miller, G.L. Life cycle and habitat preference of the facultatively arboreal wolf spider, Gladicosa pulchra (Araneae, Lycosidae). J. Arachnol. 1992, 20, 157–164. [Google Scholar]
- Herberstein, M.E. Web placement in sympatric linyphiid spiders (Arachnida, Araneae): Individual foraging decisions reveal inter-specific competition. Acta Oecologica 1998, 19, 67–71. [Google Scholar] [CrossRef]
- Carrel, J. Ecology of two burrowing wolf spiders (Araneae: Lycosidae) syntopic in Florida Scrub: Burrow/body size relationships and habitat preferences. J. Kans. Entomol. Soc. 2003, 76, 16–30. [Google Scholar]
- Michalko, R.; Košulič, O.; Hula, V.; Surovcová, K. Niche differentiation of two sibling wolf spider species, Pardosa lugubris and Pardosa alacris, along a canopy openness gradient. J. Arachnol. 2016, 44, 46–51. [Google Scholar] [CrossRef]
- Moreno-Rueda, G.; Marfil-Daza, C.; Ortiz-Sanchez, F.J.; Melic, A. Weather and the use of empty gastropod shells by arthropods. Ann. Sociétéentomol. Fr. 2008, 44, 373–377. [Google Scholar] [CrossRef]
- Bogusch, P.; Roháček, J.; Baňař, P.; Astapenková, A.; Kouklík, O.; Pech, P.; Janšta, P.; Heller, K.; Hlaváčková, L.; Heneberg, P.; et al. The presence of high numbers of empty shells in anthropogenic habitats is insufficient to attract shell adopters among the insects. Insect Conserv. Divers. 2019, 12, 193–205. [Google Scholar] [CrossRef]
- Szinetár, C.; Gál, Z.; Eichardt, J. Spiders in snail shells in different Hungarian habitats. Misc. Zool. Hung. 1998, 12, 67–75. [Google Scholar]
- Horn, H. Die Bedeutung leerer Schneckengehäuse für die Überwinterung und das Brutverhalten von Pellenes nigrociliatus L. Koch, 1874 in Steppenrasenformationen (Araneae: Salticidae). Beiträge Zur Nat. Forsch. Südwestdeutschland 1980, 39, 167–175. [Google Scholar]
- Bauchhenss, E. Überwinternde Spinnen aus Schneckenhäusern. Arachnol. Mitt. 1995, 9, 57–60. [Google Scholar] [CrossRef]
- Mikulska, I. Parental care in a rare spiders Pellenes nigrociliatus (L. Koch) var. bilunulata Simon. Nature 1961, 190, 365–366. [Google Scholar]
- Niedobová, J.; Hula, V.; Košulič, O. Prázdné ulity plžů a tajemství, která skrývají. Živa 2013, 60, 26–28. [Google Scholar]
- Metzner, H. Jumping spiders (Arachnida: Araneae: Salticidae) of the world. Available online: https://www.jumping-spiders.com (accessed on 14 September 2022).
- Nentwig, W.; Blick, T.; Bosmans, R.; Gloor, D.; Hänggi, A.; Kropf, C. Spiders of Europe. Version 10.2022. Available online: https://www.araneae.nmbe.ch (accessed on 14 September 2022). [CrossRef]
- World Spider Catalog (2022). World Spider Catalog. Version 23.5. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 14 September 2022). [CrossRef]
- Buchar, J.; Růžička, V. Catalogue of Spiders of the Czech Republic; Peres: Prague, Czech Republic, 2002; p. 351. [Google Scholar]
- Kůrka, A.; Řezáč, M.; Macek, R.; Dolanský, J. Pavouci České Republiky, 1st ed.; Academia: Prague, Czech Republic, 2014; p. 554. [Google Scholar]
- Buskirk, R.E. Sociality in arachnida. In Social Insect, 2nd ed.; Hermann, H.R., Ed.; Academic Press: New York, NY, USA, 1981; Volume 2, pp. 281–367. [Google Scholar]
- Herberstein, M.E. Spider Behaviour: Flexibility and Versatility, 1st ed.; Cambridge University Press: New York, NY, USA, 2011; p. 391. [Google Scholar]
- Jackson, R.R. Communal jumping spiders (Araneae: Salticidae) from Kenya: Interspecific nest complexes, cohabitation with web-building spiders, and intraspecific interactions. N. Z. J. Zool. 1986, 13, 13–26. [Google Scholar] [CrossRef]
- Jackson, R.R.; Pollard, S.D.; Macnab, A.M.; Cooper, K.J. The complex communicatory behaviour of Marpissa marina, a New Zealand jumping spider (Araneae: Salticidae). N. Z. J. Zool. 1990, 17, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Nelson, X.J.; Jackson, R.R. Complex display behaviour during the intraspecific interactions of myrmecomorphic jumping spiders (Araneae, Salticidae). J. Nat. Hist. 2007, 41, 1659–1678. [Google Scholar] [CrossRef] [Green Version]
- Mackovčin, P.; Jatiová, M.; Slavík, P.; Demek, J.; Grulich, V.; Hort, L.; Hudec, K.; Kříž, H.; Laštůvka, Z.; Petruš, J.; et al. Brněnsko, 1st ed.; Agentura ochrany přírody a krajiny ČR: Prague, Czech Republic, 2008; p. 932. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using Canoco 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; p. 300. [Google Scholar]
- Řezáč, M.; Kůrka, A.; Růžička, V.; Heneberg, P. Red list of Czech spiders: 3rd edition, adjusted according to evidence-based national conservation priorities. Biologia 2015, 70, 645–666. [Google Scholar] [CrossRef]
- Štempáková, K.; Hula, V. Life strategies of jumping spiders (Araneae: Salticidae) of genus Pellenes—A possible explanation of the unusual sociability. In MendelNet 2017, Proceedings of the International PhD Students Conference, Brno, Czech Republic, 8–9 November 2017; Cerkal, R., Belcredi, N.B., Eds.; Mendel University in Brno: Brno, Czech Republic, 2017. [Google Scholar]
- Bilde, T.; Lubin, Y. Kin recognition and cannibalism in a subsocial spiders. J. Evol. Biol. 2001, 14, 959–966. [Google Scholar] [CrossRef]
- Krafft, B.; Cookson, L.J. The role of silk in the behaviour and sociality of spiders. Psyche 2012, 2012, 1–25. [Google Scholar] [CrossRef]
- Yip, E.C.; Rayor, L.S. Maternal care and subsocial behaviour in spiders. Biol. Rev. 2014, 89, 427–449. [Google Scholar] [CrossRef]
- Lubin, Y.; Bilde, T. The Evolution of sociality in spiders. Adv. Study Behav. 2007, 37, 83–145. [Google Scholar]
- Kim, K.W.; Krafft, B.; Choe, J.C. Cooperative prey capture by young subsocial spiders. Behav. Ecol. Socio-Biol. 2005, 59, 92–100. [Google Scholar] [CrossRef]
- Grinsted, L.; Bilde, T.; d’Ettorre, P. Cuticular hydrocarbons as potential kin recognition cues in a subsocial spider. Behav. Ecol. 2011, 22, 1187–1194. [Google Scholar] [CrossRef]
- Gundermann, J.-L.; Horel, A.; Roland, C. Costs and benefits of maternal care in a subsocial spider, Coelotes terrestris. Ethology 1997, 103, 915–925. [Google Scholar]
- Kim, K.W.; Roland, C.; Horel, A. Functional value of matriphagy in the spider Amaurobius ferox. Ethology 2000, 106, 729–742. [Google Scholar] [CrossRef]
- Jones, J.C.; Oldroyd, B.P. Nest thermoregulation in social insect. Adv. Insect Physiol. 2006, 33, 153–191. [Google Scholar]
- Ożgo, M.; Komorowska, A. Shell banding polymorphism in Cepaea vindobonensis in relation to habitat in southeastern Poland. Malacologia 2009, 51, 81–88. [Google Scholar] [CrossRef]
- Bellmann, H. Zur Nutzungleerer Schneckenhäuser durch Arthropoden. Verh. Der Ges. Für Okol. 1999, 29, 169–172. [Google Scholar]
- Hula, V.; Niedobová, J.; Košulič, O. Overwintering of spiders in land-snail shells in South Moravia (Czech Republic). Acta Musei Morav. Sci. Biol. 2009, 94, 1–12. [Google Scholar]
- Štempáková, K.; Hula, V. Overwintering of spiders in terrestrial mollusc’s shells in the wider area of Vranov nad Topľou (Slovakia). Acta Univ. Agric. Silvic. Mendel. Brun. 2017, 65, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Nyffeler, M.; Bonte, D. Where have all the spiders gone? Observations of a dramatic population density decline in the once very abundant garden spider, Araneus diadematus (Araneae: Araneidae), in the Swiss Midland. Insects 2020, 11, 248. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Locality | GPS | Geology | Conservation Management | Elevation m a.s.l. | Number of Shells | Number of Quadrats |
|---|---|---|---|---|---|---|
| Stránská skála | 49°11′28.600″ N, 16°40′33.264″ E | Limestone quarry | mown, grazed | 255–310 | 95 | 9 |
| Bílá hora | 49°11′38.845″ N, 16°39′34.528″ E | Limestone outcrop | mown | 270–299 | 75 | 6 |
| Hády | 49°13′2.122″ N, 16°40′7.623″ E | Limestone quarry | trampling | 310–424 | 67 | 9 |
| Milovická stráň | 48°50′40.492″ N, 16°41′59.390″ E | Loess | grazed | 200–297 | 8 | 5 |
| Děvín-Kotel-Soutěska | 48°52′3.454″ N, 16°38′50.178″ E | Limestone outcrop | grazed, mown | 280–555 | 84 | 22 |
| Kamenný vrch u Kurdějova | 48°57′54.796″ N, 16°45′14.138″ E | Loess | mown, grazed | 300–340 | 36 | 11 |
| Janičův vrch | 48°48′39.428″ N, 16°39′31.473″ E | Limestone quarry | trampling | 264–306 | 186 | 8 |
| Růžový kopec | 48°49′16.353″ N, 16°37′28.167″ E | Loess, limestone | mown | 260–280 | 6 | 2 |
| Pouzdřanská step—Kolby | 48°56′49.344″ N, 16°38′34.960″ E | Loess | mown, grazed | 220–284 | 1 | 1 |
| Malhotky | 49°8′53.258″ N, 17°3′19.783″ E | Loess | mown | 235–300 | 330 | 69 |
| Člupy | 49°9′7.751″ N, 16°57′27.793″ E | Loess | mown | 250–300 | 2 | 2 |
| Species | Pellenes tripunctatus | Pellenes nigrociliatus | Attulus penicillatus | Heliophanus sp. | Euryopis quinqueguttata | Talavera aequipes | Heliophanus flavipes | Cheiracanthium pennyi | Neottiura bimaculata | Clubiona sp. | Haplodrassus sp. | Drassodes sp. | Theridiidae | Total | Occupancy of Shells |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 113/889 | |||||||||||||||
| Number of individuals | 56 | 52 | 11 | 7 | 6 | 6 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 146 | 12.71% |
| Model | 1st Axis | All Axes | ||||
|---|---|---|---|---|---|---|
| Expl. Var. | Adj. Expl. Var. | F | p | F | p | |
| Locality | 5.83 | 4.76 | 5.7 | 0.001 | 5.4 | 0.001 |
| Snail species | 1.33 | 1.11 | 6.0 | 0.006 | 6.0 | 0.006 |
| Shell density | 2.13 | 0.65 | 1.4 | 0.262 | 1.4 | 0.262 |
| Vegetation cover | 2.19 | 1.52 | 3.3 | 0.007 | 3.3 | 0.007 |
| Shell characteristics | 2.89 | 1.89 | 3.0 | 0.003 | 2.9 | 0.003 |
| Model | 1st Axis | All Axes | ||||
|---|---|---|---|---|---|---|
| Expl. Var. | Adj. Expl. Var. | F | p | F | p | |
| Locality | 25.40 | 20.64 | 3.8 | 0.001 | 5.3 | 0.001 |
| Snail species | 19.14 | 15.77 | 5.4 | 0.001 | 5.7 | 0.001 |
| Shell density | 8.26 | 0.00 | 0.4 | 0.661 | 0.5 | 0.761 |
| Vegetation cover | 23.98 | 13.61 | 1.4 | 0.001 | 2.3 | 0.003 |
| Shell characteristics | 27.53 | 11.63 | 0.8 | 0.002 | 1.7 | 0.004 |
| Locality | Type of Shell | Species of Spiders | ||||
|---|---|---|---|---|---|---|
| P. tripunctatus | P. nigrociliatus | A. penicillatus | Other Species of Spiders | Total | ||
| Malhotky | Caucasotachea | 3 | 3 | |||
| Malhotky | Caucasotachea | 2 | 2 | |||
| Malhotky | Caucasotachea | 1 | Neottiura bimaculata | 2 | ||
| Malhotky | Caucasotachea | 3 | 3 | |||
| Malhotky | Caucasotachea | 4 | Theridiidae sp. | 5 | ||
| Malhotky | Caucasotachea | 1 | Euryopis quinqueguttata | 2 | ||
| Malhotky | Caucasotachea | 1 | 3× Euryopis quinqueguttata | 4 | ||
| Malhotky | Caucasotachea | 1 | 2× Euryopis quinqueguttata | 3 | ||
| Malhotky | Caucasotachea | 6 | 6 | |||
| Stránská skála | Xerolenta | 1 | Heliophanus flavipes | 2 | ||
| Stránská skála | Xerolenta | 2 | 2 | |||
| Stránská skála | Xerolenta | 1 | Talavera aequipes, Heliophanus sp. | 3 | ||
| Stránská skála | Xerolenta | 3 | 3 | |||
| Stránská skála | Xerolenta | 1 | Talavera aequipes | 2 | ||
| Stránská skála | Xerolenta | 2× Heliophanus sp. | 2 | |||
| Stránská skála | Xerolenta | 2× Heliophanus sp. | 2 | |||
| Děvín | Xerolenta | 2 | 2 | |||
| Janičův vrch | Xerolenta | 2 | 2 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziváková, K.; Fric, Z.F.; Hula, V. Microhabitat Selection for Overwintering: Overwintering Conditions of Three Jumping Spiders (Pellenes tripunctatus, P. nigrociliatus, and Attulus penicillatus) Living in Terrestrial Shells in the Czech Republic. Insects 2022, 13, 950. https://doi.org/10.3390/insects13100950
Dziváková K, Fric ZF, Hula V. Microhabitat Selection for Overwintering: Overwintering Conditions of Three Jumping Spiders (Pellenes tripunctatus, P. nigrociliatus, and Attulus penicillatus) Living in Terrestrial Shells in the Czech Republic. Insects. 2022; 13(10):950. https://doi.org/10.3390/insects13100950
Chicago/Turabian StyleDziváková, Kristína, Zdeněk Faltýnek Fric, and Vladimír Hula. 2022. "Microhabitat Selection for Overwintering: Overwintering Conditions of Three Jumping Spiders (Pellenes tripunctatus, P. nigrociliatus, and Attulus penicillatus) Living in Terrestrial Shells in the Czech Republic" Insects 13, no. 10: 950. https://doi.org/10.3390/insects13100950
APA StyleDziváková, K., Fric, Z. F., & Hula, V. (2022). Microhabitat Selection for Overwintering: Overwintering Conditions of Three Jumping Spiders (Pellenes tripunctatus, P. nigrociliatus, and Attulus penicillatus) Living in Terrestrial Shells in the Czech Republic. Insects, 13(10), 950. https://doi.org/10.3390/insects13100950