

Spider Assemblages of Tree Trunks and Tree Branches in Three Developmental Phases of Primeval Oak–Lime–Hornbeam Forest in the Białowieża National Park


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Statistical Analysis
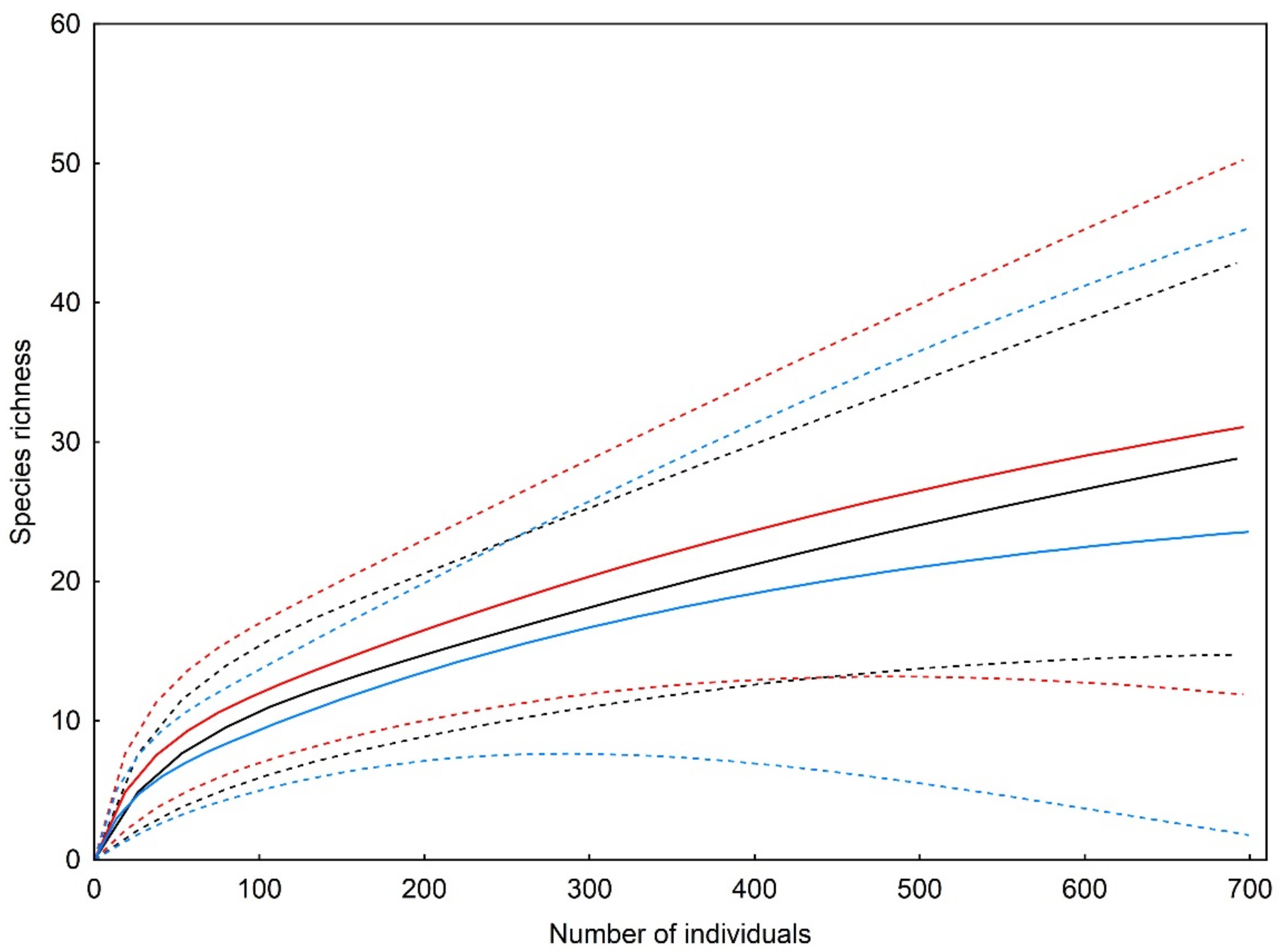
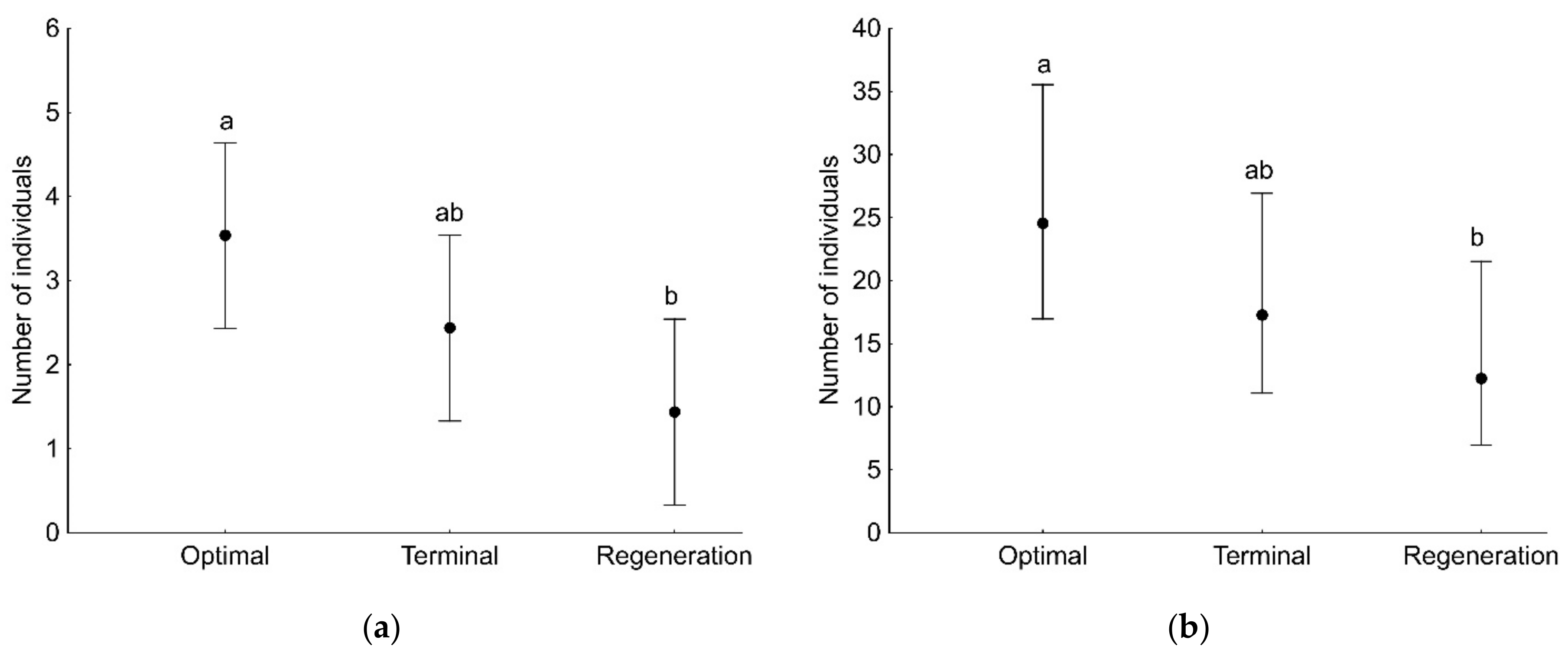
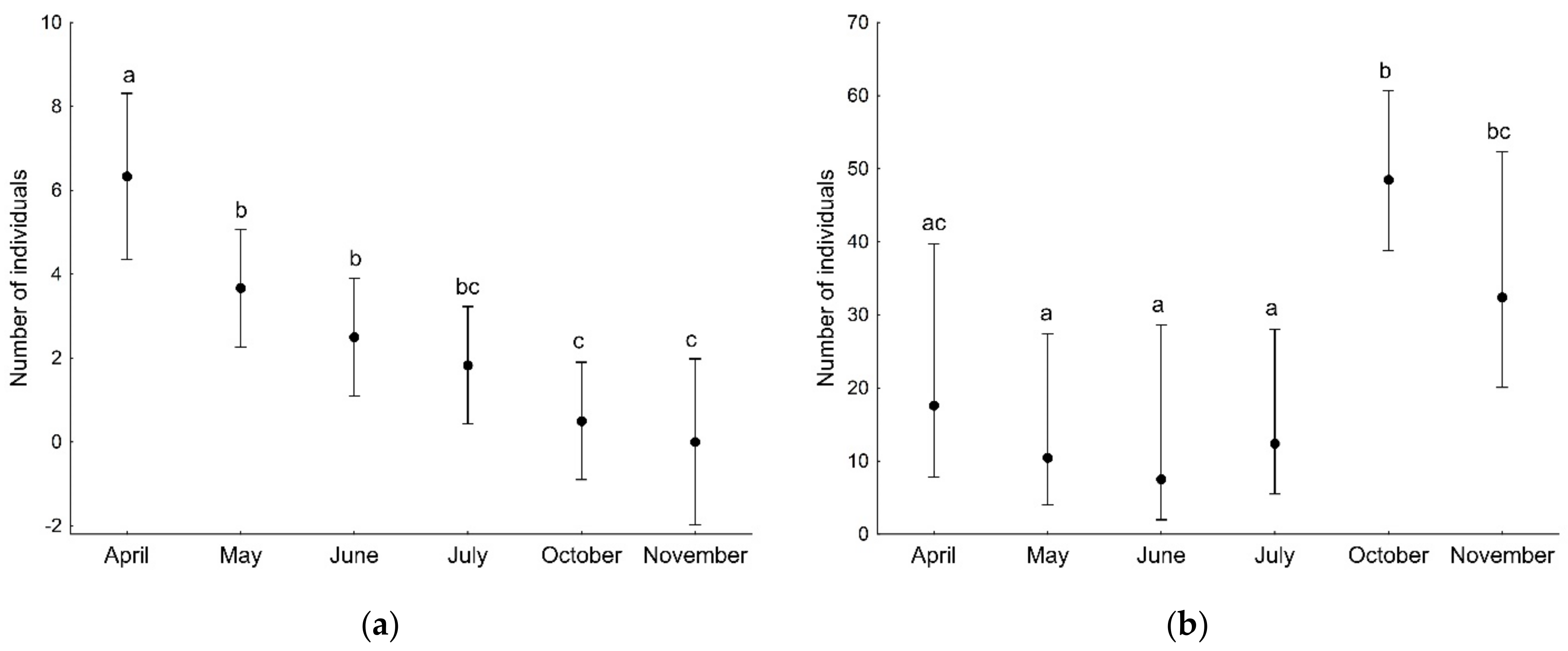
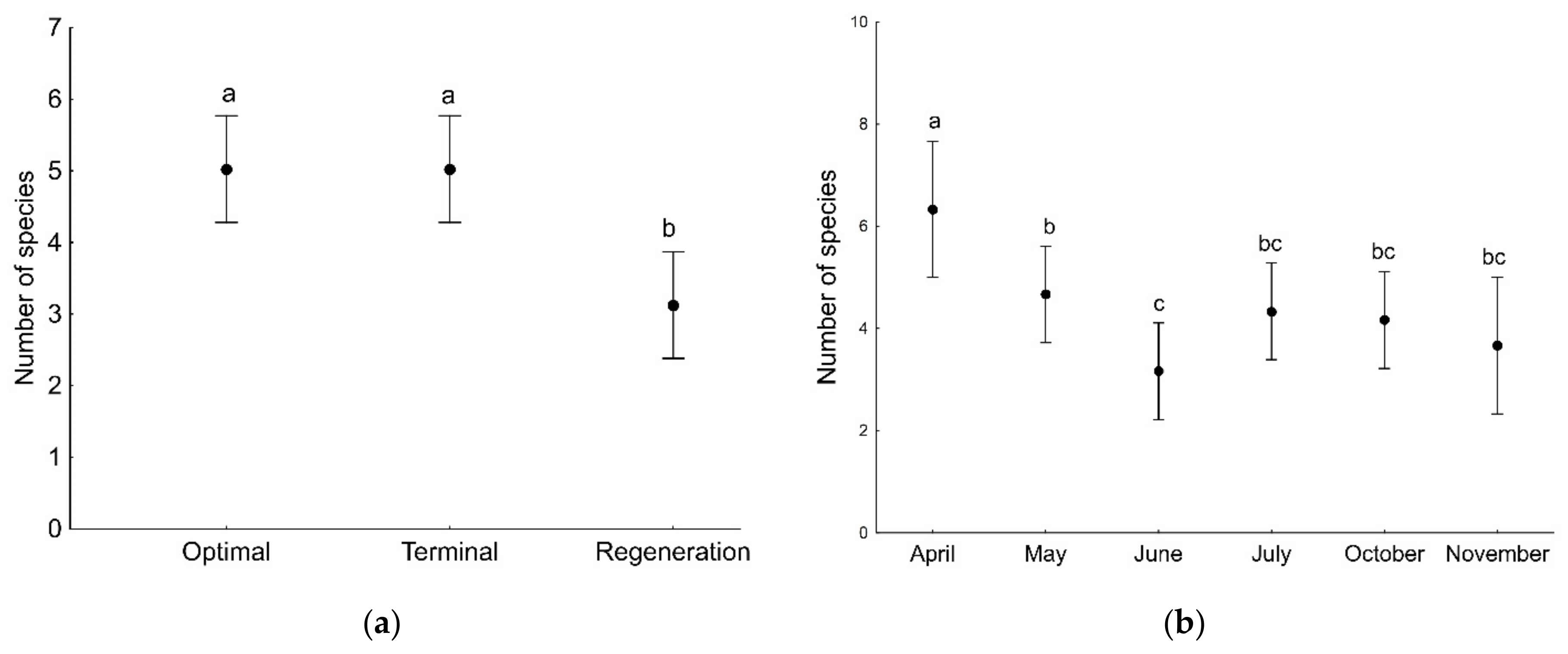
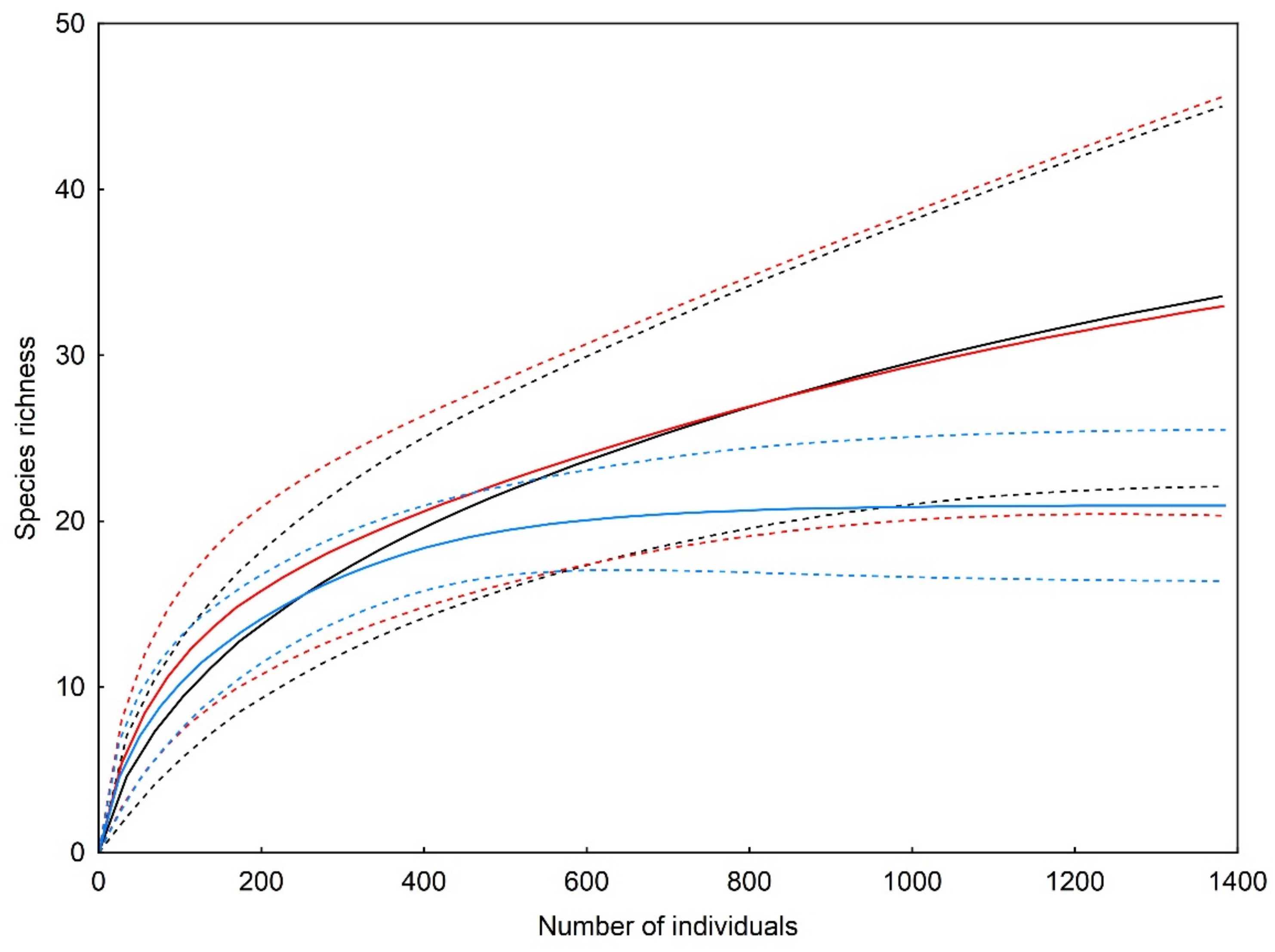
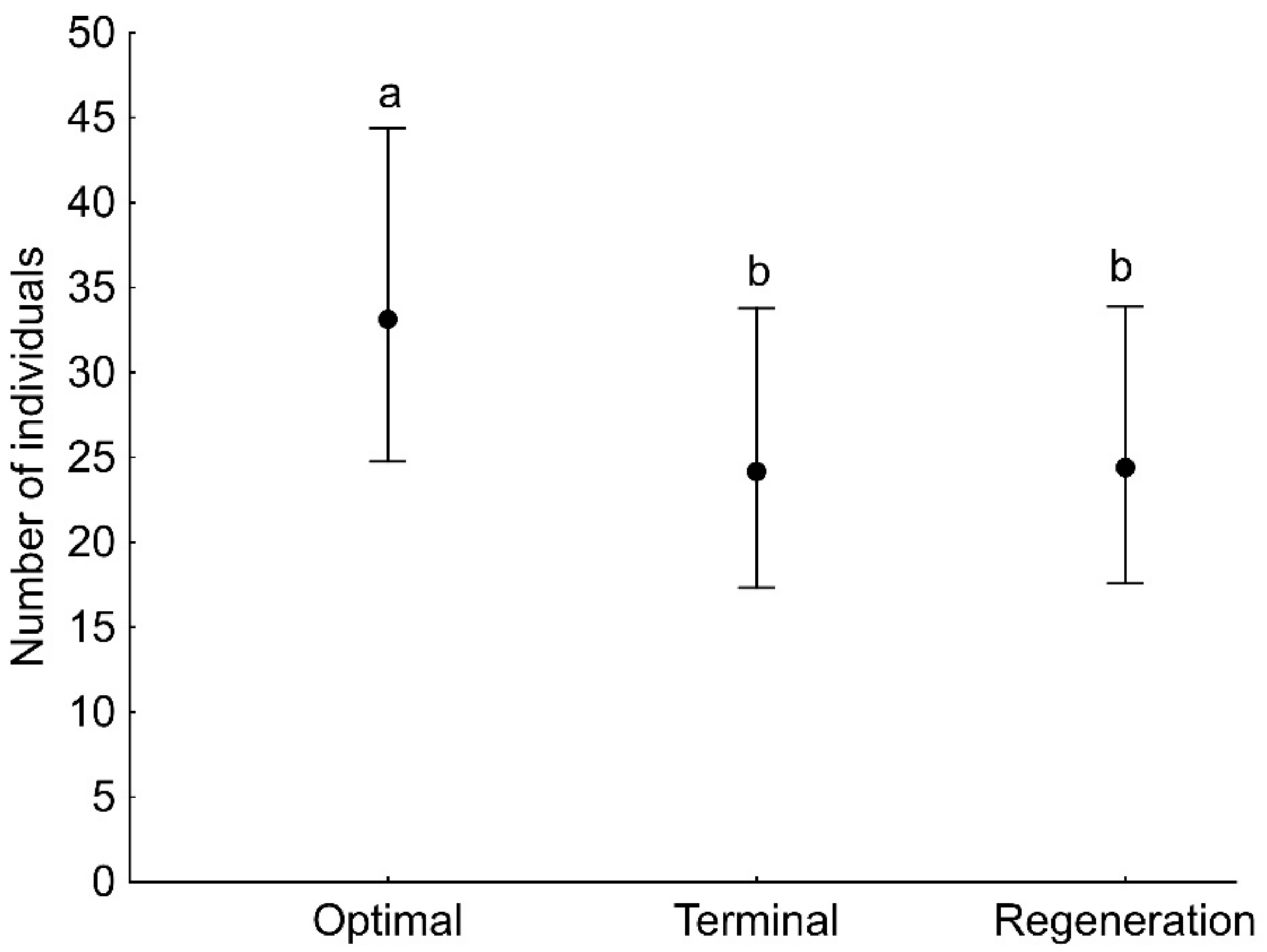
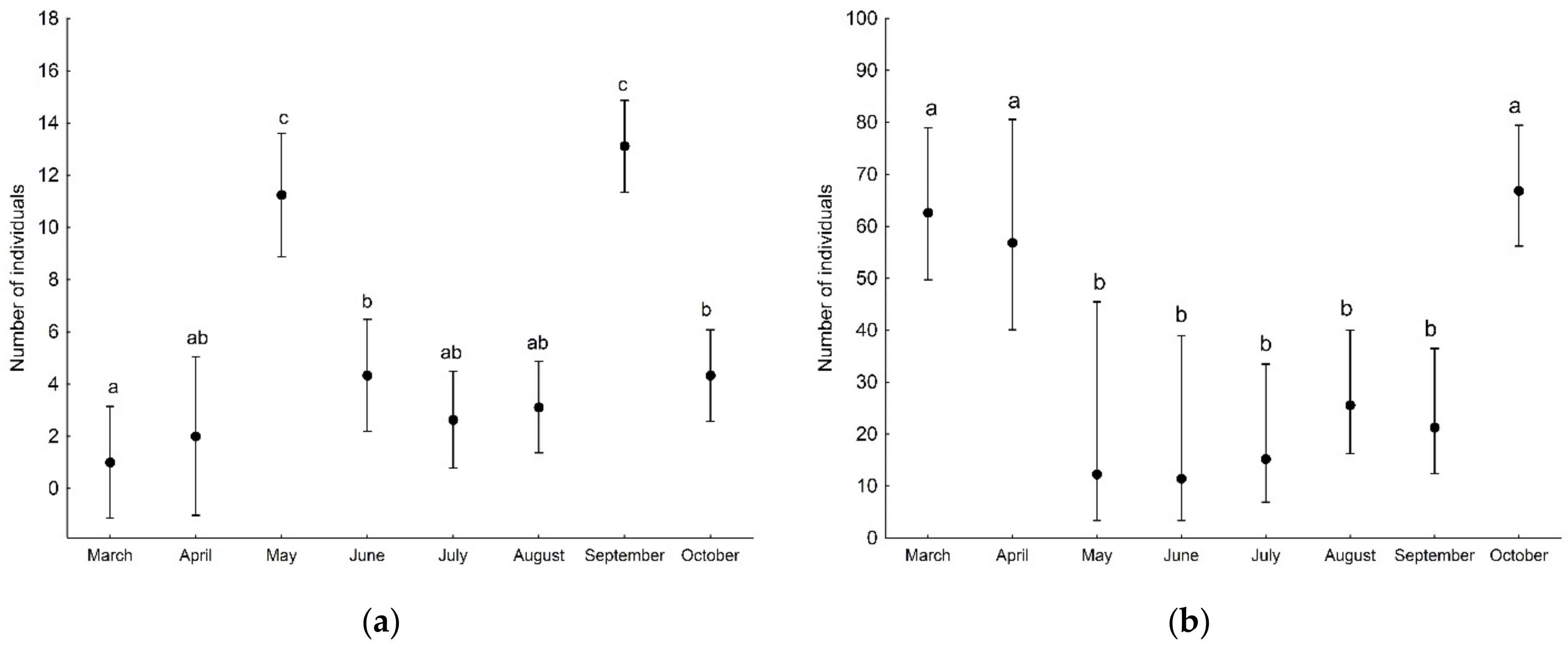
3. Results
3.1. Spiders of Tree Branches
3.2. Spiders of Tree Trunks
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicolai, V. The bark of trees: Thermal properties, microclimate and fauna. Oecologia 1986, 69, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Koponen, S.; Rinne, V.; Clayhills, T. Arthropods on oak branches in SW Finland, collected by a new trap type. Entomol. Fenn. 1997, 8, 177–183. [Google Scholar] [CrossRef]
- Floren, A. Abundance and ordinal composition of arboreal arthropod communities of various trees in old primary and managed forests. In Canopy Arthropod Research in Europe, 1st ed.; Floren, A., Schmidl, J., Eds.; Bioform Entomology: Nuremberg, Germany, 2008; pp. 279–298. [Google Scholar]
- Sebek, P.; Vodka, S.; Bogusch, P.; Pech, P.; Tropek, R.; Weiss, M.; Zimova, K.; Cizek, L. Open-grown trees as key habitats for arthropods in temperate woodlands: The diversity, composition, and conservation value of associated communities. For. Ecol. Manag. 2016, 380, 172–181. [Google Scholar] [CrossRef]
- Blick, T. Abundant and rare spiders on tree trunks in German forests (Arachnida, Araneae). Arachnol. Mitt. 2011, 40, 5–14. [Google Scholar] [CrossRef]
- Stańska, M.; Stański, T.; Bartos, M. Spider assemblages of tree branches in managed and primeval deciduous stand of the Białowieża Forest. Forests 2022, 13, 5. [Google Scholar] [CrossRef]
- Otto, S.; Floren, A. The spider fauna (Araneae) of tree canopies in the Białowieża Forest. Fragm. Faun. 2007, 50, 57–70. [Google Scholar] [CrossRef]
- Mupepele, A.-C.; Müller, T.; Dittrich, M.; Floren, A. Are Temperate Canopy Spiders Tree-Species Specific? PLoS ONE 2014, 9, e86571. [Google Scholar] [CrossRef]
- Tomiałojć, L. Characteristics of old growth in the Białowieża Forest, Poland. Nat. Area. J. 1991, 11, 7–18. [Google Scholar]
- Tomiałojc’, L.; Wesołowski, T. Diversity of the Białowieża Forest avifauna in space and time. J. Ornith. 2004, 145, 8–92. [Google Scholar]
- Bobiec, A.; Van der Burgt, H.; Meijer, K.; Zuyderduyn, C.; Haga, J.; Vlaanderen, B. Rich deciduous forests in Białowieża as a dynamic mosaic of developmental phases: Premises for nature conservation and management. For. Ecol. Manag. 2000, 130, 159–175. [Google Scholar] [CrossRef]
- Miścicki, S. Dynamics of the natural development phases of stands in the Białowieski National Park. Sylwan 2012, 156, 616–626. [Google Scholar]
- Miścicki, S. Using developmentl stages of forest stands as a basis for forest inventory. Sylwan 1994, 4, 29–39. [Google Scholar]
- Oxbrough, A.G.; Gittings, T.; O’Halloran, J.; Giller, P.S.; Smith, G.F. Structural indicators of spider communities across the forest plantation cycle. For. Ecol. Manag. 2005, 212, 171–183. [Google Scholar] [CrossRef]
- Mullen, K.; O’Halloran, J.; Breen, J.; Giller, P.; Pithon, J.; Helly, T. Distribution and composition of carabid beetle (Coleoptera, Carabidae) communities across the plantation forest cycle—Implications for managements. For. Ecol. Manag. 2008, 256, 624–632. [Google Scholar]
- Purchart, L.; Tuf, I.H.; Hula, V.; Suchomel, J. Arthropod assemblages in Norway Spruce monocultures during a forest cycle—A multi-taxa approach. For. Ecol. Manag. 2013, 306, 42–51. [Google Scholar] [CrossRef]
- Trojan, P.; Bańkowska, R.; Chudzicka, E.; Pilipiuk, I.; Skibińska, E.; Sterzyńska, M.; Wytwer, J. Secondary succession of fauna in the pine forests of Puszcza Białowieska. Fragm. Faun. 1994, 37, 1–104. [Google Scholar] [CrossRef][Green Version]
- Stańska, M.; Stański, T. Plant-dwelling spider communities of three developmental phases in primeval oak-lime-hornbeam forest in the Białowieża National Park, Poland. Eur. Zool. J. 2021, 88, 706–717. [Google Scholar] [CrossRef]
- Samu, F.; Lengyel, G.; Szita, É.; Bidló, A.; Ódor, P. The effect of forest stand characteristics on spider diversity and species composition in deciduous-coniferous mixed forests. J. Arachnol. 2014, 42, 135–141. [Google Scholar] [CrossRef]
- Košulič, O.; Michalko, R.; Hula, V. Impact of canopy openness on spider communities: Implications for conservation management of formerly coppiced oak forests. PLoS ONE 2016, 11, e0148585. [Google Scholar] [CrossRef]
- Stańska, M.; Stański, T.; Gładzka, A.; Bartos, M. Spider assemblages of hummocks and hollows in a primeval alder carr in the Białowieża National Park—Effect of vegetation structure and soil humidity. Pol. J. Ecol. 2016, 64, 564–577. [Google Scholar] [CrossRef]
- Esquivel-Gómez, L.; Abdala-Roberts, L.; Pinkus-Rendón, M.; Parra-Tabla, V. Effects of tree species diversity on a community of weaver spiders in a tropical forest plantation. Biotropica 2017, 49, 63–70. [Google Scholar] [CrossRef]
- Stańska, M.; Stański, T.; Hawryluk, J. Spider assemblages on tree trunks in primeval deciduous forests of the Białowieża National Park in eastern Poland. Ent. Fenn. 2018, 29, 75–85. [Google Scholar] [CrossRef]
- Gallé, R.; Gallé-Szpisjak, N.; Zsigmond, A.-R.; Könczey, B.; Urák, I. Tree species and microhabitat affect forest bog spider fauna. Eur. J. For. Res. 2021, 140, 691–702. [Google Scholar] [CrossRef]
- Malumbres-Olarte, J.; Vink, C.J.; Ross, J.G.; Cruickshank, R.H.; Paterson, A.M. The role of habitat complexity on spider communities in native alpine grasslands of New Zealand. Insect Conserv. Diver. 2013, 6, 124–134. [Google Scholar] [CrossRef]
- Pierre, J.I., St.; Kovalenko, K.E. Effect of habitat complexity attributes on species richness. Ecosphere 2014, 5, 22. [Google Scholar] [CrossRef]
- Áviala, A.C.; Stenert, C.; Rodrigues, E.N.L.; Maltchik, L. Habitat structure determines spider diversity in highland ponds. Ecol. Res. 2017, 32, 359–367. [Google Scholar] [CrossRef]
- Hamřik, T.; Košulič, O. Impact of small-scale conservation management methods on spider assemblages in xeric grassland. Agr. Ecosyst. Environ. 2021, 307, 107225. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples; Version 9.1.0.; University of Colorado: Boulder, CO, USA, 2019. [Google Scholar]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1998; p. 144. [Google Scholar]
- Hutcheson, T.P. A test for comparing diversities based on the Shannon formula. J. Theor. Biol. 1970, 29, 151–154. [Google Scholar] [CrossRef]
- Hauser, A.; Attrill, M.; Cotton, P.A. Effects of habitat complexity on the diversity and abundance of macrofauna colonising artificial kelp holdfasts. Mar. Ecol. Prog. Ser. 2006, 325, 93–100. [Google Scholar] [CrossRef]
- Lengyel, S.; Déri, E.; Magura, T. Species richness responses to structural or compositional habitat diversity between and within grassland patches: A multi-taxon approach. PLoS ONE 2016, 11, e0149662. [Google Scholar] [CrossRef]
- Oxbrough, A.G.; Gittings, T.; O’Halloran, J.; Giller, P.S.; Kelly, T.C. The influence of open space on ground-dwelling spider assemblages within plantation forests. For. Ecol. Manag. 2006, 237, 404–417. [Google Scholar] [CrossRef]
- Stenchly, K.; Clough, Y.; Tscharntke, T. Spider species richness in cocoa agroforestry systems, comparing vertical strata, local management and distance to forest. Agric. Ecosyst. Environ. 2012, 149, 189–194. [Google Scholar] [CrossRef]
- Wunderlich, J. Mitteleuropäische Spinnen (Araneae) der Baumrinde. Zeitschrift für angewandte. Entomologie 1982, 94, 9–21. [Google Scholar]
- Horváth, R.; Szinetár, C. Study of the bark-dwelling spiders (Araneae) on black pine (Pinus nigra) I. Misc. Zool. Hung. 1998, 12, 77–83. [Google Scholar]
- Koponen, S. Spiders from groves in the southwestern archipelago of Finland (Araneae). Rev. Ibérica Aracnol. 2008, 15, 97–104. [Google Scholar]
- Machač, O.; Tuf, I.H. Spiders and harvestmen on tree trunks obtained by three sampling methods. Arachnol. Mitt. 2016, 51, 67–72. [Google Scholar] [CrossRef]
- Szinetár, C.; Horváth, R. A review of spiders on tree trunks in Europe (Araneae). Acta Zool. Bulg. 2005, Suppl. 1, 221–257. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family/Genus/Species | Optimal Phase | Terminal Phase | Regeneration Phase | |||
|---|---|---|---|---|---|---|
| Ad./Juv. | Months | Ad./Juv. | Months | Ad./Juv. | Months | |
| Anyphaenidae | ||||||
| Anyphaena accentuata | (12%) -/32 | IV–VII, X, XI | (20%) 1/37 | IV–VII, X, XI | (36%) 1/49 | IV–VII, X, XI |
| Araneidae | ||||||
| Araneus diadematus | -/1 | VII | ||||
| Araniella sp. | -/1 | -/5 | ||||
| Cyclosa conica | 4/5 | IV, V, VII, X | 1/7 | IV, V, X, XI | 2/- | IV |
| Clubionidae | ||||||
| Clubiona sp. | -/2 | -/2 | ||||
| Linyphidae | ||||||
| Diplocephalus picinus | 1/- | VI | ||||
| Entelecara acuminata | 1/- | VII | 1/- | VII | ||
| Helophora insignis | -/5 | VII | -/2 | VII | ||
| Linyphia triangularis | 1/- | VII | ||||
| Linyphiidae un. | -/6 | -/12 | -/3 | |||
| Neriene clathrata | -/1 | VII | ||||
| Neriene emphana | (6%) 2/15 | IV, VI, VII | 2/7 | IV–VII, XI | -/1 | V |
| Neriene montana | -/1 | X | -/4 | VII, X | 1/- | V |
| Neriene peltata | (6%) 9/7 | IV, V, XI | (26%) 6/42 | IV–VI, X, XI | (8%) -/11 | X |
| Neriene sp. | -/20 | -/3 | -/1 | |||
| Pityohyphantes phrygianus | -/1 | X | ||||
| Porrhomma pygmaeum | 5/- | IV | 3/- | V | ||
| Tapinocyba insecta | 1/- | IV | ||||
| Tapinocyba pallens | 1/- | IV | ||||
| Trematocephalus cristatus | (42%) -/111 | V, X, XI | (22%) -/42 | X, XI | (37%) 1/50 | IV, V, X, XI |
| Philodromidae | ||||||
| Philodromus dispar | 1/- | V | ||||
| Philodromus sp. | -/7 | -/6 | -/1 | |||
| Tetragnathidae | ||||||
| Metellina sp. | -/2 | -/4 | -/5 | |||
| Metellina mengei | 2/- | X | 1/- | X | ||
| Tetragnatha montana | 1/- | V | ||||
| Tetragnatha sp. | -/8 | -/9 | -/7 | |||
| Theridiidae | ||||||
| Enoplognatha ovata | (9%) 6/19 | V, VI | (9%) 7/10 | V–VII | (8%) 3/8 | V–VII |
| Robertus scoticus | 1/- | IV | ||||
| Theridiidae un. | -/1 | |||||
| Theridion sp. | -/7 | -/12 | -/7 | |||
| Theridion varians | 1/- | VII | ||||
| Thomisidae | ||||||
| Diaea dorsata | (14%) 1/36 | IV–VII, X | (7%) 1/12 | IV–VII, XI | (4%) -/5 | IV–VII |
| Ozyptila sp. | -/1 | |||||
| Xysticus lanio | 1/- | V | ||||
| Xysticus sp. | -/1 | -/1 | ||||
| Total no. of individuals | 34/286 | 23/213 | 13/156 | |||
| Optimal Phase | Terminal Phase | Regeneration Phase | |
|---|---|---|---|
| Observed richness | 17 | 16 | 11 |
| Estimates | |||
| Chao1 ± SD | 41 ± 31 | 40 ± 31 | 19 ± 12 |
| Chao2 ± SD | 53 ± 44 | 45 ± 36 | 27 ± 21 |
| Jackknife1 ± SD | 25 ± 3 | 23 ± 2 | 16 ± 2 |
| Jackknife2 | 32 | 29 | 20 |
| Michaelis–Menten | 22 | 20 | 15 |
| Sampling completeness | 41% | 40% | 58% |
| Effect | Wald χ2 | df | p |
|---|---|---|---|
| Abundance of adult individuals | |||
| Intercept | 53.95 | 1 | <0.001 |
| Developmental phase | 7.21 | 2 | 0.027 |
| Sampling month | 31.66 | 5 | <0.001 |
| Abundance of juvenile individuals | |||
| Intercept | 235.53 | 1 | <0.001 |
| Developmental phase | 7.91 | 2 | 0.019 |
| Sampling month | 29.18 | 5 | <0.001 |
| Species richness | |||
| Intercept | 375.06 | 1 | <0.001 |
| Developmental phase | 17.36 | 2 | <0.001 |
| Sampling month | 16.01 | 5 | 0.007 |
| Family/Genus/Species | Optimal Phase | Terminal Phase | Regeneration Phase | |||
|---|---|---|---|---|---|---|
| Ad./Juv. | Months | Ad./Juv. | Months | Ad./Juv. | Months | |
| Agelenidae | ||||||
| Agelenidae un. | -/1 | |||||
| Coelotes atropos | 5/- | VIII, IX | 5/6 | IV, VI–IX | 2/- | VII, IX |
| Amaurobiidae | ||||||
| Amaurobius fenestralis | (32%) 52/146 | III–X | (31%) 62/106 | III–X | (25%) 38/74 | III–X |
| Anyphaenidae | ||||||
| Anyphaena accentuata | (49%) 4/299 | III–V, VII–X | (38%) 6/197 | III–V, VII–X | (52%) 7/227 | III–VI, VIII–X |
| Araneidae | ||||||
| Araneidae un. | -/1 | |||||
| Cyclosa conica | -/2 | IV | ||||
| Nuctenea umbratica | 2/- | VIII, IX | ||||
| Clubionidae | ||||||
| Clubiona caerulescens | 1/- | V | ||||
| Clubiona lutescens | 3/- | VII, IX | 2/- | VII, VIII | 4/- | VII, IX |
| Clubiona sp. | -/110 | -/33 | -/4 | |||
| Clubiona subsultans | 1/- | X | 3/- | X | ||
| Dictynidae | ||||||
| Dictyna sp. | -/1 | |||||
| Gnaphosidae | ||||||
| Haplodrassus cognatus | 1/- | III | 1/- | VI | 1/- | V |
| Haplodrassus sp. | -/24 | -/48 | -/44 | |||
| Linyphiidae | ||||||
| Agyneta ramosa | 1/- | VI | ||||
| Drapetisca socialis | 18/7 | VI–X | 11/13 | VI–X | 13/3 | V–IX |
| Helophora insignis | 1/- | VIII | 1/- | VIII | ||
| Labulla thoracica | 1/5 | VI–VIII | -/3 | VII, VIII | 1/- | IX |
| Lepthyphantes minutus | 2/- | VIII, IX | 7/1 | VII–IX | ||
| Lepthyphantes sp. | -/7 | |||||
| Linyphiidae un. | -/15 | -/22 | -/39 | |||
| Lophomma punctatum | 1/- | III | ||||
| Neriene clathrata | 1/- | VI | ||||
| Neriene montana | -/4 | III, IV, VIII, X | -/15 | IV, VIII–X | 2/8 | III, V, VI, VIII, X |
| Neriene sp. | -/2 | |||||
| Savignia frontata | 1/- | X | ||||
| Trematocephalus cristatus | -/11 | III, IV, X | -/4 | III | ||
| Lycosidae | ||||||
| Piratula hygrophila | 1/- | IX | ||||
| Mimetidae | ||||||
| Ero furcata | 1/- | V | ||||
| Philodromidae | ||||||
| Philodromus sp. | -/11 | -/19 | -/13 | |||
| Pisauridae | ||||||
| Dolomedes fimbriatus | -/1 | X | ||||
| Salticidae | ||||||
| Neon reticulatus | 3/- | V, VII | ||||
| Neon sp. | -/1 | |||||
| Salticidae un. | -/1 | -/1 | ||||
| Segestriidae | ||||||
| Segestria senoculata | 1/10 | V, VIII–X | (9%) 5/43 | V–X | (6%) 3/24 | V–X |
| Tetragnathidae | ||||||
| Metellina merianae | 1/- | VII | ||||
| Tetragnatha sp. | -/2 | -/1 | -/2 | |||
| Theridiidae | ||||||
| Dipoena nigroreticulata | 1/- | VI | 1/1 | III, X | ||
| Enoplognatha ovata | 2/1 | VI | 3/3 | V–VIII | 1/4 | VI, IX |
| Steatoda bipunctata | 1/2 | VI, IX | 2/11 | V–IX | 3/15 | III, VII–X |
| Theridion mystaceum | 1/- | V | 3/- | V, VI | 3/- | V–VII |
| Theridion sp. | -/28 | -/34 | -/58 | |||
| Platnickina tincta | 1/2 | VII, X | -/1 | IV | 2/1 | VI, IX |
| Theridion varians | -/2 | III | ||||
| Thomisidae | ||||||
| Diaea dorsata | (5%) -/32 | III, IV, X | -/18 | III, IV, X | -/7 | III, IV, X |
| Ozyptila praticola | 1/- | VIII | ||||
| Ozyptila sp. | -/5 | -/3 | ||||
| Xysticus sp. | -/3 | -/3 | ||||
| Total no. of individuals | 97/732 | 119/576 | 85/537 | |||
| Optimal Phase | Terminal Phase | Regeneration Phase | |
|---|---|---|---|
| Observed richness | 24 | 23 | 19 |
| Estimates | |||
| Chao1 ± SD | 35 ± 10 | 55 ± 40 | 22 ± 3 |
| Chao2 ± SD | 43 ± 16 | 42 ± 19 | 21 ± 2 |
| Jackknife1 ± SD | 34 ± 4 | 32 ± 3 | 24 ± 2 |
| Jackknife2 | 42 | 38 | 23 |
| Michaelis–Menten | 33 | 27 | 23 |
| Sampling completeness | 69% | 42% | 86% |
| Effect | Wald χ2 | df | p |
|---|---|---|---|
| Abundance of adult individuals | |||
| Intercept | 183.62 | 1 | <0.001 |
| Developmental phase | 3.84 | 2 | 0.146 |
| Sampling month | 138.80 | 7 | <0.001 |
| Abundance of juvenile individuals | |||
| Intercept | 583.29 | 1 | <0.001 |
| Developmental phase | 6.81 | 2 | 0.033 |
| Sampling month | 51.06 | 7 | <0.001 |
| Species richness | |||
| Intercept | 362.97 | 1 | <0.001 |
| Developmental phase | 2.14 | 2 | 0.342 |
| Sampling month | 4.11 | 7 | 0.767 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stańska, M.; Stański, T. Spider Assemblages of Tree Trunks and Tree Branches in Three Developmental Phases of Primeval Oak–Lime–Hornbeam Forest in the Białowieża National Park. Insects 2022, 13, 1115. https://doi.org/10.3390/insects13121115
Stańska M, Stański T. Spider Assemblages of Tree Trunks and Tree Branches in Three Developmental Phases of Primeval Oak–Lime–Hornbeam Forest in the Białowieża National Park. Insects. 2022; 13(12):1115. https://doi.org/10.3390/insects13121115
Chicago/Turabian StyleStańska, Marzena, and Tomasz Stański. 2022. "Spider Assemblages of Tree Trunks and Tree Branches in Three Developmental Phases of Primeval Oak–Lime–Hornbeam Forest in the Białowieża National Park" Insects 13, no. 12: 1115. https://doi.org/10.3390/insects13121115
APA StyleStańska, M., & Stański, T. (2022). Spider Assemblages of Tree Trunks and Tree Branches in Three Developmental Phases of Primeval Oak–Lime–Hornbeam Forest in the Białowieża National Park. Insects, 13(12), 1115. https://doi.org/10.3390/insects13121115