
Control of Aculops lycopersici with the Predatory Mite Transeius montdorensis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mite Colonies
2.2. Experiment in Potted Plants
2.3. Experiment in Cages
2.4. Experiment in a Greenhouse
2.5. Data Analyses
3. Results
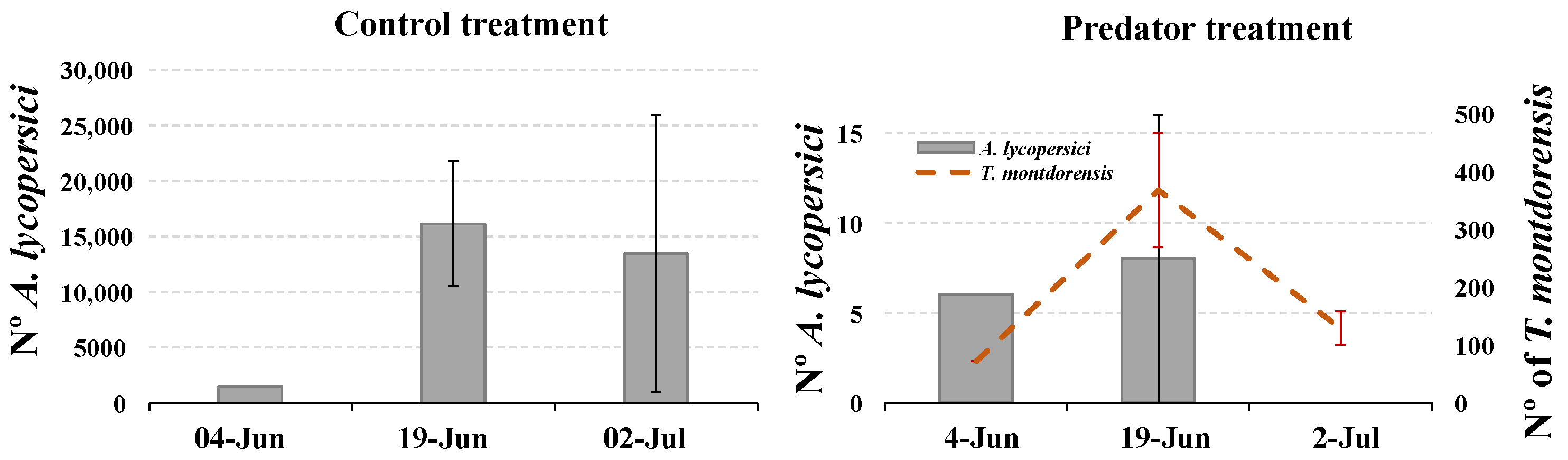
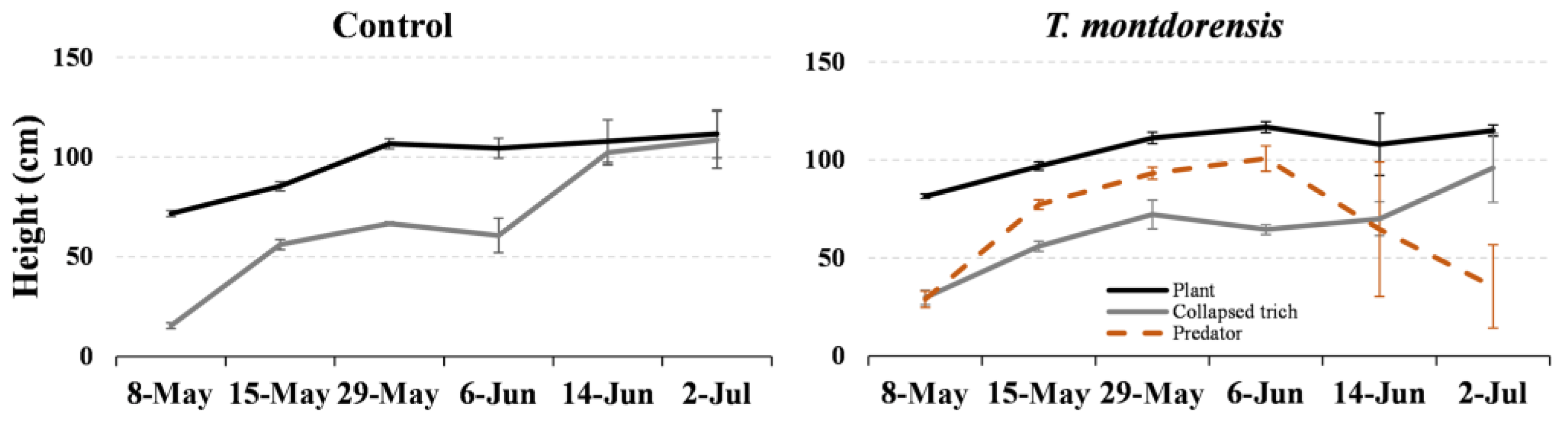
3.1. Experiment in Potted Plants
3.2. Experiment in Cages
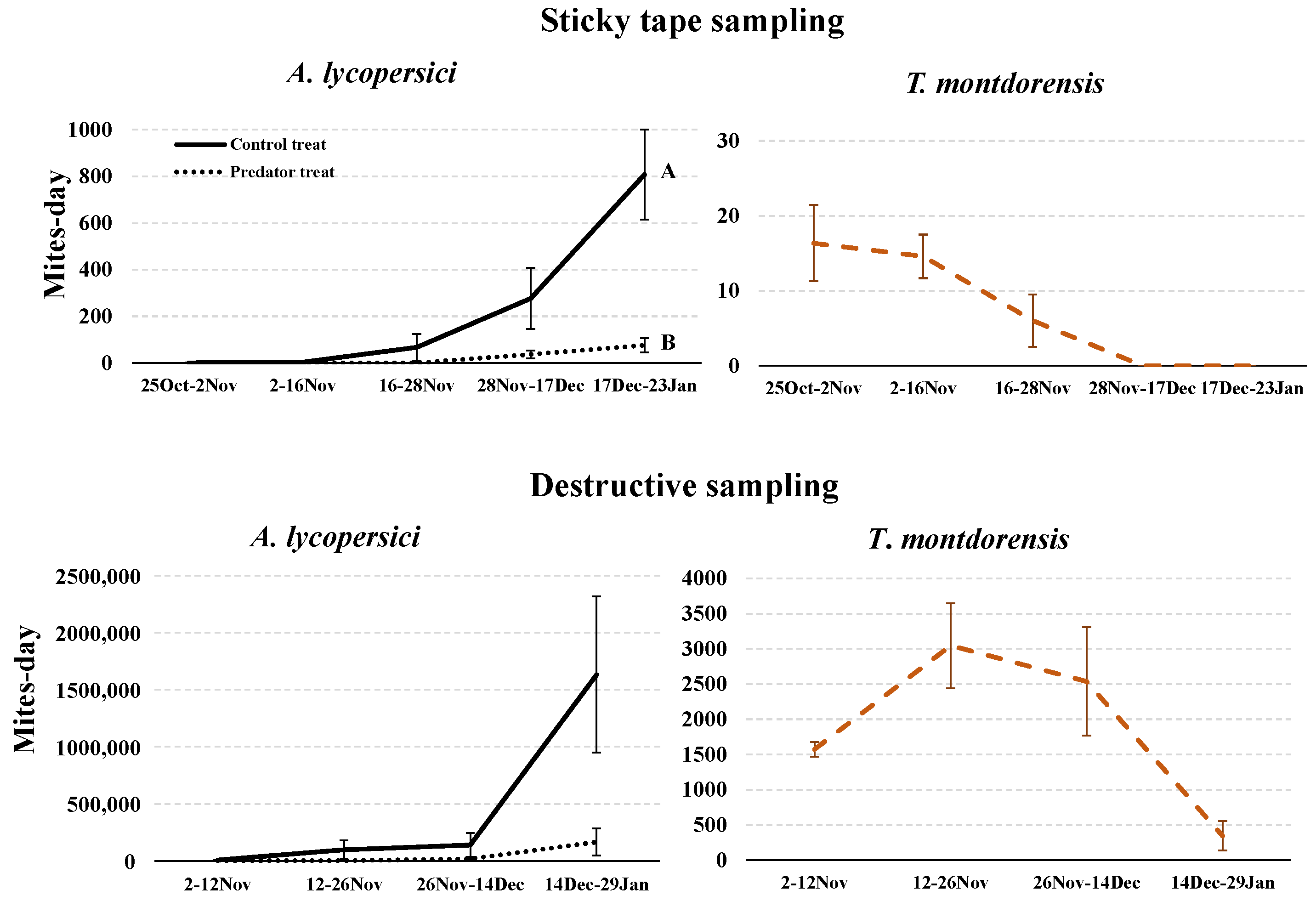
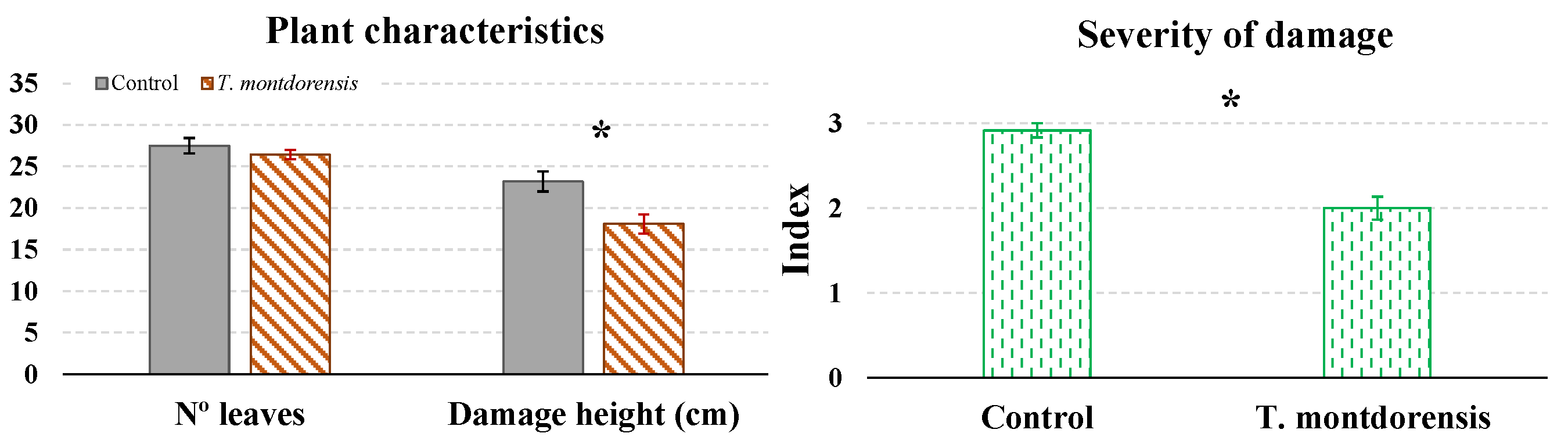
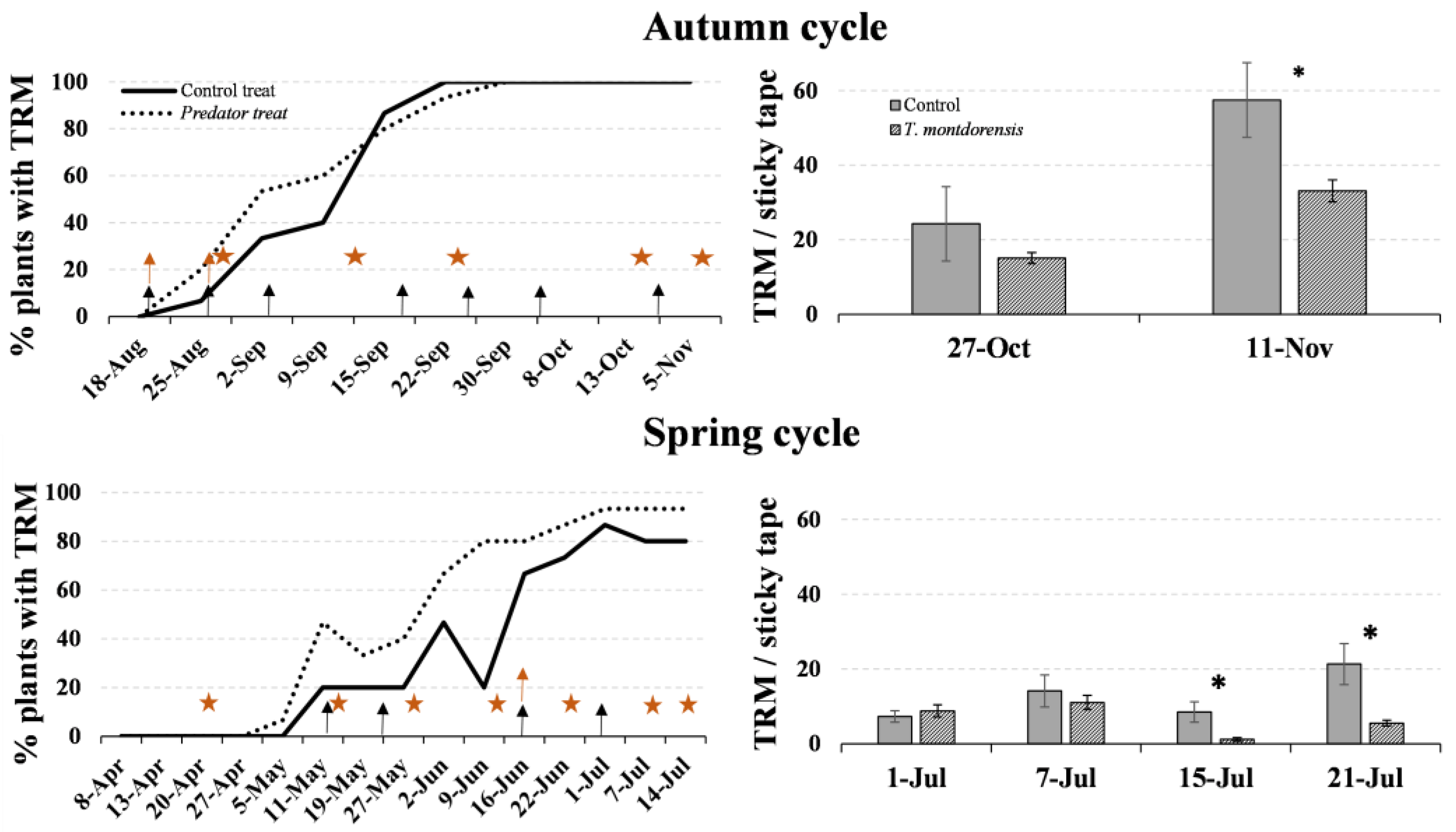
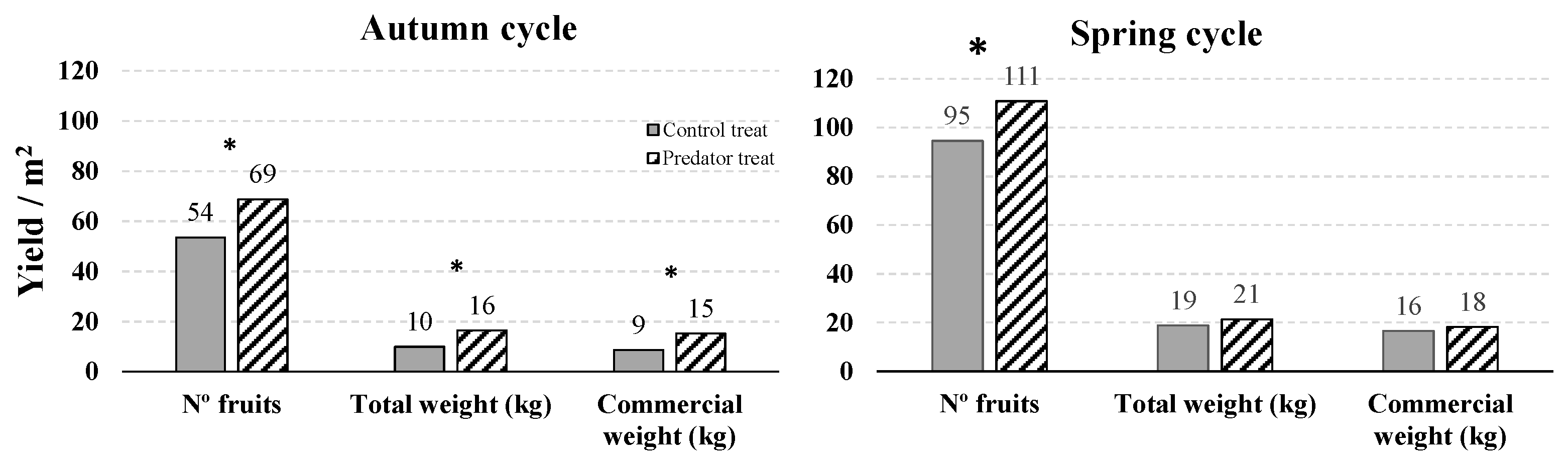
3.3. Experiment in a Greenhouse
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zalom, F.G.; Kitzmiller, J.; Wilson, L.T.; Gutierrez, P. Observation of tomato russet mite (Acari: Eriophyidae) damage symptoms in relation to tomato plant development. J. Econ. Entomol. 1986, 79, 940–942. [Google Scholar] [CrossRef]
- Haque, M.M.; Kawai, A. Population growth of tomato russet mite, Aculops lycopersici (Acari: Eriophyidae) and its injury effect on the growth of tomato plants. J. Acarol. Soc. Jpn. 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Vervaet, L.; De Vis, R.; De Clercq, P.; Van Leeuwen, T. Is the emerging mite pest Aculops lycopersici controllable? Global and genome-based insights in its biology and management. Pest Manag. Sci. 2021, 77, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Royalty, R.N.; Ring, T.M.P. Morphological analysis of damage to tomato leaflets by tomato russet mite (Acari: Eriophyidae). J. Econ. Entomol. 1988, 81, 816–820. [Google Scholar] [CrossRef]
- Royalty, R.N.; Ring, T.M.P. Reduction in photosynthesis of tomato leaflets caused by tomato russet mite (Acari: Eriophyidae). Environ. Entomol. 1989, 18, 256–260. [Google Scholar] [CrossRef]
- Duso, C.; Castagnoli, M.; Simoni, S.; Angeli, G. The impact of eriophyoids on crops: Recent issues on Aculus schlechtendali, Calepitrimerus vitis and Aculops lycopersici. Exp. Appl. Acarol. 2010, 51, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Al-Azzazy, M.M.; Alhewairini, S. Relationship between temperature and developmental rate of tomato russet mite Aculops lycopersici (Massee) (Acari: Eriophyideae) on tomato. J. Food Agric. Environ. 2018, 16, 18–23. [Google Scholar] [CrossRef]
- Kamau, A.W.; Mueke, J.M.; Khaemba, B.M. Resistance of tomato varieties to the tomato russet mite, Aculops lycopersici (Massee) (Acarina: Eriophyidae). Int. J. Trop. Insect Sci. 1992, 13, 351–356. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Bruin, J. Evolutionary Ecology: Life History Patterns, Food Plant Choice and Dispersal. In Eriophyoid Mites—Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 329–366. [Google Scholar]
- Aysan, E.; Kumral, N.A. Tritrophic relationships among tomato cultivars, the rust mite, Aculops lycopersici (Massee) (Eriophyidae), and its predators. Acarologia 2018, 58, 5–17. [Google Scholar] [CrossRef]
- van Houten, Y.M.; Glas, J.J.; Hoogerbrugge, H.; Rothe, J.; Bolckmans, K.J.F.; Simoni, S.; van Arkel, J.; Alba, J.M.; Kant, M.R.; Sabelis, M.W. Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Exp. Appl. Acarol. 2013, 60, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.Y.; Goodwin, S.; Wellham, T.M.; Barchia, I.M.; Spohr, L.J. Biological studies of the Australian predatory mite Typhlodromips montdorensis (Schicha) (Acari: Phytoseiidae), a potential biocontrol agent for western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Aust. J. Entomol. 2003, 42, 124–130. [Google Scholar] [CrossRef]
- Rahman, T.; Broughton, S.; Spafford, H. Can spinosad-resistant Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) be managed with spinosad and predatory mites (Acari)? Crop Prot. 2012, 42, 281–288. [Google Scholar] [CrossRef]
- Pfaff, A.; Gabriel, D.; Böckmann, E. Mitespotting: Approaches for Aculops lycopersici monitoring in tomato cultivation. Exp. Appl. Acarol. 2020, 80, 1–15. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. JMP Statistics and Graphics Guide. Version 14.2.0; SAS Institute: Cary, NC, USA, 2018. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castañé, C.; Alomar, O.; Rocha, A.; Vila, E.; Riudavets, J. Control of Aculops lycopersici with the Predatory Mite Transeius montdorensis. Insects 2022, 13, 1116. https://doi.org/10.3390/insects13121116
Castañé C, Alomar O, Rocha A, Vila E, Riudavets J. Control of Aculops lycopersici with the Predatory Mite Transeius montdorensis. Insects. 2022; 13(12):1116. https://doi.org/10.3390/insects13121116
Chicago/Turabian StyleCastañé, Cristina, Oscar Alomar, Alfred Rocha, Enric Vila, and Jordi Riudavets. 2022. "Control of Aculops lycopersici with the Predatory Mite Transeius montdorensis" Insects 13, no. 12: 1116. https://doi.org/10.3390/insects13121116
APA StyleCastañé, C., Alomar, O., Rocha, A., Vila, E., & Riudavets, J. (2022). Control of Aculops lycopersici with the Predatory Mite Transeius montdorensis. Insects, 13(12), 1116. https://doi.org/10.3390/insects13121116