
Enantiomeric Discrimination in Insects: The Role of OBPs and ORs


Abstract
Simple Summary
Abstract
1. Introduction
2. Chirality in Insect Olfaction
2.1. Semiochemistry
2.2. Chiral Pheromones
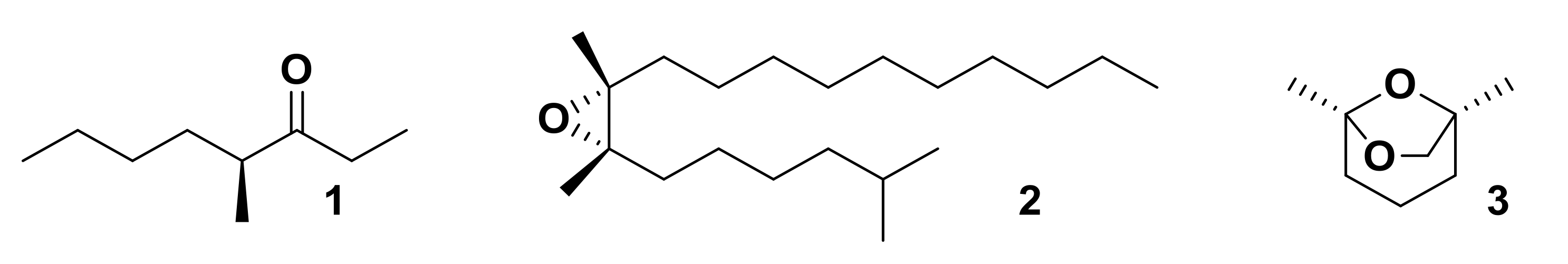
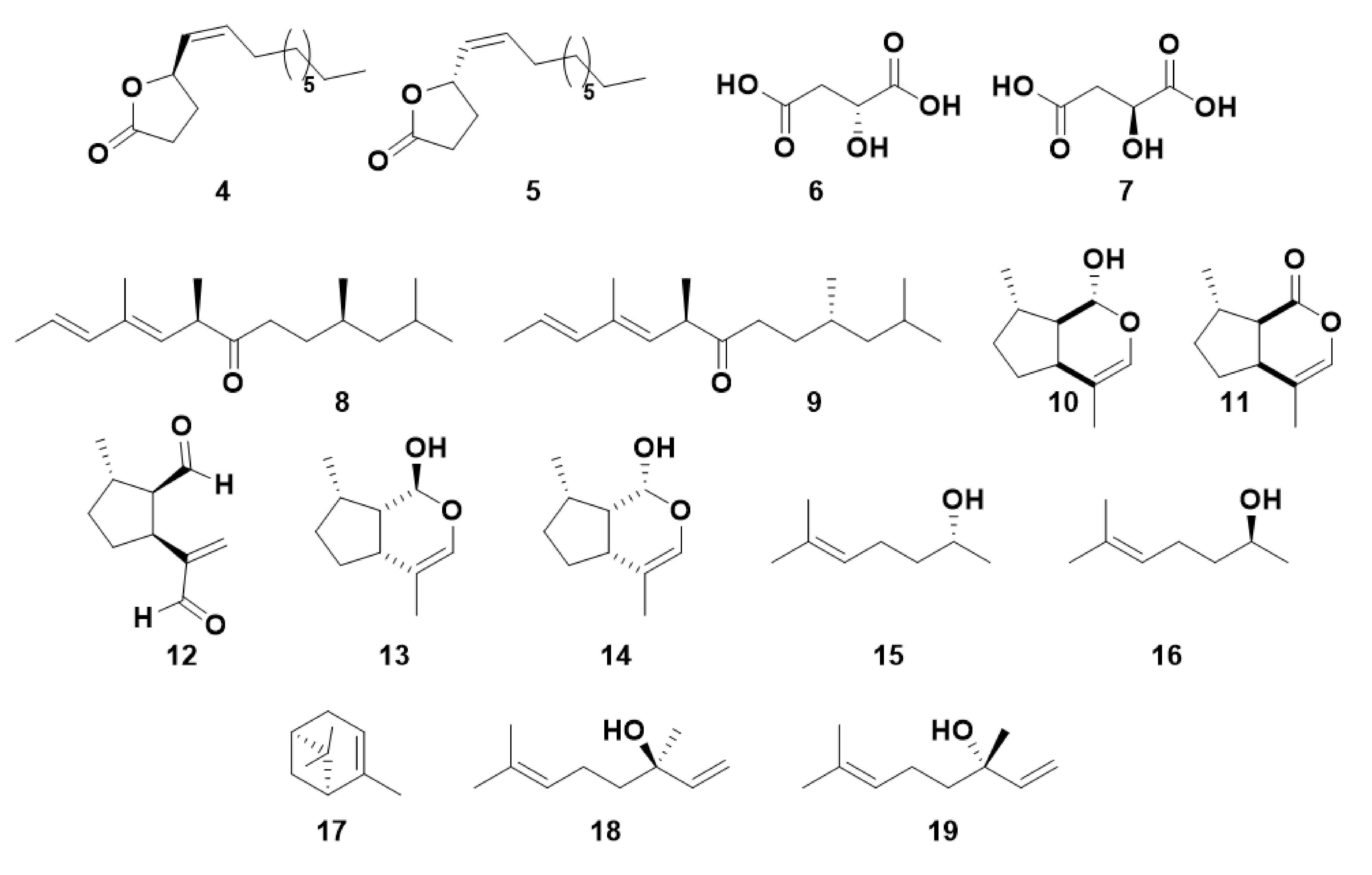
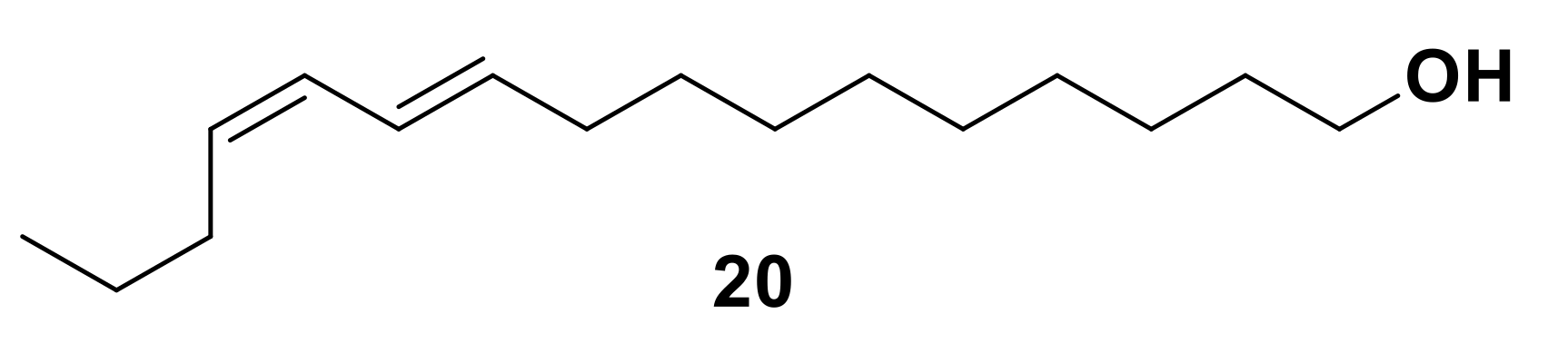
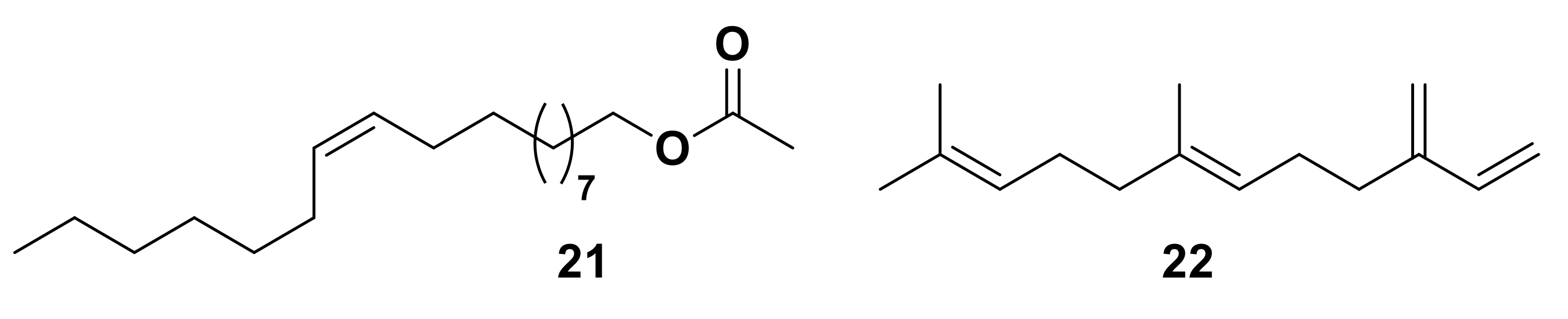
2.3. Other Chiral Semiochemicals
3. Insect Olfactory Proteins
3.1. Introduction to Insect Olfaction
3.2. Odorant-Binding Proteins
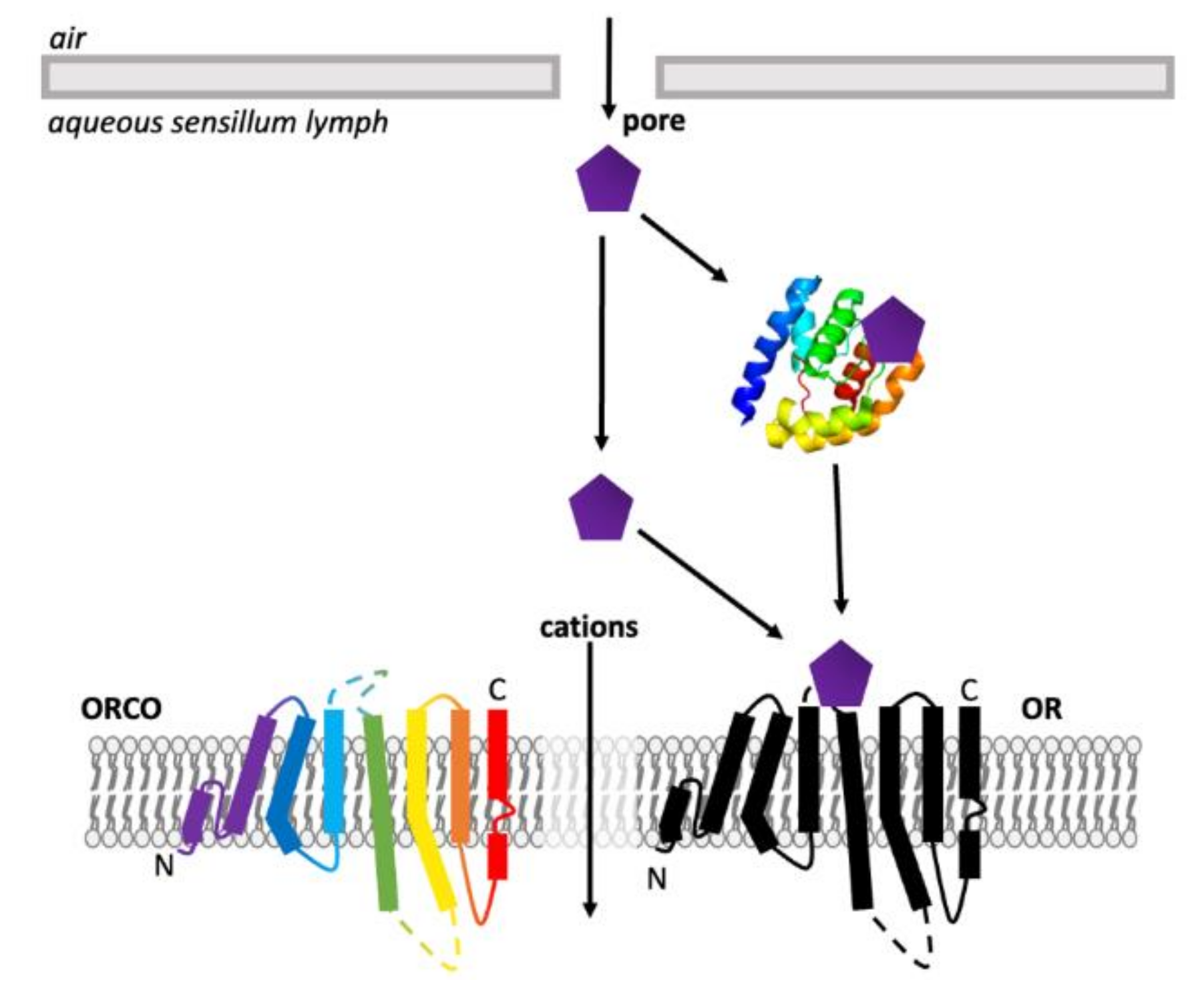
3.3. Odorant Receptors
4. Enantiomeric Discrimination by OBPs
4.1. OBP Specificity
4.2. Enantiomeric Discrimination by Insect OBPs
5. Enantiomeric Discrimination by ORs
5.1. OR Specificity
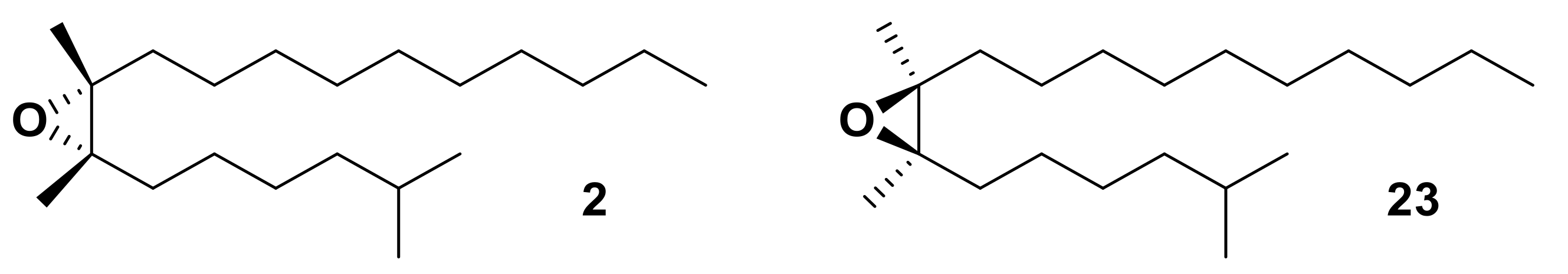
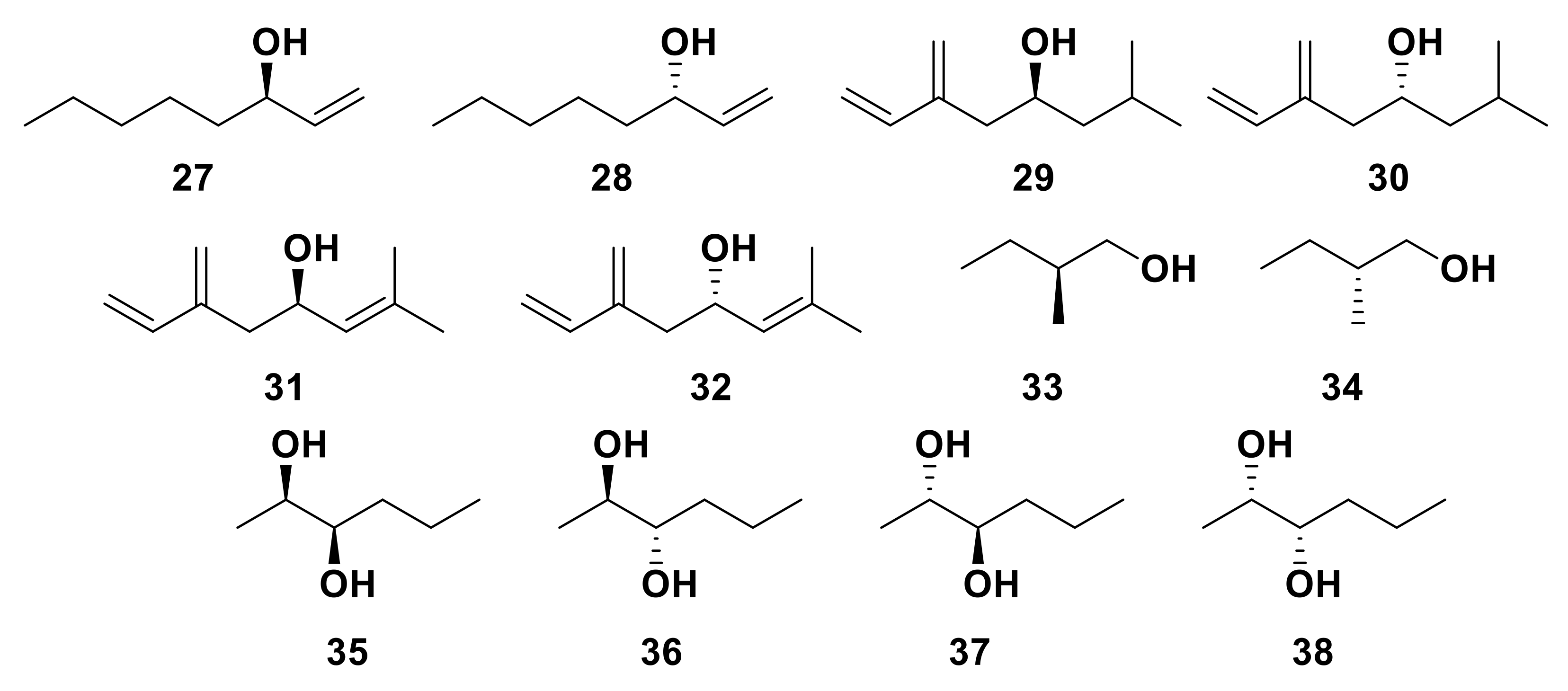
5.2. Enantioselective Insect ORs
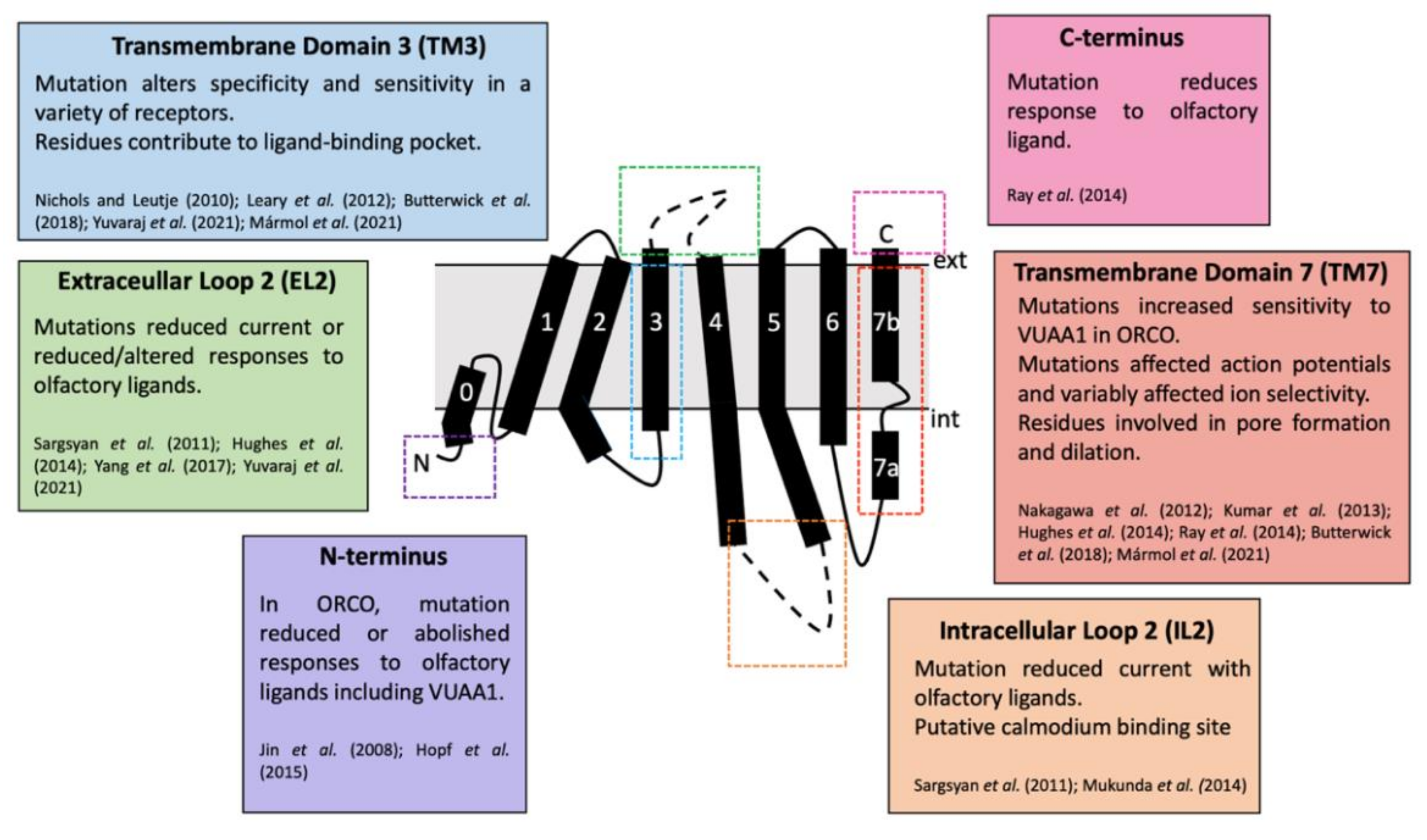
5.3. Structural Studies of Insect ORs
6. Future Research Directions
6.1. Alternative Targets
6.2. Applications of Enantiomeric Discrimination
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Rytz, R.; Croset, V.; Benton, R. Ionotropic Receptors (IRs): Chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 2013, 43, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Reisenman, C.E.; Lei, H.; Guerenstein, P.G. Neuroethology of olfactory-guided behavior and its potential application in the control of harmful insects. Front. Physiol. 2016, 7, 271. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef] [PubMed]
- Touchet, S.; Chamberlain, K.; Woodcock, C.M.; Miller, D.J.; Birkett, M.A.; Pickett, J.A.; Allemann, R.K. Novel olfactory ligands via terpene synthases. Chem. Commun. 2015, 51, 7550–7553. [Google Scholar] [CrossRef]
- Mori, K. Significance of chirality in pheromone science. Bioorganic Med. Chem. 2007, 15, 7505–7523. [Google Scholar] [CrossRef]
- Butterwick, J.A.; del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect olfactory receptors are heteromeric ligand-gated ion channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef]
- Del Mármol, J.; Yedlin, M.A.; Ruta, V. The structural basis of odorant recognition in insect olfactory receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef]
- Terrado, M.; Okon, M.; Mcintosh, L.P.; Plettner, E. Ligand- and pH-Induced Structural Transition of Gypsy Moth Lymantria dispar Pheromone-Binding Protein 1 (LdisPBP1). Biochemistry 2020, 59, 3411–3426. [Google Scholar] [CrossRef]
- Northey, T.; Venthur, H.; De Biasio, F.; Chauviac, F.X.; Cole, A.; Ribeiro, K.A.L.; Grossi, G.; Falabella, P.; Field, L.M.; Keep, N.H.; et al. Crystal structures and binding dynamics of odorant-binding protein 3 from two aphid species Megoura viciae and Nasonovia ribisnigri. Sci. Rep. 2016, 6, 24739. [Google Scholar] [CrossRef] [PubMed]
- Pesenti, M.E.; Spinelli, S.; Bezirard, V.; Briand, L.; Pernollet, J.C.; Tegoni, M.; Cambillau, C. Structural Basis of the Honey Bee PBP Pheromone and pH-induced Conformational Change. J. Mol. Biol. 2008, 380, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Ziemba, B.P.; Murphy, E.J.; Edlin, H.T.; Jones, D.N.M. A novel mechanism of ligand binding and release in the odorant binding protein 20 from the malaria mosquito Anopheles gambiae. Protein Sci. 2013, 22, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Campanacci, V.; Lartigue, A.; Hällberg, B.M.; Jones, T.A.; Giudici-Orticoni, M.T.; Tegoni, M.; Cambillau, C. Moth chemosensory protein exhibits drastic conformational changes and cooperativity on ligand binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5069–5074. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.F.; Briand, L. The 40-year mystery of insect odorant-binding proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Larter, N.K.; Sun, J.S.; Carlson, J.R. Organization and function of Drosophila odorant binding proteins. eLife 2016, 5, e20242. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Birkett, M.A.; Pickett, J.A. Aphid sex pheromones: From discovery to commercial production. Phytochemistry 2003, 62, 651–656. [Google Scholar] [CrossRef]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavoiral, and Biochemical Aspects of Insect Hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Nordlund, D.A.; Lewis, W.J. Terminology of chemical releasing stimuli in intraspecific and interspecific interactions. J. Chem. Ecol. 1976, 2, 211–220. [Google Scholar] [CrossRef]
- Burger, B.V. Mammalian semiochemicals. In The Chemistry of Pheromones and Other Semiochemicals II; Springer: Berlin/Heidelberg, Germany, 2004; pp. 231–278. [Google Scholar]
- Campagna, S.; Mardon, J.; Celerier, A.; Bonadonna, F. Potential semiochemical molecules from birds: A practical and comprehensive compilation of the last 20 years studies. Chem. Senses 2012, 37, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E.; Irving, P.W.; Henderson, P.A. Is there a fish alarm pheromone? A wild study and critique. Proc. R. Soc. B Biol. Sci. 1996, 263, 1551–1556. [Google Scholar] [CrossRef]
- Pickett, J.A.; Allemann, R.K.; Birkett, M.A. The semiochemistry of aphids. Nat. Prod. Rep. 2013, 30, 1277. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.G.; Silverstein, R.M.; Moser, J.C. Biological Responses of Atta texana to Its Alarm Pheromone and the Enantiomer of the Pheromone. Science 1974, 183, 760–762. [Google Scholar] [CrossRef]
- Marumo, S.; Iwaki, S.; Saito, T.; Yamada, M.; Katagiri, K. Synthesis and Activity of Optically Active Disparlure. J. Am. Chem. Soc. 1974, 21, 7842–7844. [Google Scholar]
- Mori, K. Synthesis of optically active forms of frontalin: The pheromone of dendroctonus bark beetles. Tetrahedron 1975, 31, 1381–1384. [Google Scholar] [CrossRef]
- Tumlinson, J.H.; Klein, M.G.; Doolittle, R.E.; Ladd, T.L.; Proveaux, A.T. Identification of the Female Japanese Beetle Sex Pheromone: Inhibition of Male Response by an Enantiomer. Science 1977, 197, 789–792. [Google Scholar] [CrossRef]
- Haniotakis, G.; Francke, W.; Mori, K.; Redlich, H.; Schurig, V. Sex-specific activity of (R)-(-)- and (S)- (+)-1,7-dioxaspiro[5.5]undecane, the major pheromone of Dacus oleae. J. Chem. Ecol. 1986, 12, 1559–1568. [Google Scholar] [CrossRef]
- Shi, X.; Webster, F.X.; Meinwald, J. Synthesis and the determination of the absolute configuration of matsuone, sex pheromone of female Matsucoccus pine scales. Tetrahedron Lett. 1995, 36, 7201–7204. [Google Scholar] [CrossRef]
- Dawson, G.; Griffiths, D.; Janes, N. Identification of an aphid sex pheromone. Nature 1987, 325, 614–616. [Google Scholar] [CrossRef]
- Marsh, D. Sex pheromone in the aphid Megoura viciae. Nature 1972, 238, 31–32. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.A.M.; Dawson, G.W.; Griffiths, D.C.; Pettersson, J.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Sex attractant pheromone of damson-hop aphid Phorodon humuli (Homoptera, aphididae). J. Chem. Ecol. 1990, 16, 3455–3465. [Google Scholar] [CrossRef]
- Dawson, G.W.; Griffiths, D.C.; Merritt, L.A.; Mudd, A.; Pickett, J.A.; Woodcock, C.M. Aphid semiochemicals—A review, and recent advances on the sex pheromone. J. Chem. Ecol. 1990, 16, 3019–3030. [Google Scholar] [CrossRef] [PubMed]
- Borden, J.H.; Hong, C.L.; McLean, J.A.; Slessor, K.N.; Mori, K. Gnathotrichus sulcatus: Synergistic response to enantiomers of the aggregation pheromone sulcatol. Science 1976, 192, 894–896. [Google Scholar] [CrossRef]
- Bello, J.E.; McElfresh, J.S.; Millar, J.G. Isolation and determination of absolute configurations of insect-produced methyl-branched hydrocarbons. Proc. Natl. Acad. Sci. USA 2015, 112, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Szendrei, Z.; Rodriguez-Saona, C. A meta-analysis of insect pest behavioral manipulation with plant volatiles. Entomol. Exp. Appl. 2010, 134, 201–210. [Google Scholar] [CrossRef]
- De Vega, C.; Herrera, C.M.; Dötterl, S. Floral volatiles play a key role in specialized ant pollination. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 32–42. [Google Scholar] [CrossRef]
- Bellesia, F.; Grandi, R.M.; Pagnoni, U.; Pinetti, A.; Trave, R. Biosynthesis of nepetalactone in Nepeta cataria. Phytochemistry 1984, 23, 83–87. [Google Scholar] [CrossRef]
- Lichman, B.R.; Godden, G.T.; Hamilton, J.P.; Palmer, L.; Kamileen, M.O.; Zhao, D.; Vaillancourt, B.; Wood, J.C.; Sun, M.; Kinser, T.J.; et al. The evolutionary origins of the cat attractant nepetalactone in catnip. Sci. Adv. 2020, 6, eaba0721. [Google Scholar] [CrossRef]
- Bakke, A.; Frøyen, P.; Skattebøl, L. Field response to a new pheromonal compound isolated from Ips typographus. Naturwissenschaften 1977, 64, 98–99. [Google Scholar] [CrossRef]
- Erbilgin, N.; Mori, S.R.; Sun, J.H.; Stein, J.D.; Owen, D.R.; Merrill, L.D.; Bolaños, R.C.; Raffa, K.F.; Montiel, T.M.; Wood, D.L.; et al. Response to host volatiles by native and introduced populations of Dendroctonus valens (Coleoptera: Curculionidae, Scolytinae) in North America and China. J. Chem. Ecol. 2007, 33, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Ulland, S.; Ian, E.; Borg-Karlson, A.K.; Mustaparta, H. Discrimination between enantiomers of linalool by olfactory receptor neurons in the cabbage moth Mamestra brassicae (L.). Chem. Senses 2006, 31, 325–334. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Huff, R.M.; Jason Pitts, R. An odorant receptor from Anopheles gambiae that demonstrates enantioselectivity to the plant volatile, linalool. PLoS ONE 2019, 14, e0225637. [Google Scholar] [CrossRef]
- Schneider, D. 100 years of pheromone research—An essay on lepidoptera. Naturwissenschaften 1992, 79, 241–250. [Google Scholar] [CrossRef]
- Hansson, B.S. Olfaction in Lepidoptera. Experientia 1995, 51, 1003–1027. [Google Scholar] [CrossRef]
- Von Butenandt, A.; Beckmann, R.; Stamm, D.; Hecker, E. Über den Sexual-Lockstoff des Seidenspinners Bombyx mori. Reindarstellung Konstitution. Z. Nat. B 1959, 14, 283–284. [Google Scholar]
- Dawson, G.W.; Pickett, J.A.; Smiley, D.W.M. The aphid sex pheromone cyclopentanoids: Synthesis in the elucidation of structure and biosynthetic pathways. Bioorgan. Med. Chem. 1996, 4, 351–361. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A novel family of divergent seven-transmembrane proteins: Candidate odorant receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef]
- Dobritsa, A.A.; Van Der Goes Van Naters, W.; Warr, C.G.; Steinbrecht, R.A.; Carlson, J.R. Integrating the molecular and cellular basis of odor coding in the Drosophila antenna. Neuron 2003, 37, 827–841. [Google Scholar] [CrossRef]
- De Biasio, F.; Riviello, L.; Bruno, D.; Grimaldi, A.; Congiu, T.; Sun, Y.F.; Falabella, P. Expression pattern analysis of odorant-binding proteins in the pea aphid Acyrthosiphon pisum. Insect Sci. 2015, 22, 220–234. [Google Scholar] [CrossRef] [PubMed]
- Bruno, D.; Grossi, G.; Salvia, R.; Scala, A.; Farina, D.; Grimaldi, A.; Zhou, J.J.; Bufo, A.; Vogel, H.; Grosse-Wilde, E.; et al. Sensilla Morphology and Complex Expression Pattern of Odorant Binding Proteins in the Vetch Aphid Megoura viciae (Hemiptera: Aphididae). Front. Phys. 2018, 9, 777. [Google Scholar] [CrossRef]
- Yuvaraj, J.; Andersson, M.; Anderbant, O.; Löfstedt, C. Diversity of olfactory structures: A comparative study of antennal sensilla in Trichoptera and Lepidoptera. Micron 2018, 111, 9–18. [Google Scholar] [CrossRef]
- Gomez-Diaz, C.; Reina, J.H.; Cambillau, C.; Benton, R. Ligands for Pheromone-Sensing Neurons Are Not Conformationally Activated Odorant Binding Proteins. PLoS Biol. 2013, 11, e1001546. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Kristoffersen, L.; Hansson, B.S.; Anderbrant, O.; Larsson, M.C. Aglomerular hemipteran antennal lobes—Basic neuroanatomy of a small nose. Chem. Senses 2008, 33, 771–778. [Google Scholar] [CrossRef]
- Benton, R. On the ORigin of smell: Odorant receptors in insects. Cell. Mol. Life Sci. 2006, 63, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.; Celniker, S.; Holt, R.; Evans, C.; Gocyane, J.; Amanatides, P.; Scherer, S.; Li, P.; Hoskins, R. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Guo, Z.; Liu, Y.; Zhang, Y. Progress and prospects of CRISPR/Cas systems in insects and other arthropods. Front. Physiol. 2017, 8, 608. [Google Scholar] [CrossRef]
- Pelosi, P.; Maida, R. Odorant-binding proteins in insects. Comp. Biochem. Physiol. 1995, 111, 503–514. [Google Scholar] [CrossRef]
- Zhou, J.J.; Vieira, F.G.; He, X.L.; Smadja, C.; Liu, R.; Rozas, J.; Field, L.M. Genome annotation and comparative analyses of the odorant-binding proteins and chemosensory proteins in the pea aphid Acyrthosiphon pisum. Insect Mol. Biol. 2010, 19, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Mosbah, A.; Campanacci, V.; Lartigue, A.; Tegoni, M.; Cambillau, C.; Darbon, H. Solution structure of a chemosensory protein from the moth Mamestra brassicae. Biochem. J. 2003, 369, 39–44. [Google Scholar] [CrossRef]
- Leal, W.S.; Chen, A.M.; Ishida, Y.; Chiang, V.P.; Erickson, M.L.; Morgan, T.I.; Tsuruda, J.M. Kinetics and molecular properties of pheromone binding and release. Proc. Natl. Acad. Sci. USA 2005, 102, 5386–5391. [Google Scholar] [CrossRef] [PubMed]
- Kowcun, A.; Honson, N.; Plettner, E. Olfaction in the gypsy moth, Lymantria dispar: Effect of pH, ionic strength, and reductants on pheromone transport by pheromone-binding proteins. J. Biol. Chem. 2001, 276, 44770–44776. [Google Scholar] [CrossRef] [PubMed]
- Leite, N.R.; Krogh, R.; Xu, W.; Ishida, Y.; Iulek, J.; Leal, W.S.; Oliva, G. Structure of an Odorant-Binding Protein from the Mosquito Aedes aegypti Suggests a Binding Pocket Covered by a pH-Sensitive “Lid”. PLoS ONE 2009, 4, e8006. [Google Scholar] [CrossRef] [PubMed]
- Zubkov, S.; Gronenborn, A.M.; Byeon, I.J.L.; Mohanty, S. Structural consequences of the pH-induced conformational switch in A. polyphemus pheromone-binding protein: Mechanisms of ligand release. J. Mol. Biol. 2005, 354, 1081–1090. [Google Scholar] [CrossRef]
- Wogulis, M.; Morgan, T.; Ishida, Y.; Leal, W.S.; Wilson, D.K. The crystal structure of an odorant binding protein from Anopheles gambiae: Evidence for a common ligand release mechanism. Biochem. Biophys. Res. Commun. 2006, 339, 157–164. [Google Scholar] [CrossRef]
- Han, L.; Zhang, Y.J.; Zhang, L.; Cui, X.; Yu, J.; Zhang, Z.; Liu, M.S. Operating mechanism and molecular dynamics of pheromone-binding protein ASP1 as influenced by pH. PLoS ONE 2014, 9, e110565. [Google Scholar] [CrossRef]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.; Francesca, R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Tuccori, E.; He, X.; Gazzano, A.; Field, L.; Zhou, J.J.; Pelosi, P. Discrimination of alarm pheromone (E)-β-farnesene by aphid odorant-binding proteins. Insect Biochem. Mol. Biol. 2009, 39, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, P.; He, X.L.; Woodcock, C.; Pickett, J.A.; Field, L.M.; Birkett, M.A.; Kalinova, B.; Gomulski, L.M.; Scolari, F.; Gasperi, G.; et al. Identification of pheromone components and their binding affinity to the odorant binding protein CcapOBP83a-2 of the Mediterranean fruit fly, Ceratitis capitata. Insect Biochem. Mol. Biol. 2014, 48, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori Odorant-binding Proteins Reveals that a General Odorant-binding Protein Discriminates Between Sex Pheromone Components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Pelosi, P.; Liu, Y.; Lin, K.J.; Yuan, H.B.; Wang, G.R. Ligand-binding properties of three odorant-binding proteins of the diamondback moth Plutella xylostella. J. Integr. Agric. 2016, 15, 580–590. [Google Scholar] [CrossRef]
- Leal, G.M.; Leal, W.S. Binding of a fluorescence reporter and a ligand to an odorant-binding protein of the yellow fever mosquito, Aedes aegypti. F1000Research 2015, 3, 305. [Google Scholar] [CrossRef]
- Zhang, T.T.; Mei, X.D.; Feng, J.N.; Berg, B.G.; Zhang, Y.J.; Guo, Y.Y. Characterization of three pheromone-binding proteins (PBPs) of Helicoverpa armigera (Hübner) and their binding properties. J. Insect Physiol. 2012, 58, 941–948. [Google Scholar] [CrossRef]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of Pheromone-Sensitive Neurons Is Mediated by Conformational Activation of Pheromone-Binding Protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef]
- Kim, M.S.; Repp, A.; Smith, D.P. LUSH odorant-binding protein mediates chemosensory responses to alcohols in Drosophila melanogaster. Genetics 1998, 150, 711–721. [Google Scholar] [CrossRef]
- Hooper, A.M.; Dufour, S.; He, X.; Muck, A.; Zhou, J.-J.; Almeida, R.; Field, L.M.; Svatos, A.; Pickett, J. High-throughput ESI-MS analysis of binding between the Bombyx mori pheromone-binding protein BmorPBP1, its pheromone components and some analogues. Chem. Commun. 2009, 38, 5725–5727. [Google Scholar] [CrossRef]
- Oldham, N.J.; Krieger, J.; Breer, H.; Fischedick, A.; Hoskovec, M.; Svatos, A. Analysis of the Silkworm Moth Pheromone. Angew. Chemie Int. Ed. 2000, 39, 4341–4343. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, B.; Grossi, G.; Falabella, P.; Liu, Y.; Yan, S.; Lu, J.; Xi, J.; Wang, G. Molecular Basis of Alarm Pheromone Detection in Aphids. Curr. Biol. 2017, 27, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, J.-T.; Zhang, Y.-J.; Chen, J.-L.; Li, X.-C.; Liang, P.; Gao, X.-W.; Zhou, J.-J.; Gu, S.-H. Coordinative mediation of the response to alarm pheromones by three odorant binding proteins in the green peach aphid Myzus persicae. Insect Biochem. Mol. Biol. 2021, 130, 103528. [Google Scholar] [CrossRef]
- Dong, X.-T.; Liao, H.; Zhu, G.-H.; Khuhro, S.A.; Ye, Z.-F.; Yan, Q.; Dong, S.-L. CRISPR/Cas9 mediated PBP1 and PBP3 mutagenesis induced significant reduction in electrophysiological response to sex pheromones in male Chilo suppressalis. Insect Sci. 2017, 26, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Liu, Y.; Yang, T.; Pelosi, P.; Dong, S.; Wang, G. Pheromone binding proteins enhance the sensitivity of olfactory receptors to sex pheromones in Chilo suppressalis. Sci. Rep. 2015, 5, 13093. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.H.; Xu, J.; Cui, Z.; Dong, X.T.; Ye, Z.F.; Niu, D.J.; Huang, Y.P.; Dong, S.L. Functional characterization of SlitPBP3 in Spodoptera litura by CRISPR/Cas9 mediated genome editing. Insect Biochem. Mol. Biol. 2016, 75, 1–9. [Google Scholar] [CrossRef]
- Matsuo, T.; Sugaya, S.; Yasukawa, J.; Aigaki, T.; Fuyama, Y. Odorant-binding proteins OBP57d and OBP57e affect taste perception and host-plant preference in Drosophila sechellia. PLoS Biol. 2007, 5, e118. [Google Scholar] [CrossRef]
- Dani, F.R.; Iovinella, I.; Felicioli, A.; Niccolini, A.; Calvello, M.A.; Carucci, M.G.; Qiao, H.; Pieraccini, G.; Turillazzi, S.; Moneti, G.; et al. Mapping the expression of soluble olfactory proteins in the honeybee. J. Proteome Res. 2010, 9, 1822–1833. [Google Scholar] [CrossRef]
- Wang, S.; Minter, M.; Homem, R.A.; Michaelson, L.V.; Venthur, H.; Lim, K.S.; Withers, A.; Xi, J.; Jones, C.M.; Zhou, J.-J. Odorant binding proteins promote flight activity in the migratory insect, Helicoverpa armigera. Mol. Ecol. 2020, 29, 3795–3808. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Zhu, X.Y.; Fang, L.P.; He, P.; Wang, Z.Q.; Chen, G.; Sun, L.; Ye, Z.F.; Deng, D.G.; Li, J.B. Identification and expression profiles of sex pheromone biosynthesis and transport related genes in Spodoptera litura. PLoS ONE 2015, 10, e0120270. [Google Scholar] [CrossRef]
- Benton, R. Sensitivity and specificity in Drosophila pheromone perception. Trends Neurosci. 2007, 30, 512–519. [Google Scholar] [CrossRef]
- Ban, L.; Napolitano, E.; Serra, A.; Zhou, X.; Iovinella, I.; Pelosi, P. Identification of pheromone-like compounds in male reproductive organs of the oriental locust Locusta migratoria. Biochem. Biophys. Res. Commun. 2013, 437, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Forêt, S.; Wanner, K.W.; Maleszka, R. Chemosensory proteins in the honey bee: Insights from the annotated genome, comparative analyses and expressional profiling. Insect Biochem. Mol. Biol. 2007, 37, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Spehr, M.; Munger, S.D. Olfactory receptors: GPCRs and beyond. J. Neurochem. 2009, 109, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef]
- Wicher, D.; Schafer, R.; Bauernfeind, R.; Stensmyr, M.C.; Heller, R.; Heinemann, S.H.; Hansson, B.S. Drosophila odorant receptors are both ligand-gated and cyclic-nucleotide-activated cation channels. Nature 2008, 452, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef]
- Brand, P.; Robertson, H.M.; Lin, W.; Pothula, R.; Klingeman, W.E.; Jurat-Fuentes, J.L.; Johnson, B.R. The origin of the odorant receptor gene family in insects. eLife 2018, 7, e38340. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Chen, D.; Yang, P.; Jiang, F.; Wang, X.; Kang, L. CRISPR/Cas9 in locusts: Successful establishment of an olfactory deficiency line by targeting the mutagenesis of an odorant receptor co-receptor (Orco). Insect Biochem. Mol. Biol. 2016, 79, 27–35. [Google Scholar] [CrossRef]
- Fandino, R.A.; Haverkamp, A.; Bisch-Knaden, S.; Zhang, J.; Bucks, S.; Nguyen, T.A.T.; Schröder, K.; Werckenthin, A.; Rybak, J.; Stengl, M.; et al. Mutagenesis of odorant coreceptor Orco fully disrupts foraging but not oviposition behaviors in the hawkmoth Manduca sexta. Proc. Natl. Acad. Sci. USA 2019, 116, 15677–15685. [Google Scholar] [CrossRef]
- Trible, W.; Olivos-Cisneros, L.; McKenzie, S.K.; Saragosti, J.; Chang, N.C.; Matthews, B.J.; Oxley, P.R.; Kronauer, D.J.C. Orco Mutagenesis Causes Loss of Antennal Lobe Glomeruli and Impaired Social Behavior in Ants. Cell 2017, 170, 727–735. [Google Scholar] [CrossRef]
- Koutroumpa, F.; Monsempès, C.; Anton, S.; François, M.C.; Montagné, N.; Jacquin-Joly, E. Pheromone Receptor Knock-Out Affects Pheromone Detection and Brain Structure in a Moth. Biomolecules 2022, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Batra, S.; Corcoran, J.; Zhang, D.D.; Pal, P.; Umesh, K.P.; Kulkarni, R.; Löfstedt, C.; Sowdhamini, R.; Olsson, S.B. A functional agonist of insect olfactory receptors: Behavior, physiology and structure. Front. Cell. Neurosci. 2019, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Yuvaraj, J.; Roberts, R.; Sonntag, Y.; Hou, X.; Grosse-Wilde, E.; Machara, A.; Hansson, B.; Johanson, U.; Löfstedt, C.; Andersson, M. Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. BMC Biol. 2021, 19, 16. [Google Scholar] [CrossRef] [PubMed]
- Radadiya, A.; Pickett, J.A. Characterizing human odorant signals: Insights from insect semiochemistry and in silico modelling. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190263. [Google Scholar] [CrossRef]
- Field, L.M.; Pickett, J.A.; Wadhams, L.J. Molecular studies in insect olfaction. Insect Mol. Biol. 2000, 9, 545–551. [Google Scholar] [CrossRef]
- Zaremska, V.; Tan, J.; Lim, S.; Knoll, W.; Pelosi, P. Isoleucine Residues Determine Chiral Discrimination of Odorant-Binding Protein. Chem. Eur. J. 2020, 26, 8720–8724. [Google Scholar] [CrossRef]
- Damberger, F.; Horst, R.; Wüthrich, K.; Peng, G.; Nikonova, L.; Leal, W.S. NMR characterization of a pH-dependent equilibrium between two folded solution conformations of the pheromone-binding protein from Bombyx mori. Protein Sci. 2000, 9, 1038–1041. [Google Scholar] [CrossRef]
- Yu, Y.; Plettner, E. Enantiomer and conformer recognition of (+) and (−)-disparlure and their analogs by the pheromone binding proteins of the gypsy moth, Lymantria dispar. Bioorgan. Med. Chem. 2013, 21, 1811–1822. [Google Scholar] [CrossRef]
- Wojtasek, H.; Hansson, B.S.; Leal, W.S. Attracted or repelled?—A matter of two neurons, one pheromone binding protein, and a chiral center. Biochem. Biophys. Res. Commun. 1998, 250, 217–222. [Google Scholar] [CrossRef]
- Plettner, E.; Lazar, J.; Prestwich, E.G.; Prestwich, G.D. Discrimination of pheromone enantiomers by two pheromone binding proteins from the gypsy moth Lymantria dispar. Biochemistry 2000, 39, 8953–8962. [Google Scholar] [CrossRef]
- Drakou, C.E.; Tsitsanou, K.E.; Potamitis, C.; Fessas, D.; Zervou, M.; Zographos, S.E. The crystal structure of the AgamOBP1•Icaridin complex reveals alternative binding modes and stereo-selective repellent recognition. Cell. Mol. Life Sci. 2016, 74, 319–338. [Google Scholar] [CrossRef] [PubMed]
- Lartigue, A.; Gruez, A.; Spinelli, S.; Rivière, S.; Brossut, R.; Tegoni, M.; Cambillau, C. The crystal structure of a cockroach pheromone-binding protein suggests a new ligand binding and release mechanism. J. Biol. Chem. 2003, 278, 30213–30218. [Google Scholar] [CrossRef] [PubMed]
- Geithe, C.; Protze, J.; Kreuchwig, F.; Krause, G.; Krautwurst, D. Structural determinants of a conserved enantiomer-selective carvone binding pocket in the human odorant receptor OR1A1. Cell. Mol. Life Sci. 2017, 74, 4209–4229. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Touhara, K. Enantioselective recognition of menthol by mouse odorant receptors. Biosci. Biotechnol. Biochem. 2015, 79, 1980–1986. [Google Scholar] [CrossRef]
- Bohbot, J.D.; Dickens, J.C. Selectivity of odorant receptors in insects. Front. Cell. Neurosci. 2012, 6, 2010–2013. [Google Scholar] [CrossRef]
- Zhang, D.D.; Löfstedt, C. Moth pheromone receptors: Gene sequences, function, and evolution. Front. Ecol. Evol. 2015, 3, 105. [Google Scholar] [CrossRef]
- Zhang, R.B.; Liu, Y.; Yan, S.C.; Wang, G.R. Identification and functional characterization of an odorant receptor in pea aphid, Acyrthosiphon pisum. Insect Sci. 2019, 26, 58–67. [Google Scholar] [CrossRef]
- Malnic, B.; Hirono, J.; Sato, T.; Buck, L.B. Combinatorial receptor codes for odors. Cell 1999, 96, 713–723. [Google Scholar] [CrossRef]
- Seki, Y.; Dweck, H.K.M.; Rybak, J.; Wicher, D.; Sachse, S.; Hansson, B.S. Olfactory coding from the periphery to higher brain centers in the Drosophila brain. BMC Biol. 2017, 15, 18. [Google Scholar] [CrossRef]
- Pask, G.M.; Slone, J.D.; Millar, J.G.; Das, P.; Moreira, J.A.; Zhou, X.; Bello, J.; Berger, S.L.; Bonasio, R.; Desplan, C.; et al. Specialized odorant receptors in social insects that detect cuticular hydrocarbon cues and candidate pheromones. Nat. Commun. 2017, 8, 297. [Google Scholar] [CrossRef]
- Hill, S.R.; Majeed, S.; Ignell, R. Molecular basis for odorant receptor tuning: A short C-terminal sequence is necessary and sufficient for selectivity of mosquito Or8. Insect Mol. Biol. 2015, 24, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.; Steinbach, N.; Stensmyr, M.C.; Hansson, B.S.; Vosshall, L.B. A natural polymorphism alters odour and DEET sensitivity in an insect odorant receptor. Nature 2011, 478, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.N.; Taylor, R.W.; Pask, G.M.; Zwiebel, L.J.; Newcomb, R.D.; Christie, D.L. A Conserved Aspartic Acid Is Important for Agonist (VUAA1) and Odorant/Tuning Receptor-Dependent Activation of the Insect Odorant Co-Receptor (Orco). PLoS ONE 2013, 8, e70218. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Leal, W.S. Probing insect odorant receptors with their cognate ligands: Insights into structural features. Biochem. Biophys. Res. Commun. 2013, 435, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Nichols, A.S.; Luetje, C.W. Transmembrane segment 3 of Drosophila melanogaster odorant receptor subunit 85b contributes to ligand-receptor interactions. J. Biol. Chem. 2010, 285, 11854–11862. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Pellegrino, M.; Sato, K.; Vosshall, L.B.; Touhara, K. Amino acid residues contributing to function of the heteromeric insect olfactory receptor complex. PLoS ONE 2012, 7, e32372. [Google Scholar] [CrossRef] [PubMed]
- Bohbot, J.D.; Dickens, J.C. Characterization of an enantioselective odorant receptor in the yellow fever mosquito Aedes aegypti. PLoS ONE 2009, 4, e7032. [Google Scholar] [CrossRef]
- Dickens, J.C.; Bohbot, J.D. Mini review: Mode of action of mosquito repellents. Pestic. Biochem. Physiol. 2013, 106, 149–155. [Google Scholar] [CrossRef]
- Dekel, A.; Pitts, R.J.; Yakir, E.; Bohbot, J.D. Evolutionarily conserved odorant receptor function questions ecological context of octenol role in mosquitoes. Sci. Rep. 2016, 6, 37330. [Google Scholar] [CrossRef]
- Bohbot, J.D.; Pitts, R.J.; Kwon, H.-W.; Rutzler, M.; Robertson, H.M.; Zwiebel, L.J. Molecular characterization of the Aedes aegypti odorant receptor gene family. Insect Mol. Biol. 2007, 16, 525–537. [Google Scholar] [CrossRef]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G protein-coupled receptors in Anopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Kythreoti, G.; Sdralia, N.; Tsitoura, P.; Papachristos, D.P.; Michaelakis, A.; Karras, V.; Ruel, D.M.; Yakir, E.; Bohbot, J.D.; Schulz, S.; et al. Volatile allosteric antagonists of mosquito odorant receptors inhibit human-host attraction. J. Biol. Chem. 2021, 296, 100172. [Google Scholar] [CrossRef]
- Chapman, O.L.; Mattes, K.C.; Sheridan, R.S.; Klun, J.A. Stereochemical Evidence of Dual Chemoreceptors for an Achiral Sex Pheromone in Lepidoptera1. J. Am. Chem. Soc. 1978, 100, 4878–4884. [Google Scholar] [CrossRef]
- Yang, K.; Huang, L.Q.; Ning, C.; Wang, C.Z. Two single-point mutations shift the ligand selectivity of a pheromone receptor between two closely related moth species. eLife 2017, 6, e29100. [Google Scholar] [CrossRef]
- Leary, G.P.; Allen, J.E.; Bunger, P.L.; Luginbill, J.B.; Linn, C.E.; Macallister, I.E.; Kavanaugh, M.P.; Wanner, K.W. Single mutation to a sex pheromone receptor provides adaptive specificity between closely related moth species. Proc. Natl. Acad. Sci. USA 2012, 109, 14081–14086. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.T.; Wang, G.; Zwiebel, L.J.; Luetje, C.W. A determinant of odorant specificity is located at the extracellular loop 2-transmembrane domain 4 interface of an anopheles gambiae odorant receptor subunit. Chem. Senses 2014, 39, 761–769. [Google Scholar] [CrossRef]
- Sargsyan, V.; Getahun, M.N.; Llanos, S.L.; Olsson, S.B.; Hansson, B.S.; Wicher, D. Phosphorylation via PKC regulates the function of the Drosophila odorant co-receptor. Front. Cell. Neurosci. 2011, 5, 1–8. [Google Scholar] [CrossRef]
- Hopf, T.A.; Morinaga, S.; Ihara, S.; Touhara, K.; Marks, D.S.; Benton, R. Amino acid coevolution reveals three-dimensional structure and functional domains of insect odorant receptors. Nat. Commun. 2015, 6, 6077. [Google Scholar] [CrossRef]
- Jin, X.; Ha, T.S.; Smith, D.P. SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 10996–11001. [Google Scholar] [CrossRef]
- Ray, A.; van Naters, W.G.; Carlson, J.R. Molecular determinants of odorant receptor function in insects. J. Biosci. 2014, 39, 555–563. [Google Scholar] [CrossRef]
- Turner, R.M.; Derryberry, S.L.; Kumar, B.N.; Brittain, T.; Zwiebel, L.J.; Newcomb, R.D.; Christie, D.L. Mutational analysis of cysteine residues of the insect odorant co-receptor (Orco) from Drosophila melanogaster reveals differential effects on agonist- and odorant-tuning receptor-dependent activation. J. Biol. Chem. 2014, 289, 31837–31845. [Google Scholar] [CrossRef] [PubMed]
- Mukunda, L.; Miazzi, F.; Kaltofen, S.; Hansson, B.S.; Wicher, D. Calmodulin modulates insect odorant receptor function. Cell Calcium 2014, 55, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Antony, B.; Johny, J.; Montagné, N.; Jacquin-Joly, E.; Capoduro, R.; Cali, K.; Persaud, K.; Al-Saleh, M.A.; Pain, A. Pheromone receptor of the globally invasive quarantine pest of the palm tree, the red palm weevil (Rhynchophorus ferrugineus). Mol. Ecol. 2021, 30, 2025–2039. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Leal, W.S. Chiral discrimination of the Japanese beetle sex pheromone and a behavioral antagonist by a pheromone-degrading enzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 9076–9080. [Google Scholar] [CrossRef]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: Ionotropic receptors and odorant receptors. Cell Tissue Res. 2021, 383, 7–19. [Google Scholar] [CrossRef]
- Croset, V.; Schleyer, M.; Arguello, J.R.; Gerber, B.; Benton, R. A molecular and neuronal basis for amino acid sensing in the Drosophila larva. Sci. Rep. 2016, 6, 34871. [Google Scholar] [CrossRef]
- Pelosi, P.; Mastrogiacomo, R.; Iovinella, I.; Tuccori, E.; Persaud, K.C. Structure and biotechnological applications of odorant-binding proteins. Appl. Microbiol. Biotechnol. 2014, 98, 61–70. [Google Scholar] [CrossRef]
- Wang, S.; Duan, H.; Yang, X.; Sun, Y.F.; Du, S.; Qin, Y.; Duan, H.; Yang, X. Computer-aided rational design of novel EBF analogues with an aromatic ring. J. Mol. Model. 2016, 22, 144. [Google Scholar] [CrossRef]
- Scorsone, E.; Manai, R.; Cali, K.; Ricatti, M.J.; Farno, S.; Persaud, K.; Mucignat, C. Biosensor array based on ligand binding proteins for narcotics and explosives detection. Sensors Actuators B Chem. 2021, 334, 129587. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | Receptor | Ligands | Reference |
|---|---|---|---|---|
| Aedes aegypti | Yellow Fever Mosquito | AaegOR8 | (R)-1-Octen-3-ol (S)-1-Octen-3-ol | Bohbot and Dickens, 2009 [128] |
| Culex quinquefasciatus | Southern House Mosquito | CquiOR8 | Hill et al., 2015 [122] | |
| Toxorhynchites amboinensis | Elephant Mosquito | TambOR8 | Dekel et al., 2016 [130] | |
| Anopheles gambiae | Malaria Vector Mosquito | AgamOR29 | (R)-Linalool (S)-Linalool | Huff and Pitts, 2019 [45] |
| Megacyllene caryae | Hickory Borer | McarOR20 | (2S,3R)-2,3-Hexanediol (2R,3S)-2,3-Hexanediol (2S,3S)-2,3-Hexanediol | Mitchell et al., 2013 [133] |
| Ips typographus | Eurasian spruce Bark Beetle | ItypOR46 | (R)-Ipsenol (S)-Ipsenol | Yuvaraj et al., 2021 [104] |
| ItypOR49 | (R)-Ipsendiol (S)-Ipsendiol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sims, C.; Birkett, M.A.; Withall, D.M. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects 2022, 13, 368. https://doi.org/10.3390/insects13040368
Sims C, Birkett MA, Withall DM. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects. 2022; 13(4):368. https://doi.org/10.3390/insects13040368
Chicago/Turabian StyleSims, Cassie, Michael A. Birkett, and David M. Withall. 2022. "Enantiomeric Discrimination in Insects: The Role of OBPs and ORs" Insects 13, no. 4: 368. https://doi.org/10.3390/insects13040368
APA StyleSims, C., Birkett, M. A., & Withall, D. M. (2022). Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects, 13(4), 368. https://doi.org/10.3390/insects13040368