
Potential of Antimicrobial Peptide-Overexpressed Tenebrio molitor Larvae Extract as a Natural Preservative for Korean Traditional Sauces


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tenebrio molitor Larvae, Fungal, and Bacterial Strains
2.2. Preparation of TML Extract Containing Overexpressed AMPs Induced by Bacterial Challenge
2.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) for the Analysis of AMP Expression in iTML
2.4. Radial Diffusion Assay (RDA) for the Antibacterial and Antifungal Activities of iTME
2.5. Determination of the Minimum Inhibitory Concentration (MIC) of iTME against Food Poisoning Bacteria
2.6. Stability of iTME
2.6.1. Stability of iTME under High Temperature
2.6.2. Stability of iTME under Various pH
2.6.3. Stability of iTME under Various Salinity
2.7. Analysis of the Antibacterial Activity of iTME under Human Proteolytic Enzymes
2.8. Evaluation of the Preservative Potential of iTME for Ganjang
2.9. Statistical Analysis
3. Results
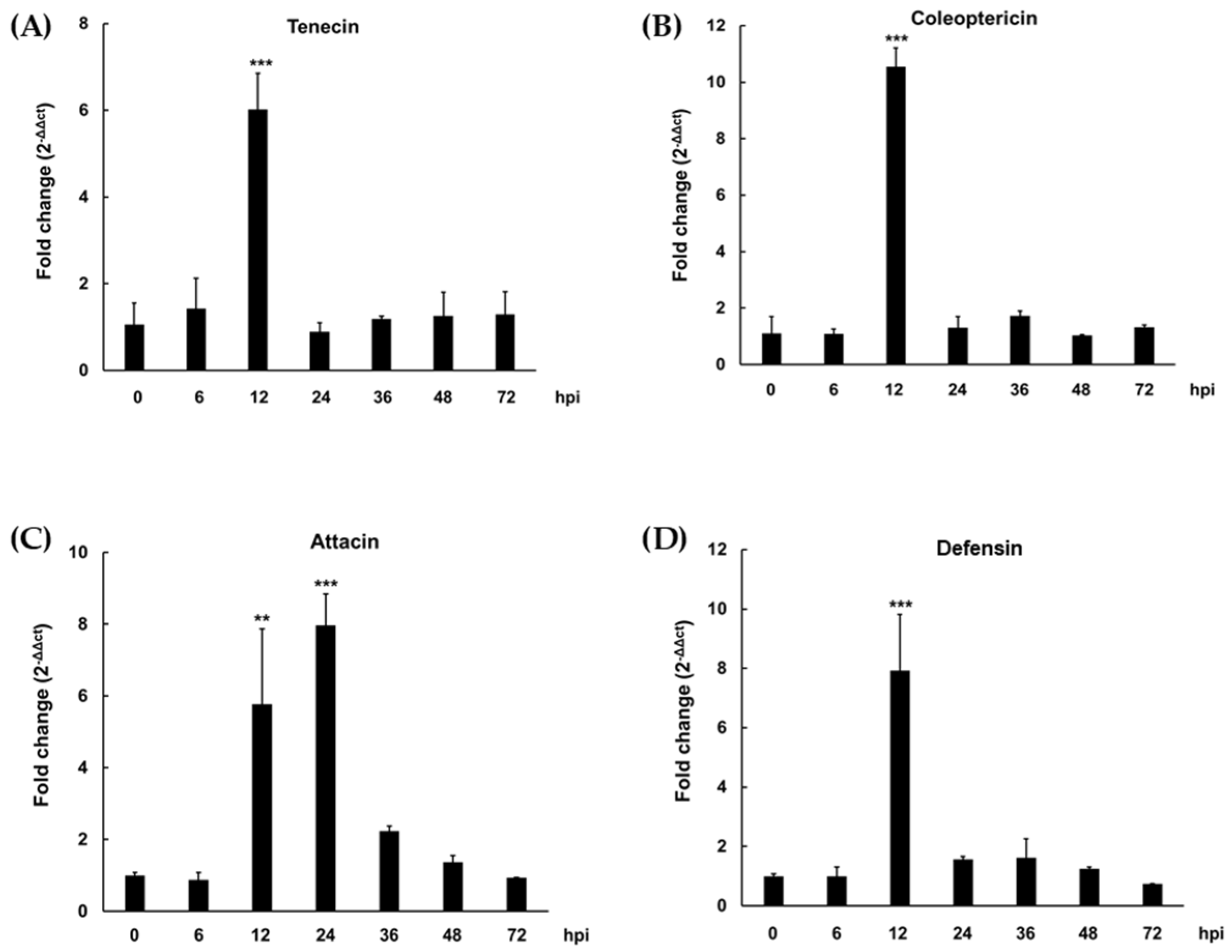
3.1. Determination of the Timing of Maximal Expression of AMP in TML Injected with L. plantarum
3.2. Inhibition Activity of iTME against Various Microorganisms
3.3. MIC of iTME against Food Poisoning Bacteria
3.4. Stability of the Antimicrobial Activity of iTME under Various Environments
3.4.1. High Temperature
3.4.2. pH
3.4.3. Salinity
3.5. Inactivation of the Antibacterial Activity of iTME under Human Proteolytic Enzymes
3.6. Analysis of the Antimicrobial Activity of iTME in Korean Traditional Soy Sauce Inoculated with Harmful Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kang, S.H.; Lee, S.; Ko, J.M.; Hwang, I.K. Comparisons of the physicochemical characteristics of Korean traditional soy sauce with varying soybean seeding periods and regions of production. Korean J. Food Nutr. 2011, 24, 761–769. [Google Scholar] [CrossRef]
- Bechman, A.; Phillips, R.D.; Chen, J. Changes in selected physical property and enzyme activity of rice and barley koji during fermentation and storage. J. Food Sci. 2012, 77, M318–M322. [Google Scholar] [CrossRef] [PubMed]
- Nout, M.; Ruikes, M.; Bouwmeester, H.; Beljaars, P. Effect of processing conditions on the formation of biogenic amines and ethyl carbamate in soybean tempe. J. Food Saf. 1993, 13, 293–303. [Google Scholar] [CrossRef]
- Sserumaga, J.P.; Ortega-Beltran, A.; Wagacha, J.M.; Metegu, C.K.; Bandyopadhyay, R. Aflatoxin-producing fungi associated with pre-harvest maize contamination in Uganda. Int. J. Food Microbiol. 2020, 313, 108376. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.K.; Kang, S.M.; Noh, Y.S. Quality properties on soy bean pastes made with microorganisms isolated fromn traditional soy bean pastes. Korean J. Food Sci. Technol. 2000, 32, 1266–1270. [Google Scholar]
- Ibrahim, S.A.; Yang, H.; Seo, C.W. Antimicrobial activity of lactic acid and copper growth of Salmonella and Ecsherichia coli O157:H7 in laboratory medium and carrot juice. Food Chem. 2008, 109, 137–143. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Madrazo, A.L.; Campos, M.R.S. Review of antimicrobial peptides as promoters of food safety: Limitations and posibilities within the food industry. J. Food Saf. 2020, 40, e12854. [Google Scholar] [CrossRef]
- Wu, Q.; Patočka, J.; Kuča, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Shin, E. Antimicrobials and antimicrobial resistant superbacteria. Ewha Med. J. 2017, 40, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Mills, S.; Stanton, C.; Hill, C.; Ross, R.P. New development and applications of bactericins and peptides in food. Ann. Rev. Food Sci. Technol. 2011, 2, 299–329. [Google Scholar] [CrossRef] [PubMed]
- Upendra, R.S.; Khandelwal, P.; Jana, K.; Ajay Kumar, N.; Gayathri Devi, M.; Stephaney, M.L. Bactericin production from indigenous strains of lactic acid bacteria isolated from selected fermented food sources. Int. J. Pharm. Res. Health Sci. 2016, 4, 982–990. [Google Scholar]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial peptides: A new hope in biomedical and pharmaceutical fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632. [Google Scholar] [CrossRef]
- Mohammad, R.; Vilcinskas, A. Short antimicrobial peptides as cosmetic ingredients to deter dermatological pathogens. Appl. Microbiol. Biotechnol. 2015, 99, 8847–8855. [Google Scholar] [CrossRef] [Green Version]
- Galvez, A.M.; Grande Burges, M.J.; Lucas Loper, R.; Perez Pulido, R. Natural antimicrobials for food presevation. In Food Biopreservation; Galvez, A., GrandeBurgos, M.J., LucasLopez, R., Perez Pulido, R., Eds.; Springer: New York, NY, USA, 2014; pp. 1–14. [Google Scholar] [CrossRef]
- Bedoya, R.U.; Buchhop, J.; Joy, J.B.; Durvasula, R.; Lowenberger, C. Prolixicin: A novel antimicrobial peptide isolated from Rhodnius prolixus with differential activity against bacteria and Trypanosoma cruzi. Insect Mol. Biol. 2011, 20, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Leung, K.W.; Thakur, N.N.; Tan, A.; Jack, R.W. Distinguishing between different pathways of bilayer disruption by the related antimicrobial peptides cecropin B, B1 and B3. Eur. J. Biochem. 2003, 270, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Sommam, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study of antimicrobial peptides identified in the Black Soldier Fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 16875. [Google Scholar] [CrossRef]
- Shelomi, M.; Jacobs, C.; Vilcinskas, A.; Vogel, H. The unique antimicrobial peptide repertoire of stick insects. Dev. Comp. Immunol. 2020, 103, 103471. [Google Scholar] [CrossRef]
- Manniello, M.D.; Moretta, A.; Salvia, R.; Scieuzo, C.; Lucchetti, D.; Vogel, H.; Sgambato, A.; Falabella, P. Insect antimicrobial peptides: Potential weapons to counteract the antibiotic resistance. Cell. Mol. Life. Sci. 2021, 78, 4259–4282. [Google Scholar] [CrossRef]
- Di Somma, A.; Moretta, A.; Cané, C.; Scieuzo, C.; Salvia, R.; Falabella, P.; Duilio, A. Structural and functional characterization of a novel recombinant antimicrobial peptide from Hermetia illucens. Curr. Issues Mol. Biol. 2022, 44, 1. [Google Scholar] [CrossRef]
- Mitsuhara, I.; Matsufuru, H.; Ohshima, M.; Kaku, H.; Nakajima, Y.; Murai, N.; Natori, S.; Ohashi, Y. Induced expression of sarcotoxin IA enhanced host resistance against both bacterial and fungal pathogens in transgenic tobacco. Mol. Plant-Microbe Interact. 2020, 13, 860–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coca, M.; Peñas, G.; Gómez, J.; Campo, S.; Bortolotti, C.; Messeguer, J.; Segundo, B.S. Enhanced resistance to the rice blast fungus Magnaporthe grisea conferred by expression of a cecropin A gene in transgenic rice. Planta 2006, 223, 392–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, Y.N.; Jan, L.Y. Branching out: Mechanisms of dendritic arborization. Nat. Rev. Neurosci. 2010, 11, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sharma, R.; Imamura, M.; Yamakawa, M.; Machii, H. Transgenic expression of cecropin B, an antibacterial peptide from Bombyx moro, confers enhanced resistance to bacterial leaf blight in rice. FEBS Lett. 2000, 484, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Hwang, D.; Lim, C.-H.; Lee, S.H.; Goo, T.-W.; Yun, E.-Y. Effect of feed containing Hermetia illucens larvae immunized by Lactobacillus plantarum injection on the growth and immunity of rainbow trout (Oncorhynchus mykiss). Insects 2021, 12, 801. [Google Scholar] [CrossRef]
- Xie, Z.; Zhao, Q.; Wang, H.; Wen, L.; Li, W.; Zhang, X.; Lin, W.; Xie, Q.; Wang, Y. Effects of antibacterial peptide combinations on growth performance, intestinal health, and immune function of broiler chikens. Poult. Sci. 2020, 99, 6481–6492. [Google Scholar] [CrossRef]
- Sultana, A.; Luo, H.; Ramakrishna, S. Harvesting of antimicrobial peptides from insect (Hermetia illucens) and its applications in the food packaging. Appl. Sci. 2021, 11, 6991. [Google Scholar] [CrossRef]
- Tang, C.; Yang, D.; Liao, H.; Sun, H.; Liu, C.; Wei, L.; Li, F. Edible insects as a food source: A review. Food Prod. Process. Nutr. 2019, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- MFDS. Food Code; Ministry of Food and Drug Safety: Cheongju, Korea, 2021. [Google Scholar]
- Hwang, D.; Lee, W.S.; Goo, T.W.; Park, S.W.; Yun, E.Y. Effect of pretreatment with paste and sauce extract made using Tenebrio molitor larvae on ethanol-damaged HepG2 cells. Entomol. Res. 2019, 49, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Cotty, P.J. Influence of field application of an atoxigenic strain of Aspergillus flavus on the population of A. flavus infecting cotton bolls and on the aflatoxin content of cottonseed. Biol. Control. 1994, 84, 1270–1277. [Google Scholar]
- Amike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, N.; Kocsubé, S.; Vágvölgyi, C.; Varga, J. Current trends in aflatoxin research. Acta Biol. Szeged. 2013, 57, 95–107. [Google Scholar]
- Keshavarz, M.; Jo, Y.H.; Park, K.B.; Ko, H.J.; Edosa, T.T.; Lee, Y.S.; Han, Y.S. TmDorX2 positively regulated antimicrobial peptides in Tenebrio molitor gut, fat body, and hemocytes in response to bacterial and fungal infection. Sci. Rep. 2019, 9, 16878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Z.; Luo, J.; Liu, F.; Zhang, Q.; Jia, S. Effects of pH, temperature, storage time, and protective agents on nisin antibacterial stability. Adv. Appl. Biotechnol. 2015, 333, 305–312. [Google Scholar] [CrossRef]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major food-borne zoonotic bacterial pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Phupiewkham, W.; Sirithorn, P.; Saksirirat, W.; Thammasirirak, S. Study of Biological Activity of Trichoderma harzianum Isolate T9. Khon Kaen Univ. J. (Grad. Stud.) 2010, 10, 9–13. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [Green Version]
- Bulet, P.; Hetru, C.; Dimarcq, J.L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Santiyanont, P.; Chantarasakha, K.; Tepkasikul, P.; Srimaruem, Y.; Mhuantong, W.; Tangphatsornruang, S.; Zo, Y.G.; Chokesajjawatee, N. Dynamics of biogenic amines and bacterial communities in a Thai fermented pork product Nham. Food Res. Int. 2019, 119, 110–118. [Google Scholar] [CrossRef]
- Kim, H.S.; Jung, S.K.; Cho, S.H.; Ku, J.G.; Lee, S.C. Preparation and effect of eudragit E100 microcapsules containing grapefruit seed extract of Kimchi. Korean J. Food Sci. Nutr. 2003, 32, 1239–1244. [Google Scholar]
- Selim, S.; Alfy, S.; Abdel Aziz, M.; Mashait, M.; Warrad, M. Evolution of bactericidal activity of selected food additives against food borne microbial pathogens. Biosci. Biotechnol. Res. Asia 2012, 9, 7–17. [Google Scholar] [CrossRef]
- Cho, S.H.; Kim, C.H.; Par, W.P. Antimicrobial activities of botanical antimicrobial agent-grapefruit seed extract mixture for the preparation of antimicrobial packaging paper. Korean J. Food Preserv. 2004, 11, 411–416. [Google Scholar]
- Torres, J.A.; Bouzas, J.O.; Karel, M. Sorbic acid stability durin processing and storage of an intermediate moisture cheeze analog. J. Food Process. Preserv. 1989, 13, 409–415. [Google Scholar] [CrossRef]
- Hyun, I.K.; Kim, M.Y.; Kim, S.; Lee, J.S.; Choi, A.R.; Kang, S.S. Funtional properties of yogurt fermented by bacteriocin-producing Pediococcus acidilactici. J. Dairy Sci. Biotechnol. 2020, 38, 154–160. [Google Scholar] [CrossRef]
- Han, S.-R.; Yun, E.-Y.; Kim, J.-Y.; Hwang, J.S.; Jeong, E.J.; Moon, K.S. Evaluation of genotoxicity and 28-day oral dose toxicity on freeze-dried powder of Tenebio molitor larvae (yellow mealworm). Toxicol. Res. 2014, 30, 121–130. [Google Scholar] [CrossRef] [PubMed]
- NIFDS. Guidelines for Safety Assessment of New Food Ingredients; National Institute of Food and Drug Safety Evaluation: Cheongju, Korea, 2017. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Ganjang | Gochujang | |
|---|---|---|
| pH | 4.74 ± 0.09 | 5.04 ± 0.07 |
| Salinity (%) | 19.33 ± 1.68 | 6.96 ± 0.30 |
| iTME (mg/mL) | 0 | 0.8 | 0.9 | 1 |
|---|---|---|---|---|
| - | 0.040 ± 0.001 | 0.040 ± 0.002 | 0.041 ± 0.001 | 0.040 ± 0.001 |
| E. coli | 1.159 ± 0.121 | 1.152 ± 0.045 | 0.208 ± 0.014 | 0.044 ± 0.002 |
| B. cereus | 0.862 ± 0.092 | 0.646 ± 0.076 | 0.267 ± 0.015 | 0.042 ± 0.001 |
| S. aureus | 1.095 ± 0.071 | 1.103 ± 0.047 | 0.164 ± 0.005 | 0.041 ± 0.003 |
| E. coli iTME (mg/mL) | − 0 | + 0.8 | + 0.9 | + 1 | ||
|---|---|---|---|---|---|---|
| Heating time | 40 °C (h) | 0 | 0.041 ± 0.001 | 1.131 ± 0.142 | 0.257 ± 0.012 | 0.042 ± 0.001 |
| 36 | 0.040 ± 0.000 | 1.148 ± 0.087 | 0.195 ± 0.014 | 0.051 ± 0.002 | ||
| 48 | 0.042 ± 0.001 | 1.195 ± 0.098 | 0.210 ± 0.010 | 0.043 ± 0.001 | ||
| 72 | 0.041 ± 0.001 | 1.145 ± 0.081 | 0.187 ± 0.009 | 0.041 ± 0.001 | ||
| 100 °C (s) | 5 | 0.040 ± 0.000 | 1.200 ± 0.104 | 0.174 ± 0.014 | 0.038 ± 0.001 | |
| 10 | 0.040 ± 0.000 | 1.317 ± 0.124 | 0.147 ± 0.024 | 0.038 ± 0.000 | ||
| 15 | 0.041 ± 0.001 | 1.166 ± 0.099 | 0.121 ± 0.041 | 0.042 ± 0.002 | ||
| 4 | 0.041 ± 0.001 | 1.138 ± 0.045 | 0.217 ± 0.047 | 0.040 ± 0.001 | ||
| pH | 5 | 0.041 ± 0.002 | 1.097 ± 0.084 | 0.225 ± 0.035 | 0.041 ± 0.000 | |
| 6 | 0.042 ± 0.001 | 1.126 ± 0.091 | 0.237 ± 0.087 | 0.036 ± 0.002 | ||
| 7 | 0.042 ± 0.000 | 0.042 ± 0.001 | 0.041 ± 0.001 | 0.042 ± 0.000 | ||
| 0 | 0.042 ± 0.000 | 0.041 ± 0.001 | 0.041 ± 0.001 | 0.040 ± 0.001 | ||
| 5 | 0.042 ± 0.001 | 0.635 ± 0.092 | 0.585 ± 0.024 | 0.041 ± 0.001 | ||
| 7.5 | 0.041 ± 0.000 | 0.873 ± 0.052 | 0.581 ± 0.068 | 0.045 ± 0.002 | ||
| Salinity (%) | 10 | 0.041 ± 0.001 | 0.646 ± 0.089 | 0.267 ± 0.042 | 0.042 ± 0.001 | |
| 15 | 0.043 ± 0.002 | 0.710 ± 0.052 | 0.244 ± 0.023 | 0.042 ± 0.000 | ||
| 20 | 0.042 ± 0.002 | 0.823 ± 0.038 | 0.243 ± 0.012 | 0.041 ± 0.000 | ||
| 25 | 0.042 ± 0.001 | 0.748 ± 0.047 | 0.242 ± 0.027 | 0.042 ± 0.001 | ||
| Bacteria Reaction Time (Min) | - | E. coli | B. cereus | S. aureus |
|---|---|---|---|---|
| 0 | 0.040 ± 0.001 | 1.052 ± 0.152 | 1.014 ± 0.087 | 1.151 ± 0.124 |
| 5 | 0.041 ± 0.002 | 1.125 ± 0.102 | 0.812 ± 0.084 | 1.043 ± 0.098 |
| 10 | 0.040 ± 0.000 | 1.248 ± 0.108 | 0.931 ± 0.096 | 1.012 ± 0.125 |
| 20 | 0.042 ± 0.001 | 1.302 ± 0.138 | 0.948 ± 0.090 | 0.957 ± 0.082 |
| 30 | 0.042 ± 0.000 | 1.147 ± 0.085 | 1.027 ± 0.153 | 0.995 ± 0.097 |
| 60 | 0.042 ± 0.001 | 1.217 ± 0.257 | 0.985 ± 0.185 | 1.092 ± 0.127 |
| iTME (1 mg/mL) | Temperature (°C) | Bacteria | Storage Time (d) | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | |||
| − | 37 | E. coli (CFU/mL) | 0 | 0 | 0 | 0 | 0 | 0 |
| − | 37 | 1870 ± 72 | 1680 ± 65 | 2150 ± 74 | 2480 ± 89 | 2520 ± 41 | 2860 ± 87 | |
| + | 4 | 1562 ± 54 | 0 | 0 | 0 | 0 | 0 | |
| + | 18 | 1847 ± 49 | 0 | 0 | 0 | 0 | 0 | |
| + | 37 | 2012 ± 86 | 0 | 0 | 0 | 0 | 0 | |
| − | 37 | B. cereus (CFU/mL) | 0 | 0 | 0 | 0 | 0 | 0 |
| − | 37 | 4852 ± 105 | 3420 ± 47 | 5064 ± 190 | 4890 ± 187 | 5385 ± 215 | 5597 ± 301 | |
| + | 4 | 4181 ± 214 | 0 | 0 | 0 | 0 | 0 | |
| + | 18 | 3988 ± 171 | 0 | 0 | 0 | 0 | 0 | |
| + | 37 | 4773 ± 114 | 0 | 0 | 0 | 0 | 0 | |
| − | 37 | S. aureus (CFU/mL) | 0 | 0 | 0 | 0 | 0 | 0 |
| − | 37 | 160 ± 7 | 350 ± 12 | 470 ± 9 | 620 ± 24 | 2830 ± 57 | 3541 ± 107 | |
| + | 4 | 80 ± 4 | 0 | 0 | 0 | 0 | 0 | |
| + | 18 | 110 ± 14 | 0 | 0 | 0 | 0 | 0 | |
| + | 37 | 230 ± 19 | 0 | 0 | 0 | 0 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.; Lee, S.H.; Goo, T.-W.; Yun, E.-Y. Potential of Antimicrobial Peptide-Overexpressed Tenebrio molitor Larvae Extract as a Natural Preservative for Korean Traditional Sauces. Insects 2022, 13, 381. https://doi.org/10.3390/insects13040381
Hwang D, Lee SH, Goo T-W, Yun E-Y. Potential of Antimicrobial Peptide-Overexpressed Tenebrio molitor Larvae Extract as a Natural Preservative for Korean Traditional Sauces. Insects. 2022; 13(4):381. https://doi.org/10.3390/insects13040381
Chicago/Turabian StyleHwang, Dooseon, Seung Hun Lee, Tae-Won Goo, and Eun-Young Yun. 2022. "Potential of Antimicrobial Peptide-Overexpressed Tenebrio molitor Larvae Extract as a Natural Preservative for Korean Traditional Sauces" Insects 13, no. 4: 381. https://doi.org/10.3390/insects13040381