

Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pollen and Amino Acid Analysis
2.2. Honey Bee Feeding and Hemolymph Collection
2.3. Extraction of Polar Metabolites from Bee Hemolymph
2.4. UPLC-QTOF-MS Analysis
2.5. Data Analysis
3. Results
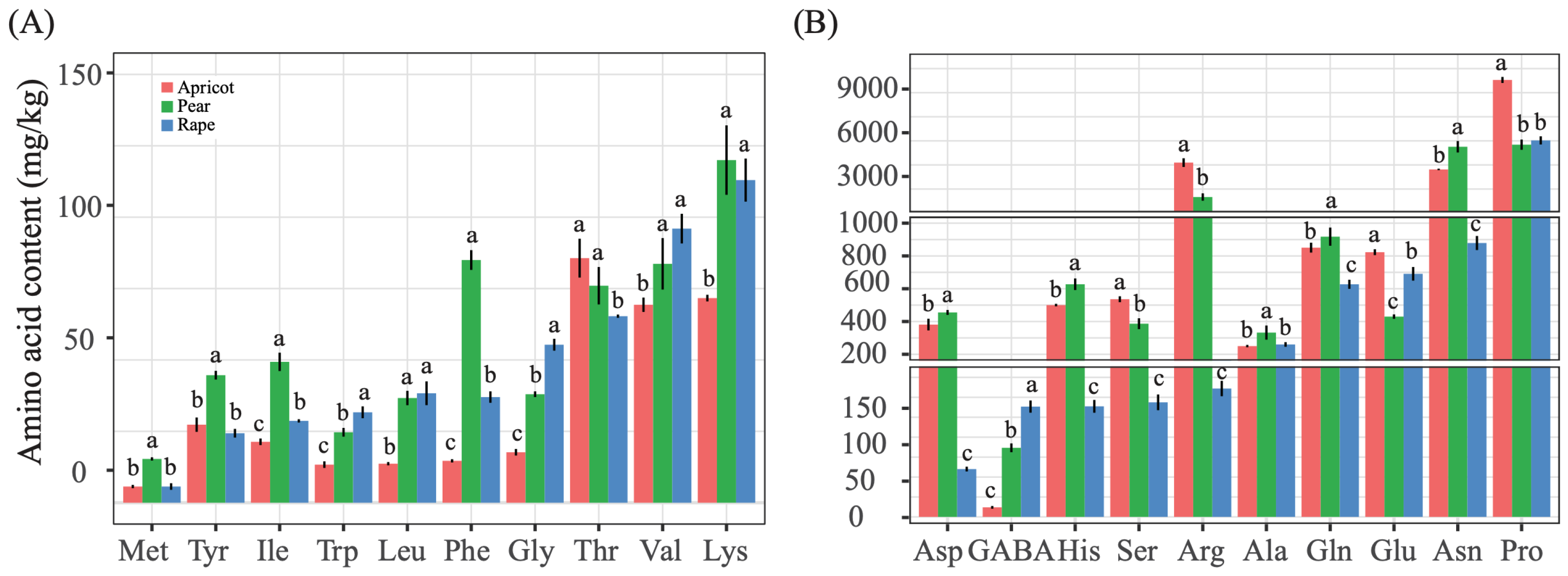
3.1. Amino Acid Components of Pollen
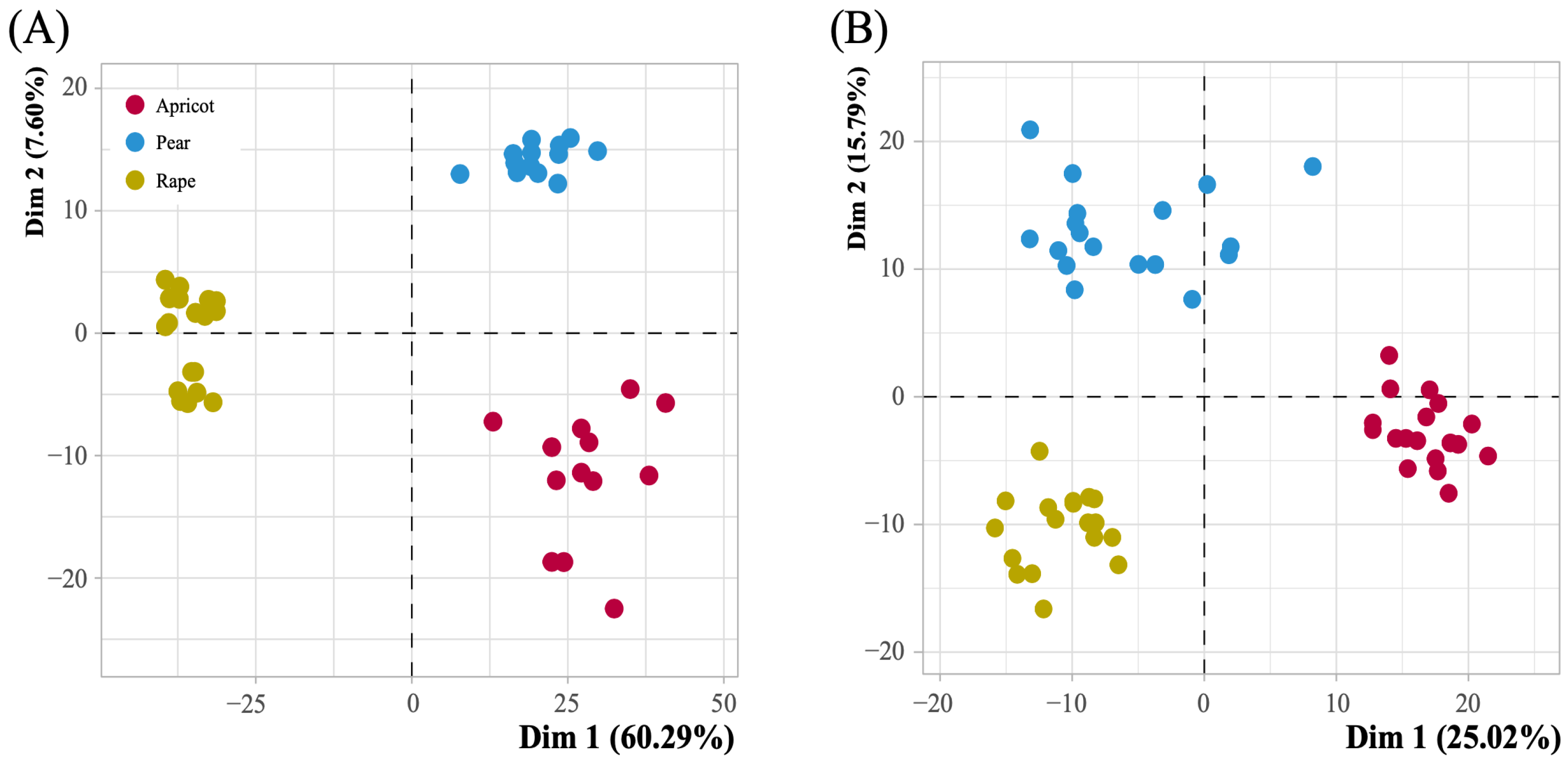
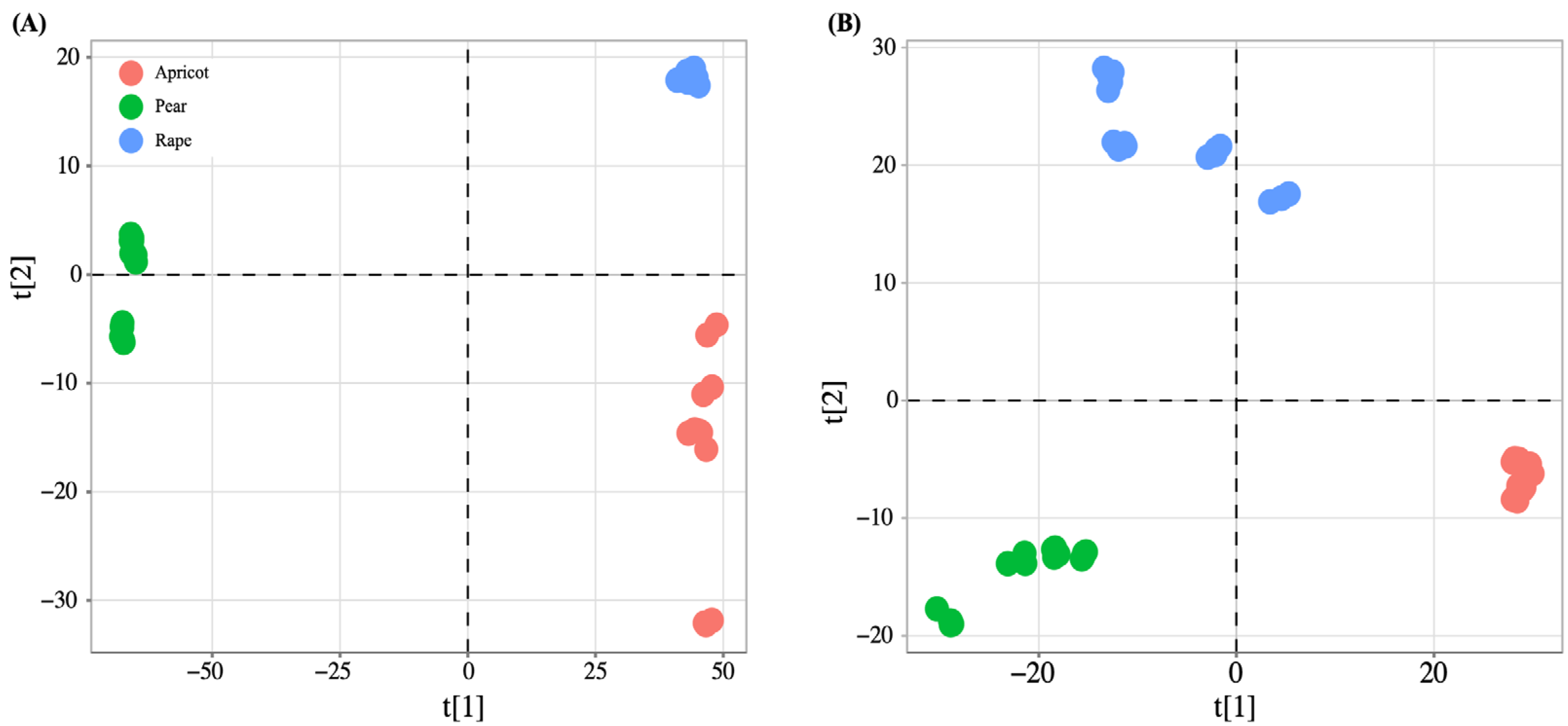
3.2. Metabolite Data Modeling
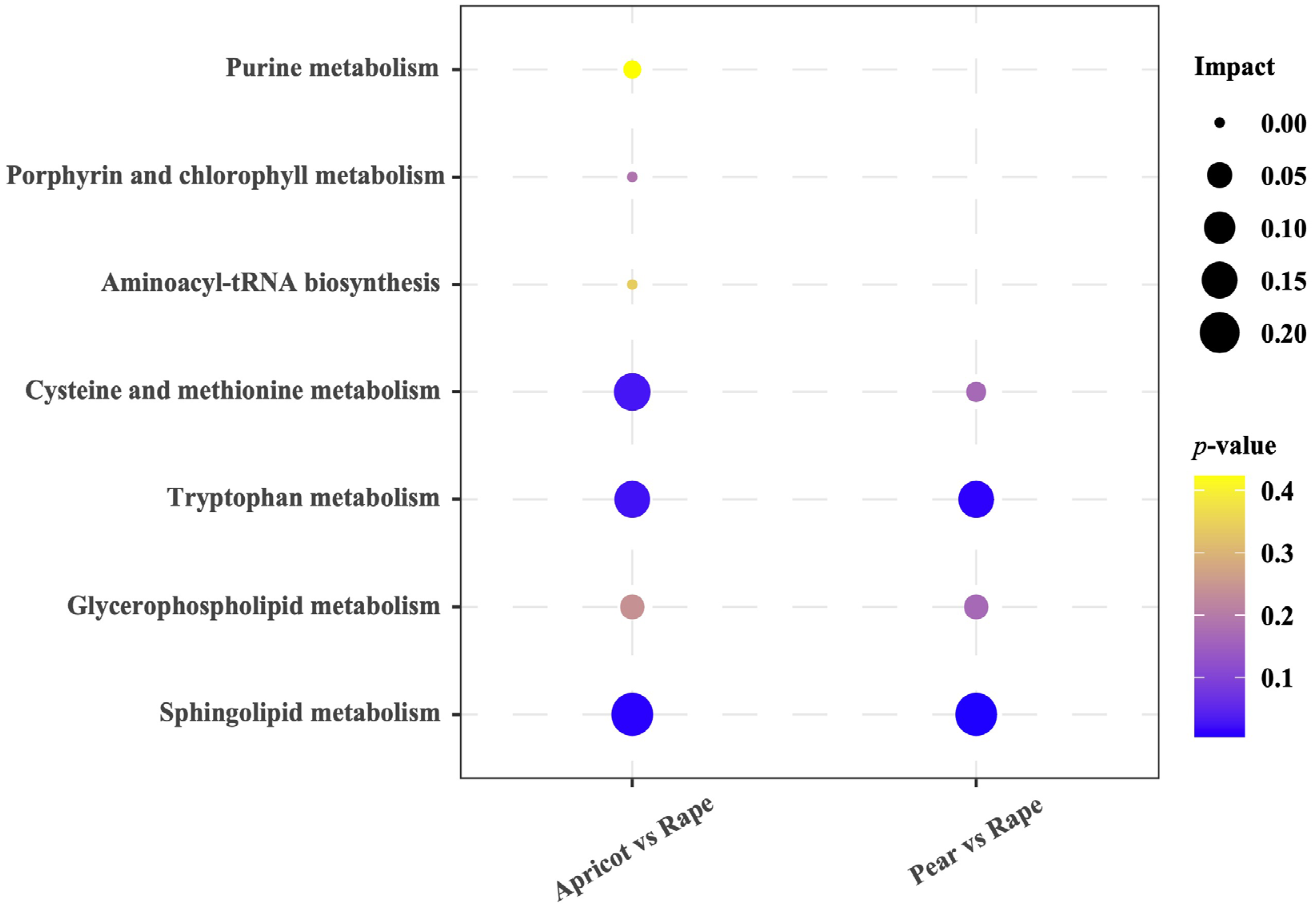
3.3. Metabolic Pathway Analysis
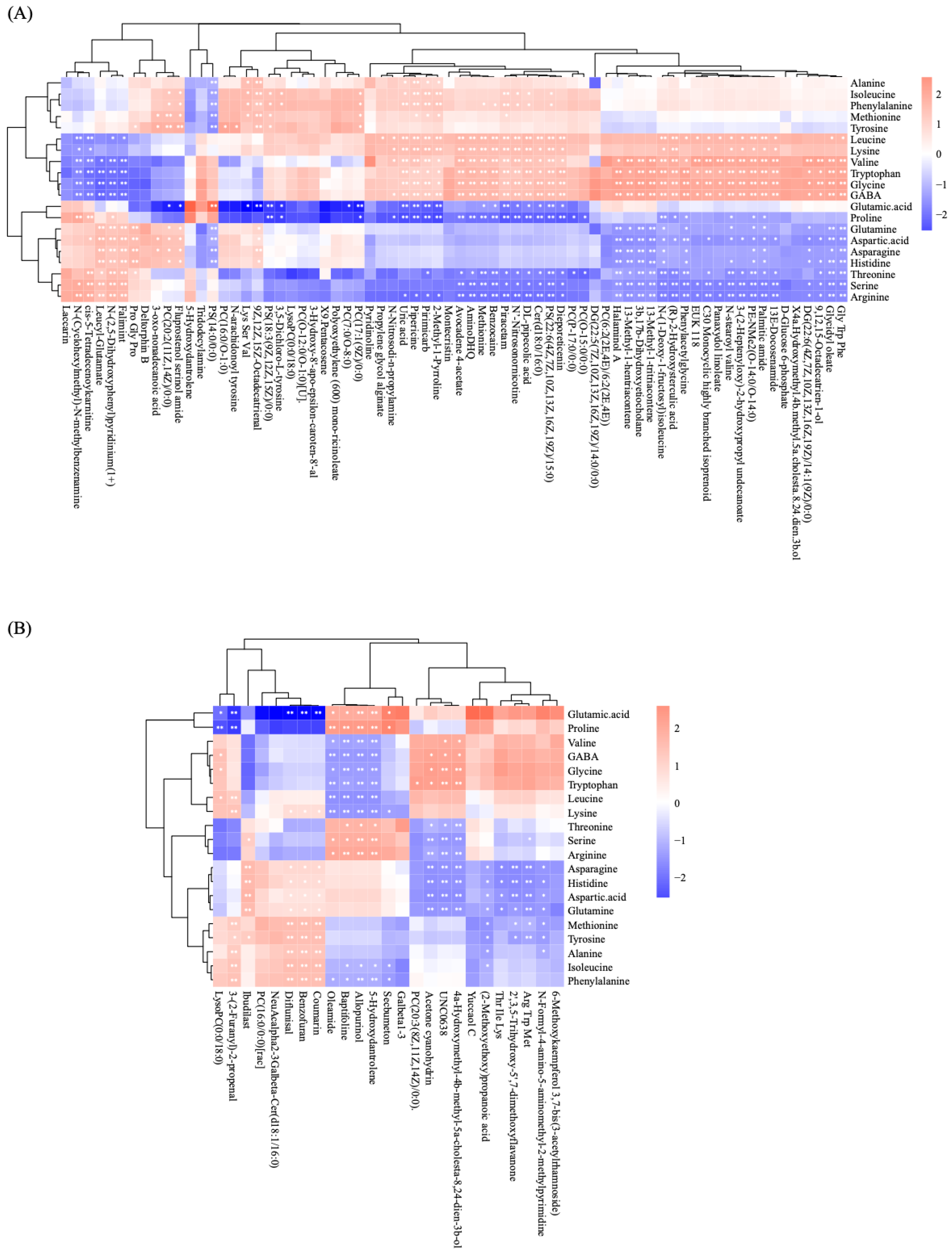
3.4. Correlation Analysis of Amino Acids and Differential Metabolites
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: Digital gene expression analysis of pollen’s nutritive effects on healthy and varroa-parasitized bees. BMC Genom. 2011, 12, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roulston, T.H.; Cane, J.H.; Buchmann, S.L. What governs protein content of pollen: Pollinator preferences, pollen-pistil interactions, or phylogeny? Ecol. Monogr. 2000, 70, 617–643. [Google Scholar]
- Decourtye, A.; Mader, E.; Desneux, N. Landscape enhancement of floral resources for honey bees in agro-ecosystems. Apidologie 2010, 41, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Day, S.; Bey, R.; Mercer, A.; Ogden, S. The nutrient composition of honey bee-collected pollen in Otago, New Zealand. J. Apic. Res. 1990, 29, 138–146. [Google Scholar] [CrossRef]
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Cook, S.M.; Awmack, C.S.; Murray, D.A.; Williams, I.H. Are honey bees’ foraging preferences affected by pollen amino acid composition? Ecol. Entomol. 2003, 28, 622–627. [Google Scholar] [CrossRef]
- Schmidt, J.O.; Thoenes, S.C.; Levin, M.D. Survival of honey bees, Apis mellifera (Hymenoptera: Apidae), fed various pollen sources. Ann. Entomol. Soc. Am. 1987, 80, 176–183. [Google Scholar] [CrossRef]
- McCaughey, W.F.; Gilliam, M.; Standifer, L.N. Amino acids and protein adequacy for honey bees of pollens from desert plants and other floral sources. Apidologie 1980, 11, 75–86. [Google Scholar] [CrossRef]
- De Groot, A.P. Protein and amino acid requirements of the honeybee (Apis mellifica L.). Physiol. Comp. Oecol. 1953, 3, 197–285. [Google Scholar]
- Pang, C.; Dong, K.; Guo, Y.; Ding, G.; Lu, Y.; Guo, Z.; Wu, J.; Huang, J. Effects of three types of pollen on the growth and development of honey bee larvae (Hymenoptera, Apidae). Front. Ecol. Evol. 2022, 10, 870081. [Google Scholar] [CrossRef]
- Su, W.; Ma, W.; Zhang, Q.; Hu, X.; Ding, G.; Jiang, Y.; Huang, J. Honey bee foraging decisions influenced by pear volatiles. Agriculture 2022, 12, 1074. [Google Scholar] [CrossRef]
- Faoro, I.D.; Orth, A.I. Flower visiting insects during the bloom period of Japanese pear orchards in Brazil. Acta Hortic. 2015, 1094, 275–279. [Google Scholar] [CrossRef]
- Monzón, V.H.; Bosch, J.; Retana, J. Foraging behavior and pollinating effectiveness of Osmia cornuta (Hymenoptera: Megachilidae) and Apis mellifera (Hymenoptera: Apidae) on “Comice” pear. Apidologie 2004, 35, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Lan, J.; Ding, G.; Ma, W.; Jiang, Y.; Huang, J. Pollen source affects development and behavioral preferences in honey bees. Insects 2021, 12, 130. [Google Scholar] [CrossRef]
- Hagedorn, H.H.; Moeller, F.E. The rate of pollen consumption by newly emerged honeybees. J. Apic. Res. 1967, 6, 159–162. [Google Scholar] [CrossRef]
- Behmer, S.T. Insect herbivore nutrient regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef]
- Ruedenauer, F.A.; Biewer, N.W.; Nebauer, C.A.; Scheiner, M.; Spaethe, J.; Leonhardt, S.D. Honey bees can taste amino and fatty acids in pollen, but not sterols. Front. Ecol. Evol. 2021, 9, 684175. [Google Scholar] [CrossRef]
- Ruedenauer, F.A.; Wohrle, C.; Spaethe, J.; Leonhardt, S.D. Do honeybees (Apis mellifera) differentiate between different pollen types? PLoS ONE 2018, 13, e205821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamani, V.; Ory, P.; Bodet, P.; Murillo, L.; Graber, M. Untargeted metabolomics reveals a complex impact on different metabolic pathways in scallop Mimachlamys varia (Linnaeus, 1758) after short-term exposure to copper at. Metabolites 2021, 11, 862. [Google Scholar] [CrossRef] [PubMed]
- Horgan, R.; Clancy, O.; Myers, J.; Baker, P. An overview of proteomic and metabolomic technologies and their application to pregnancy research. BJOG—Int. J. Obstet. Gynaecol. 2009, 116, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Bundy, J.G.; Davey, M.P.; Viant, M.R. Environmental metabolomics: A critical review and future perspectives. Metabolomics 2009, 5, 3–21. [Google Scholar] [CrossRef]
- Wang, L.; Meeus, I.; Rombouts, C.; Van Meulebroek, L.; Vanhaecke, L.; Smagghe, G. Metabolomics-based biomarker discovery for bee health monitoring: A proof of concept study concerning nutritional stress in Bombus terrestris. Sci. Rep. 2019, 9, 11423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danihlík, J.; Škrabišová, M.; Lenobel, R.; Šebela, M.; Omar, E.; Petřivalský, M.; Crailsheim, K.; Brodschneider, R. Does the pollen diet influence the production and expression of antimicrobial peptides in individual honey bees? Insects 2018, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunner, F.S.; Schmid-Hempel, P.; Barribeau, S.M. Protein-poor diet reduces host-specific immune gene expression in Bombus terrestris. Proc. R. Soc. B 2014, 281, 20140128. [Google Scholar] [CrossRef] [Green Version]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Domaciuk, M.; Krutmuang, P.; Paleolog, J. A new method for quick and easy hemolymph collection from Apidae adults. PLoS ONE 2017, 12, e170487. [Google Scholar] [CrossRef]
- Li, Q.; Sun, M.; Wan, Z.; Liang, J.; Betti, M.; Hrynets, Y.; Xue, X.; Wu, L.; Wang, K. Bee pollen extracts modulate serum metabolism in lipopolysaccharide-lnduced acute lung lnjury mice with anti-inflammatory effects. J. Agric. Food Chem. 2019, 67, 7855–7868. [Google Scholar] [CrossRef]
- Crailsheim, K. The protein balance of the honey bee worker. Apidologie 1990, 21, 417–429. [Google Scholar] [CrossRef]
- Nepi, M. Beyond nectar sweetness: The hidden ecological role of non-protein amino acids in necta. J. Ecol. 2014, 102, 108–115. [Google Scholar] [CrossRef]
- Carlesso, D.; Smargiassi, S.; Bertelli, E.P.G.; Baracchi, D. Nectar non-protein amino acids (NPAAs) do not change nectar palatability but enhance learning and memory in honey bees. Sci. Rep. 2011, 11, 11721. [Google Scholar] [CrossRef] [PubMed]
- Paoli, P.P.; Donley, D.; Stabler, D.; Saseendranath, A.; Nicolson, S.W.; Simpson, S.J.; Wright, G.A. Nutritional balance of essential amino acids and carbohydrates of the adult worker honeybee depends on age. Amino Acids 2014, 46, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teulier, L.; Weber, J.; Crevier, J.; Darveau, C. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, C.; Shafir, S.; Yehonatan, L.; Palmer, R.G.; Thornburg, R. A novel role for proline in plant floral nectars. Sci. Nat. 2006, 93, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Baquet, A.; Lavoinne, A.; Hue, L. Comparison of the effects of various amino acids on glycogen synthesis, lipogenesis and ketogenesis in isolated rat hepatocytes. Biochem. J. 1991, 273, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Inouye, D.W.; Waller, G.D. Responses of honey bees (Apis mellifera) to amino acid solutions mimicking floral nectars. Ecology 1984, 65, 618–625. [Google Scholar] [CrossRef]
- Shayman, J.A. Sphingolipids. Kidney Int. 2000, 58, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, S.; Merrill, A.H., Jr. Sphingolipid metabolism and cell growth regulation. FASEB J. 1996, 10, 1388–1397. [Google Scholar] [CrossRef]
- Beard, C.B.; Benedict, M.Q.; Primus, J.P.; Finnerty, V.; Collins, F.H. Eye pigments in wild-type and eye-color mutant strains of the african malaria vector Anopheles gambiae. J. Hered. 1995, 86, 375–380. [Google Scholar] [CrossRef]
- Linzen, B. The tryptophan → ommochrome pathway in insects. Adv. Insect Physiol. 1974, 10, 117–246. [Google Scholar]
- Zhang, Z.; Mu, X.; Cao, Q.; Shi, Y.; Hu, X.; Zheng, H. Honeybee gut Lactobacillus modulates host learning and memory behaviors via regulating tryptophan metabolism. Nat. Commun. 2022, 13, 2037. [Google Scholar] [CrossRef]
- Fengkui, Z.; Baohua, X.; Ge, Z.; Hongfang, W. The appropriate supplementary level of tryptophan in the diet of Apis mellifera (Hymenoptera: Apidae) worker bees. J. Insect Sci. 2015, 15, 161. [Google Scholar] [CrossRef]
- Harris, J.W.; Woodring, J. Effects of dietary precursors to biogenic amines on the behavioural response from groups of caged worker honey bees (Apis mellifera) to the alarm pheromone component isopentyl acetate. Physiol. Entomol. 1999, 24, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Cerstiaens, A.; Huybrechts, J.; Kotanen, S.; Lebeau, I.; Meylaers, K.; De Loof, A.; Schoofs, L. Neurotoxic and neurobehavioral effects of kynurenines in adult insects. Biochem. Biophys. Res. Commun. 2003, 312, 1171–1177. [Google Scholar] [CrossRef]
- Lee, B.C.; Kaya, A.; Ma, S.; Kim, G.; Gerashchenko, M.V.; Yim, S.H.; Hu, Z.; Harshman, L.G.; Gladyshev, V.N. Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nat. Commun. 2014, 5, 3592. [Google Scholar] [CrossRef] [Green Version]
- Grandison, R.C.; Piper, M.D.W.; Partridge, L. Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila. Nature 2009, 462, 1061–1064. [Google Scholar] [CrossRef] [Green Version]
- Winblad, B. Piracetam: A review of pharmacological properties and clinical uses. CNS Drug Rev. 2005, 11, 169–182. [Google Scholar] [CrossRef]
- Paula-Barbosa, M.M.; BrandBao, F.; Pinho, M.C.; Andrade, J.P.; Madeira, M.D.; Cadete-Leite, A. The effects of piracetam on lipofuscin of the rat cerebellar and hippocampal neurons after long-term alcohol treatment and withdrawal: A quantitative study. Alcohol. Clin. Exp. Res. 1991, 15, 834–838. [Google Scholar] [CrossRef]
- Gage, S.L.; Calle, S.; Jacobson, N.; Carroll, M.; DeGrandi-Hoffman, G. Pollen alters amino acid levels in the honey bee brain and this relationship changes with age and parasitic stress. Front. Neurosci. 2020, 14, 231. [Google Scholar] [CrossRef]
- Berger, B.; Crailsheim, K.; Leonhard, B. Proline, leucine and phenylalanine metabolism in adult honeybee drones (Apis mellifica carnica Pollm). Insect Biochem. Mol. Biol. 1997, 27, 587–593. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, H.; Ding, G.; Jia, G.; Feng, M.; Huang, J. Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens. Insects 2023, 14, 37. https://doi.org/10.3390/insects14010037
Chang H, Ding G, Jia G, Feng M, Huang J. Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens. Insects. 2023; 14(1):37. https://doi.org/10.3390/insects14010037
Chicago/Turabian StyleChang, Hongcai, Guiling Ding, Guangqun Jia, Mao Feng, and Jiaxing Huang. 2023. "Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens" Insects 14, no. 1: 37. https://doi.org/10.3390/insects14010037
APA StyleChang, H., Ding, G., Jia, G., Feng, M., & Huang, J. (2023). Hemolymph Metabolism Analysis of Honey Bee (Apis mellifera L.) Response to Different Bee Pollens. Insects, 14(1), 37. https://doi.org/10.3390/insects14010037