
Sphenophorus levis Behavior Studies: Evaluating Insect Attractiveness or Repellency to One Insecticide Treatment and Assessing Nocturnal Insect Activity and Location Pattern


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1—Insecticide Repellent/Attractive Activity
2.2. Experiment 2—Nocturnal Adult Activity Pattern
2.3. Data Analysis
3. Results
3.1. Experiment 1—Insecticide Repellent/Attractive Activity
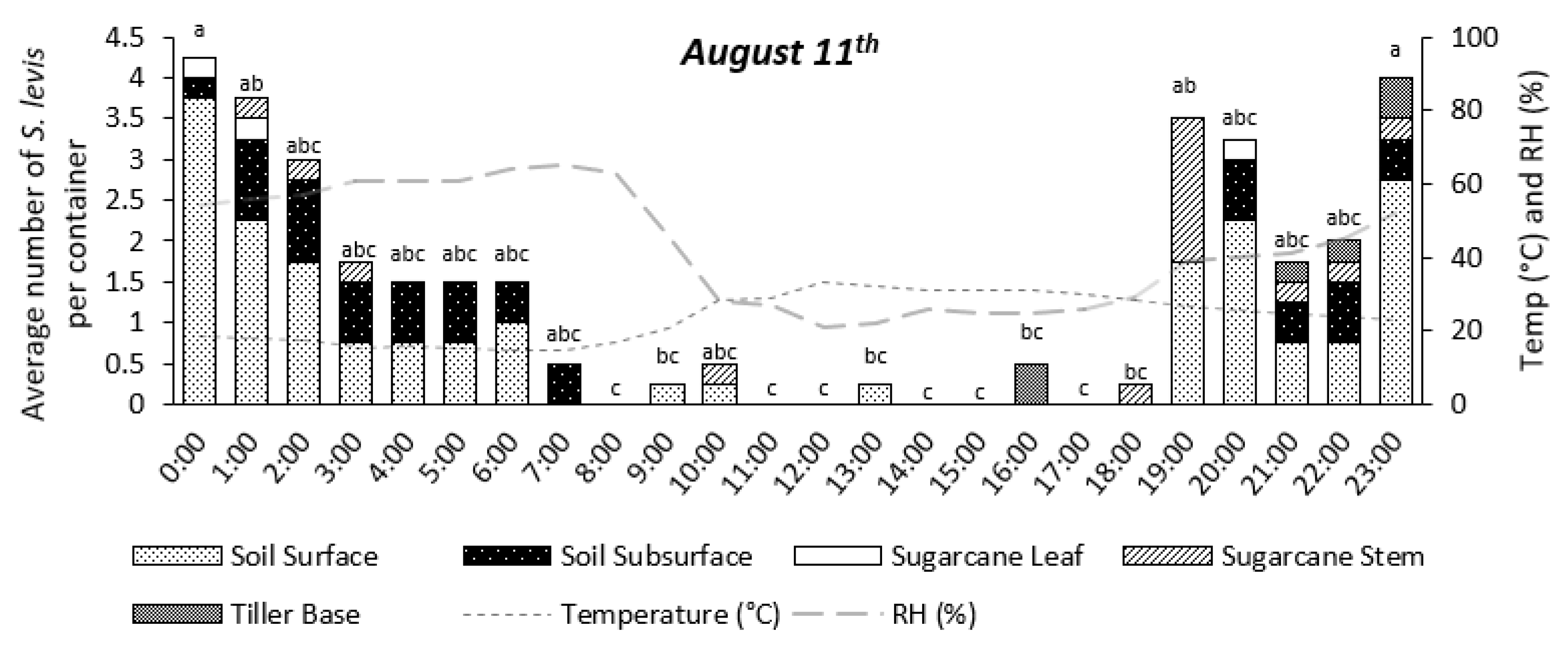
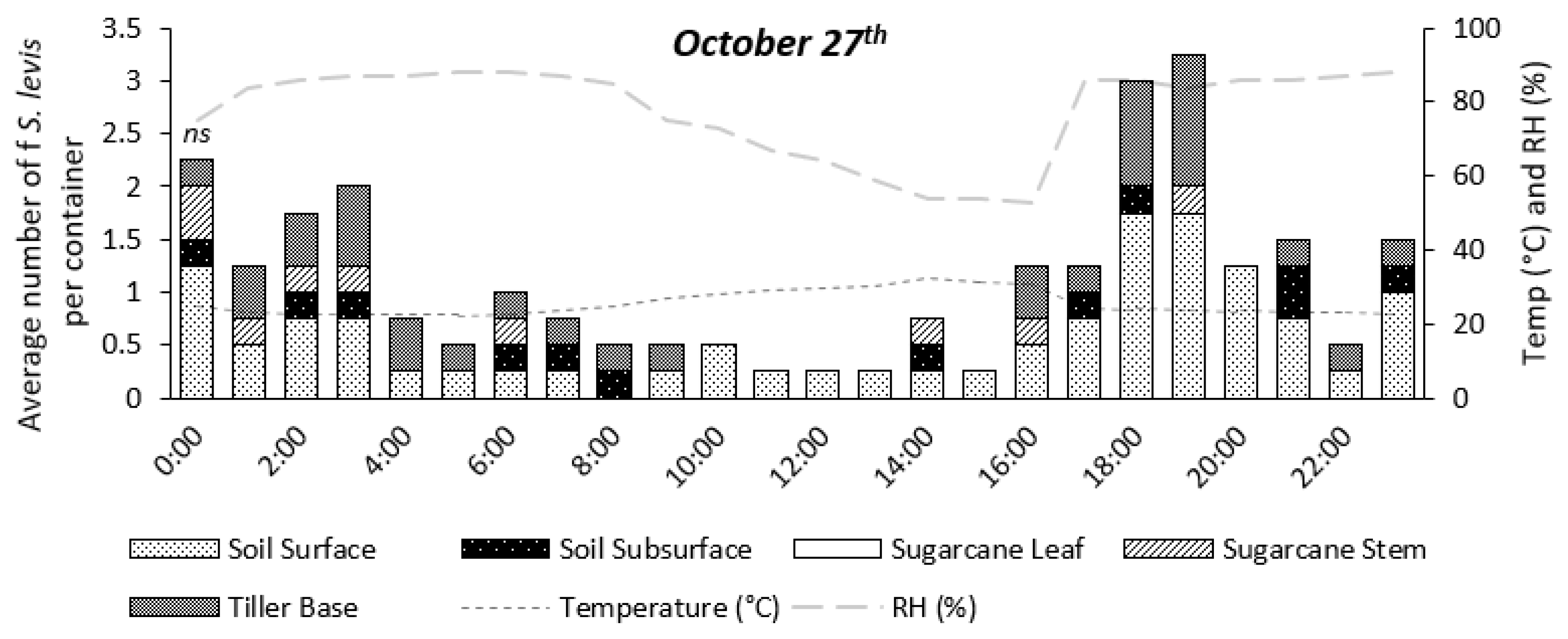
3.2. Experiment 2—Nocturnal Adult Activity Pattern
4. Discussion
4.1. Experiment 1—Insecticide Repellent/Attractive Activity
4.2. Experiment 2—Nocturnal Adult Activity Pattern
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dinardo-Miranda, L.L.; Fracasso, J.V. Sugarcane straw and the populations of pests and nematoids. Sci. Agric. 2013, 70, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Dinardo-Miranda, L.L.; Fracasso, J.V.; Cabral, S.B.; Valério, W.; Gonçalves, R.D.; Beltrame, J.Á. Eficiência de inseticidas aplicados em soqueiras de cana-de-açúcar no controle de Sphenophorus levis. STAB 2006, 24, 34–37. [Google Scholar]
- Tavares, F.M. Avaliação de Nematóides Entomopatogênicos Contra o Bicudo da Cana-de Açúcar Sphenophorus levis Vaurie, 1978, e Efeito da Associação Desses Agentes Com Inseticidas Químicos. Master’s Thesis, Agronomy, Universidade Estadual Paulista, Botucatu, SP, Brazil, 2006; p. 61. [Google Scholar]
- Alencar, M.A.V. Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae): Caracterização Macroscópica e Determinação de Inseticida e Época de Aplicação Para Controle. Ph.D. Thesis, Agricultural Entomology, Universidade Estadual Paulista, Jaboticabal, SP, Brazil, 2016; p. 68. [Google Scholar]
- Dobrin, G.C.; Hammond, R.B. The antifeeding activity of selected pyrethroids towards the Mexican Bean Beetle (Coleoptera: Coccinellidae). J. Kans. Entomol. Soc. 1985, 8, 422–427. [Google Scholar]
- Easton, A.H.; Goulson, D. The neonicotinoid insecticide imidacloprid repels pollinating flies and beetles at field-realistic concentrations. PLoS ONE. 2013, 8, e54819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canassa, F. Distribuição Especial, Efeito do Manejo da Palha Pós-Colheita e da Aplicação de Beauveria bassiana (Balsamo) Vuillemin (Hypocreales: Cordycipitaceae) na Ocorrência de Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae) em Cana-De-Açúcar. Master’s Thesis, Entomology, Universidade de São Paulo, Piracicaba, SP, Brazil, 2014; p. 98. [Google Scholar]
- Zarbin, P.H.G.; Arrigoni, E.B.; Reckziegel, A.; Moreira, J.Á.; Baraldi, P.T.; Vieira, P.C. Identification of male-specific chiral compound from the sugarcane weevil Sphenophorus levis. J. Chem. Ecol. 2003, 29, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Izeppi, T.S. Distribuição Espacial e Dinâmica Populacional de Sphenophorus levis (Coleoptera: Curculionidae) em Cana-De-Açúcar. Master’s Thesis, Entomology, Universidade Estadual Paulista, Jaboticabal, SP, Brazil, 2015; p. 74. [Google Scholar]
- Rosa, J.O. Infestação de Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae) em Relação aos Pontos de Transferência de Cana-De-Açúcar. Master’s Thesis, Faculty of Agricultural and Veterinary Sciences, Jaboticabal, SP, Brazil, 2022; p. 28. [Google Scholar]
- Precetti, A.A.C.M.; Arrigoni, E.B. Aspectos bioecológicos e controle do besouro Sphenophorus levis Vaurie, 1978 (Coleoptera, Curculionidae) em cana-de-açúcar. Bol. Téc. Copersucar Ed. Espec. 1990, 1–15. [Google Scholar]
- Simi, L.D. Controle de Sphenophorus levis e Conotrachelus humeropictus Pelo uso Combinado de Nematoides e Fungos Entomopatogênicos. Master’s Thesis, Plant Protection, Universidade Estadual Paulista, Botucatu, SP, Brazil, 2014. [Google Scholar]
- Kindler, S.D.; Spomer, S.M. Observations on the biology of the bluegrass billbug, Sphenophorus parvulus Gyllenhal (Coleoptera: Curculionidae), in an eastern Nebraska sod field. J. Kans. Entomol. Soc. 1986, 59, 26–31. [Google Scholar]
- Casteliani, A.; Martins, L.F.; Cardoso, J.F.M.; Silva, M.S.O.; Silva, R.S.A.; Chacon-Orozco, J.G.; Casteliani, A.G.B.; Půža, V.; Harakava, R.; Leite, L.G. Behavioral aspects of Sphenophorus levis (Coleoptera: Curculionidae), damage to sugarcane and its natural infection by Steinernema carpocapsae (Nematoda: Rhabditidae). Crop Prot. 2020, 137, 105262. [Google Scholar] [CrossRef]
- Gold, C.S.; Pena, J.E.; Karamura, E.B. Biology and integrated pest management for the banana weevil Cosmopolites sordidus (Germar) (Coleoptera: Curculionidae). Integr. Pest. Manag. Rev. 2001, 6, 79–155. [Google Scholar] [CrossRef]
- Huang, T.I.; Buss, E.A. Billbug (Coleoptera: Curculionidae) species composition, abundance, seasonal activity, and developmental time in Florida. J. Econ. Entomol. 2009, 102, 309–314. [Google Scholar] [CrossRef]
- Mazzonetto, F.; Vendramin, J.D. Efeito de pós de origem vegetal sobre Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae) em feijão armazenado. Neotr. Entomol. 2003, 31, 145–149. [Google Scholar] [CrossRef] [Green Version]
- Fouad, H.A.; Faroni, L.R.D.; Ribeiro, R.C.; Tavares, W.D.S.; Petacci, F. Extraction and repellent activity of Lepidoploa aurea and Memora nodosa against stored grain and by product pests. Vie Milieu 2012, 62, 11–15. [Google Scholar]
- Viteri Jumbo, L.O.; Faroni, L.R.A.; Oliveira, E.E.; Pimentel, M.A.; Silva, G.N. Potential use of clove and cinnamon essential oils to control the bean weevil, Acanthoscelides obtectus Say, in small storage units. Ind. Crops Prod. 2014, 56, 27–34. [Google Scholar] [CrossRef]
- Raij, B.V.; Andrade, J.C.; Cantarella, H.; Quaggio, J.Á. Análise Química para Avaliação da Fertilidade de Solos Tropicais; Instituto Agronômico: Campinas, Brazil, 2001; p. 285. [Google Scholar]
- Ferreira, P.H.U. Effect of Liquid and Solid Insecticide Applications and Insect Behavior on Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae) Control in Sugarcane. Ph.D. Thesis, Agricultural Entomology, Universidade Estadual Paulista, Jaboticabal, SP, Brazil, 2022; p. 163. [Google Scholar]
- Pérez, K.G. Eficiência de Iscas Tóxicas no Controle de Adultos de Sphenophorus levis Vaurie (Coleoptera: Curculinidae) em Cana-de-Açúcar (Saccharum officinarum L.). Master’s Thesis, Entomology, Universidade de São Paulo, Piracicaba, SP, Brazil, 2008. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R. v. 1.4.1717; RStudio, Inc.: Boston, MA, Boston, 2021; Available online: http://www.rstudio.com (accessed on 12 August 2021).
- Moral, R.A.; Hinde, J.; Demétrio, C.G.B. Half-Normal Plots and Overdispersed Models in R: The hnp Package. J. Stat. Softw. 2017, 81, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package v. 1.3.5.1. 2019. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 14 August 2021).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion (accessed on 15 August 2021).
- Martins, L.F.; Tonelli, M.; Bento, J.M.S.; Bueno, C.J.; Leite, L.G. Attraction of the sugarcane billbug, Sphenophorus levis, to vinasse and its volatile composition. Chemoecology 2020, 30, 205–214. [Google Scholar] [CrossRef]
- Illescas-Riquelme, C.P.; Llanderal-Cázares, C.; Ruiz-Montiel, C.; González-Hernández, H.; Alatorre-Rosas, R.; Cruz-López, L.; Rojas, J.C. Evidence for male-produced aggregation pheromone in Sphenophorus incurrens (Coleoptera: Curculionidae). Fla Entomol. 2016, 99, 522–527. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, D.C. Ecology and Behavior of the Hunting Billbug Sphenophorus venatus vestitus in Warm-Season Turfgrass. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 2014; p. 115. [Google Scholar]
- Veleri, S.; Rieger, D.; Helfrich-Förster, C.; Stanewsky, R. Hofbauer-Buchner eyelet affects circadian photosensitivity and coordinates TIM and PER expression in Drosophila clock neurons. J. Biol. Rhythms. 2007, 22, 29–42. [Google Scholar] [CrossRef]
- Klowden, M.J. Physiological Systems in Insects, 3rd ed.; Academic Press: London, UK, 2013; p. 661. [Google Scholar]
- Degaspari, N.; Botelho, N.P.S.; Almeida, L.C.; Castilho, H.J. Biologia de Sphenophorus levis Vaurie, 1978 (Coleoptera: Curculionidae) em dieta artificial e no campo. Pesqui. Agropecu. Bras. 1987, 22, 556–558. [Google Scholar]
- Izeppi, T.S.; Dinardo-Miranda, L.L.; Barbosa, J.C. Flutuação Populacional de Sphenophorus levis em cana-de-açúcar. In VIII Workshop Agroenergia Matérias Primas; IAC: Ribeirão Preto, Brazil, 2014; ISBN 978-85-85564-30-8. [Google Scholar]
- Durant, J.A. Influence of temperature relative humidity date and time of day on activity of the adult southern corn billbug Sphenophorus callosus on corn Zea mays. J. Agric. Entomol. 1985, 1, 20–26. [Google Scholar]
- Polato, S.A.; Oliveira, N.C. Eficiência do controle da lagarta-do-cartucho na cultura do milho em função de diferentes horários de aplicação de inseticida. Campo Digit. 2011, 1, 44–53. [Google Scholar]
- Rincão, R.O.; Pivaro, R.S.; Xavier, A.M.; Ferraz, M.; Domene, R.A.; Michelotto, M.D. Aplicação noturna de inseticidas para controle do percevejo-preto em amendoim. South Am. Sci. 2020, 2, e2070. [Google Scholar] [CrossRef]
- Dinardo-Miranda, L.L. Nematoides e Pragas de Cana-De-Açúcar; Instituto Agronômico: Campinas, SP, Brazil, 2014; p. 400. ISBN 978-85-85564-31-5. [Google Scholar]
- Sondhi, Y.; Jo, N.J.; Alpizar, B.; Markee, A.; Dansby, H.E.; Currea, J.P.; Fabian, S.T.; Ruiz, C.; Barredo, E.; Allen, P.; et al. Portable locomotion activity monitor (pLAM): A cost-effective setup for robust activity tracking in small animals. Methods Ecol. Evol. 2022, 4, 805–812. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urach Ferreira, P.H.; Ferreira, M.d.C. Sphenophorus levis Behavior Studies: Evaluating Insect Attractiveness or Repellency to One Insecticide Treatment and Assessing Nocturnal Insect Activity and Location Pattern. Insects 2023, 14, 205. https://doi.org/10.3390/insects14020205
Urach Ferreira PH, Ferreira MdC. Sphenophorus levis Behavior Studies: Evaluating Insect Attractiveness or Repellency to One Insecticide Treatment and Assessing Nocturnal Insect Activity and Location Pattern. Insects. 2023; 14(2):205. https://doi.org/10.3390/insects14020205
Chicago/Turabian StyleUrach Ferreira, Pedro Henrique, and Marcelo da Costa Ferreira. 2023. "Sphenophorus levis Behavior Studies: Evaluating Insect Attractiveness or Repellency to One Insecticide Treatment and Assessing Nocturnal Insect Activity and Location Pattern" Insects 14, no. 2: 205. https://doi.org/10.3390/insects14020205