Bombyx mori Ecdysone Receptor B1 May Inhibit BmNPV Infection by Triggering Apoptosis

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworms and BmN Cells
2.2. Preparation and Detection of BmNPV
2.3. Sample Preparation for Expression Pattern Analysis
2.4. Bioinformatics Analysis
2.5. RNA Extraction and the First Strand cDNA Synthesis
2.6. Real-Time Quantitative PCR (RT-qPCR)
2.7. Synthesis of siRNA and Transfection
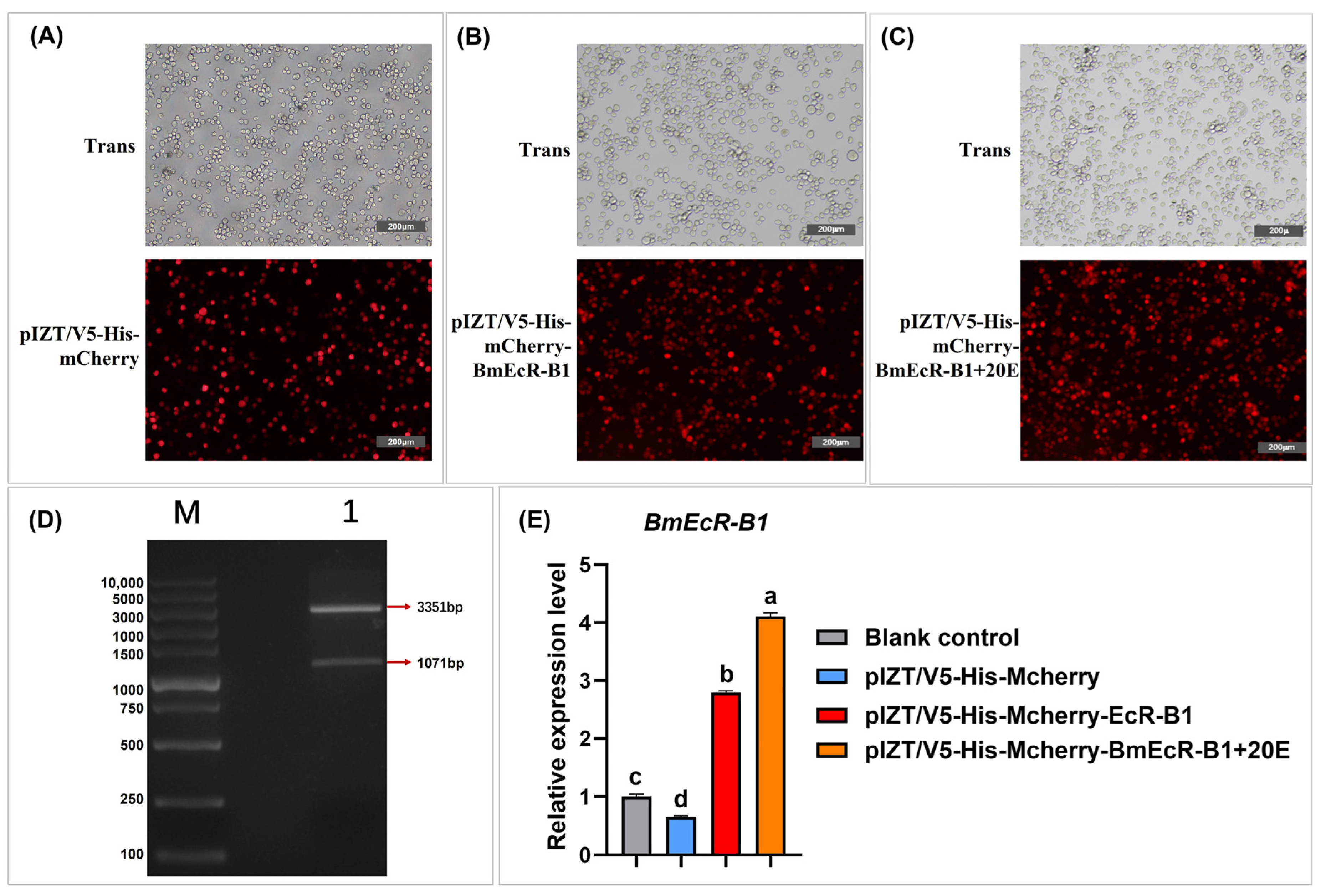
2.8. Construction of the Overexpression Vector
2.9. The Treatment of 20E on Silkworm Larvae
2.10. The Effect of 20E Treatment on BmEcR-B1 Expression and BmNPV Infection in BmN Cells
2.11. Caspase Activity Assay and Apoptotic Bodies Detection
2.12. Statistics Analysis
3. Results
3.1. Bioinformatics Analysis
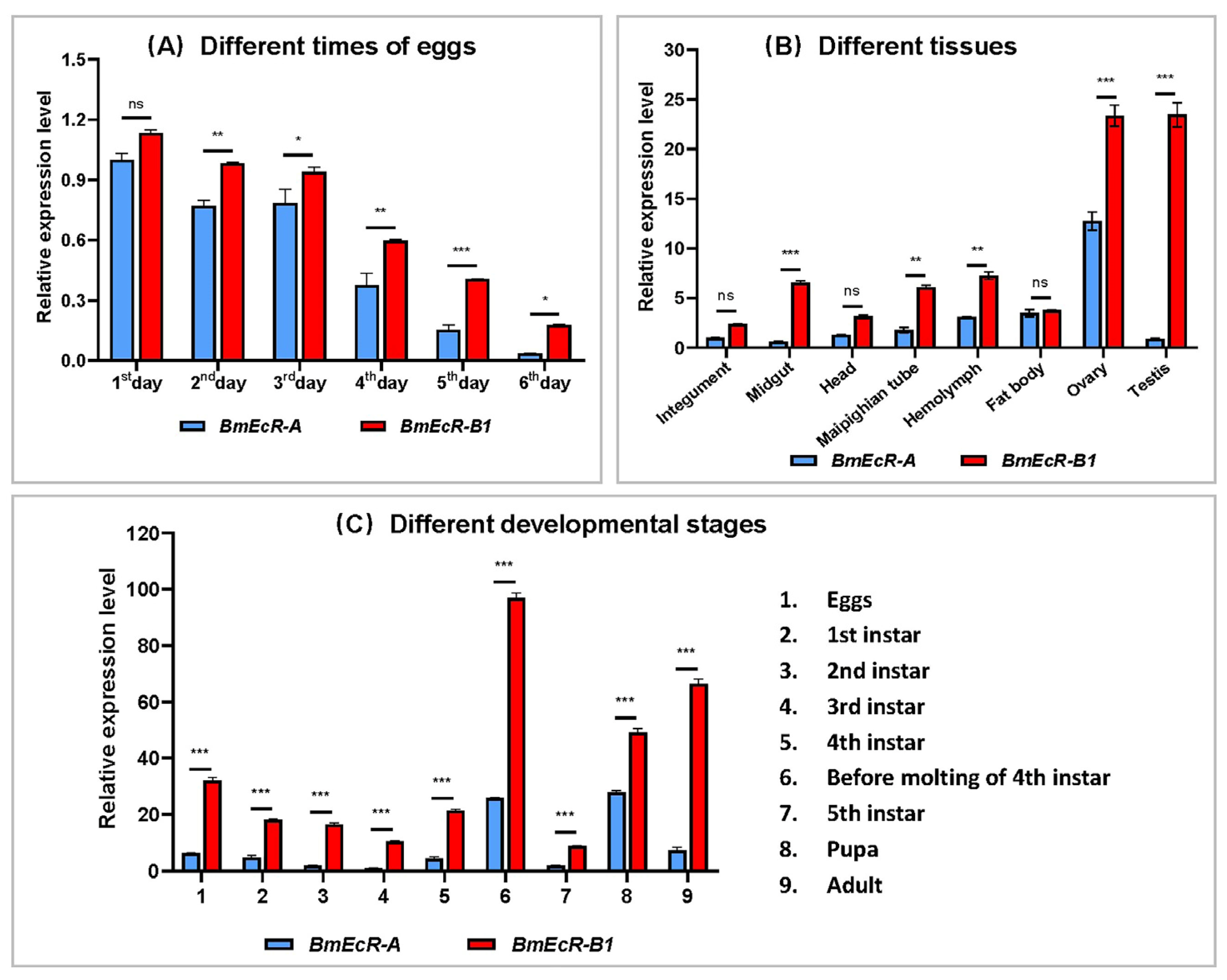
3.2. The Temporal and Spatial Expression Profile of BmEcR-B1 and BmEcR-A
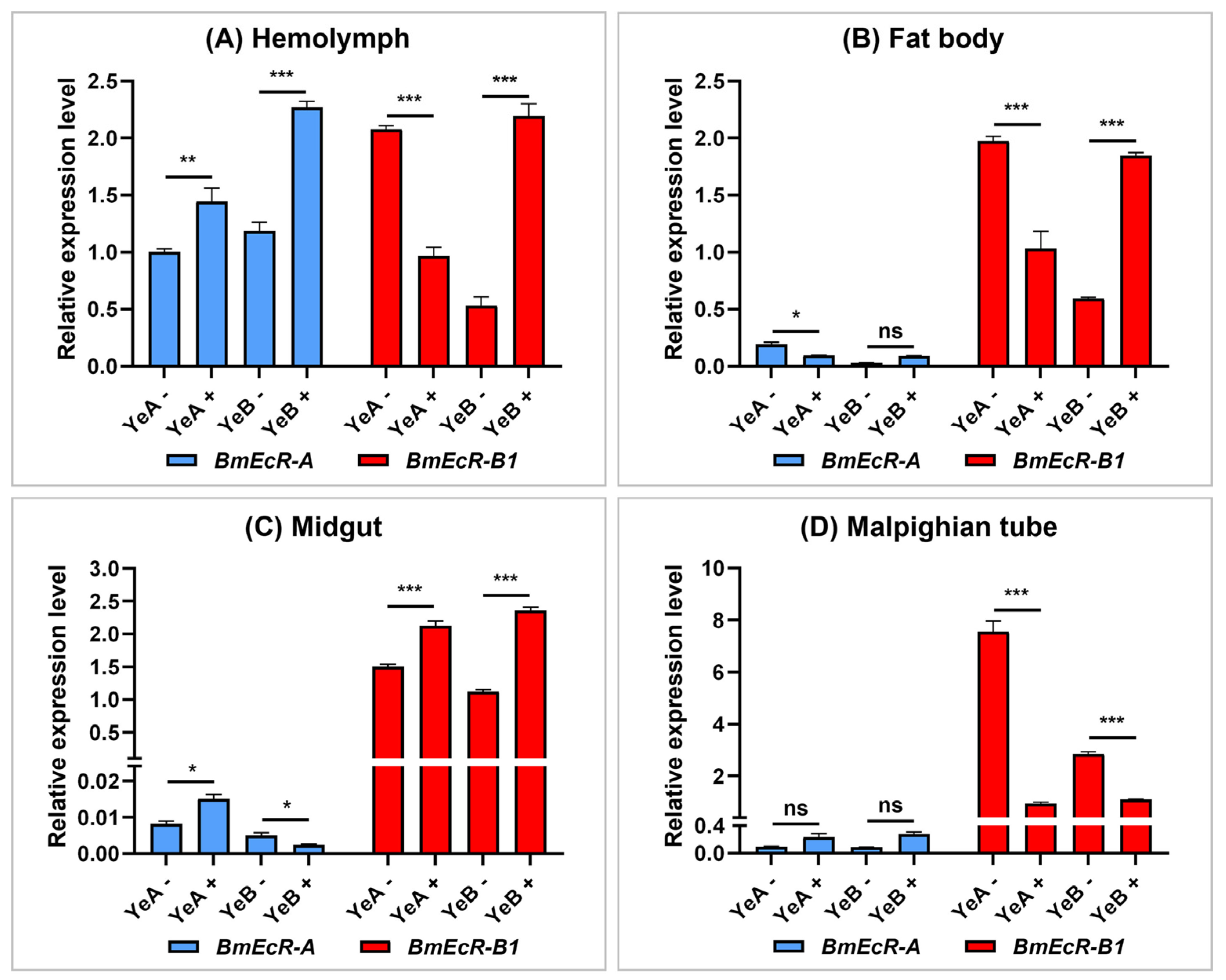
3.3. BmEcR-B1 Showed a Significant Response to BmNPV Infection
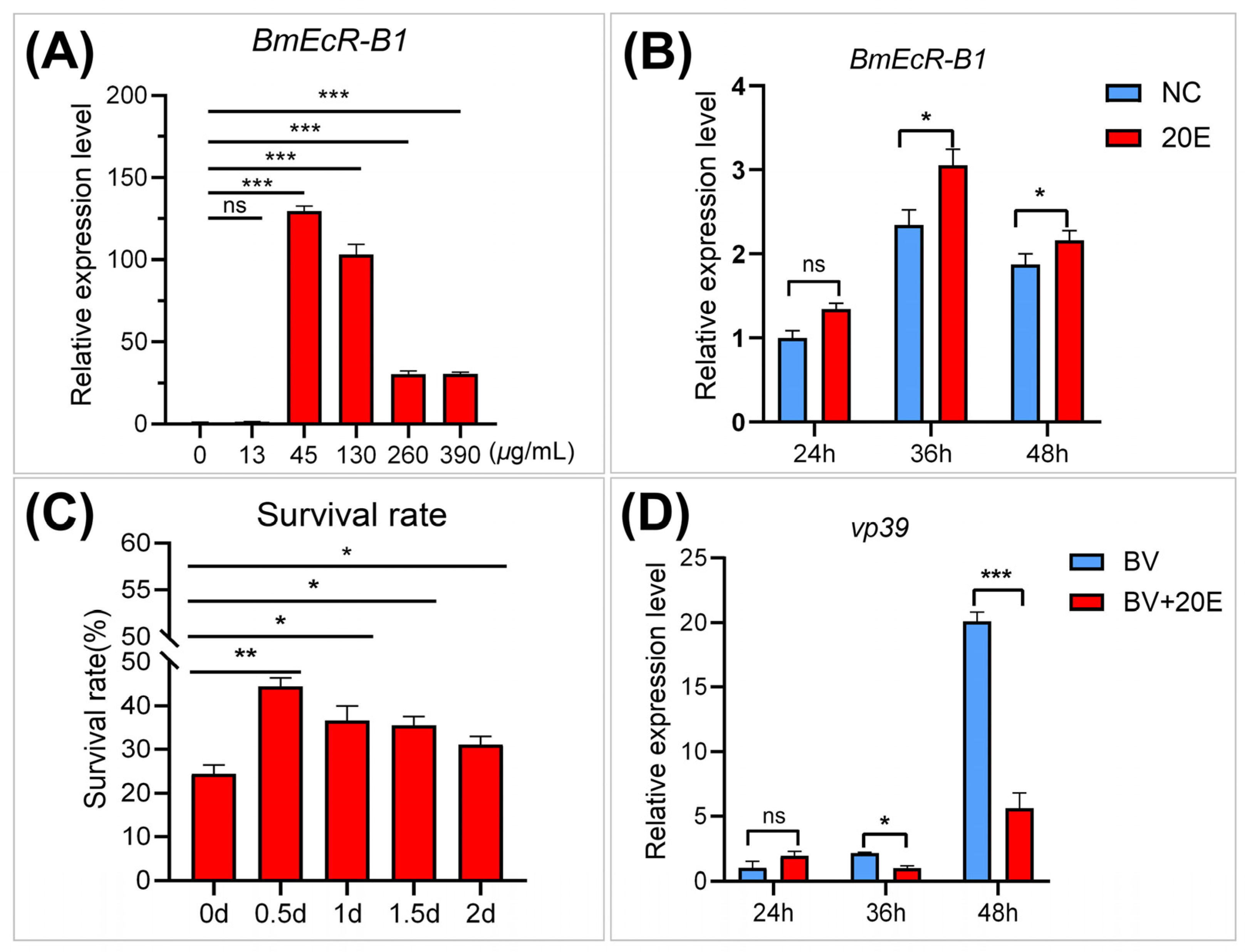
3.4. E activates BmEcR-B1 Expression and Inhibits BmNPV Infection In Vivo
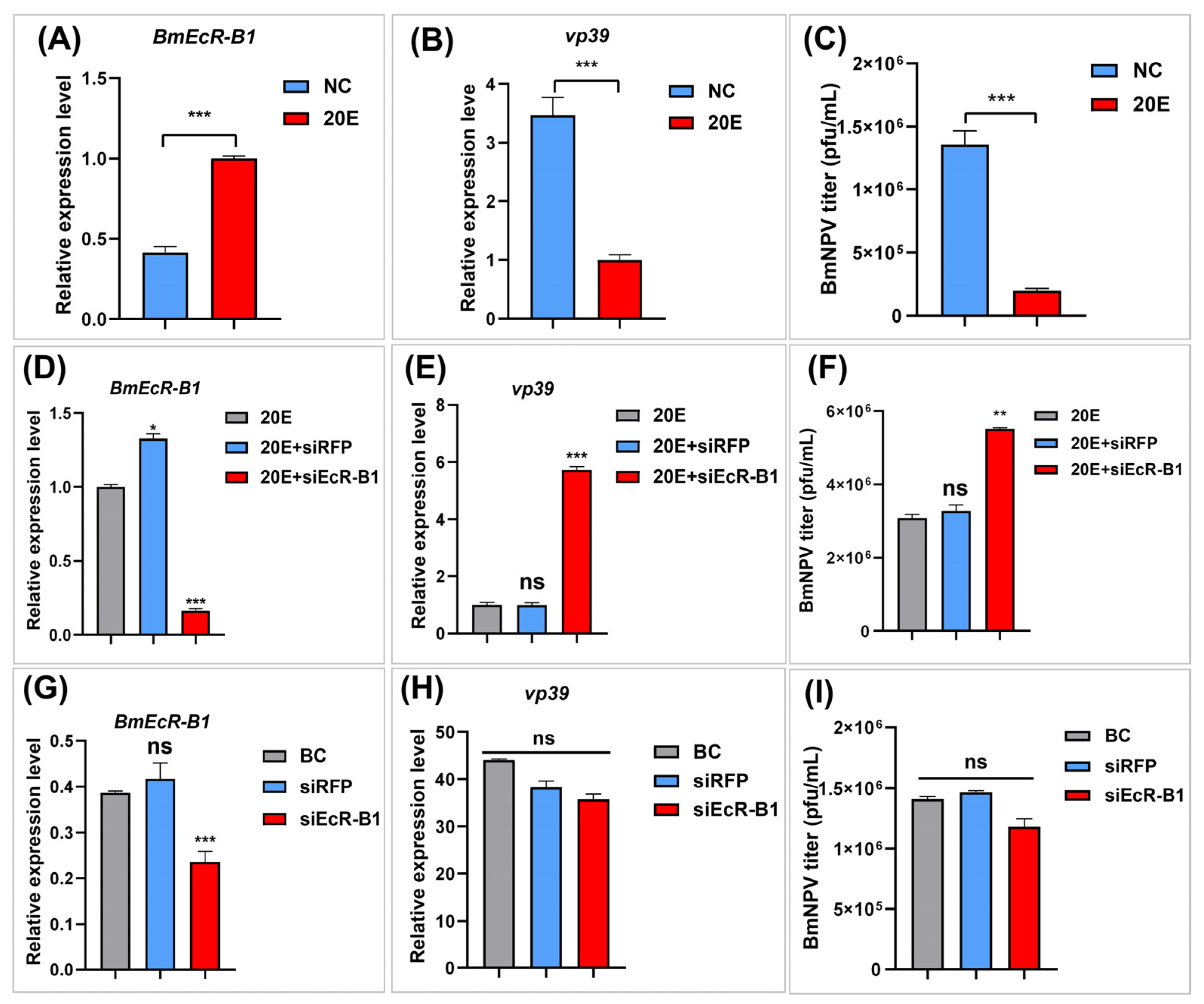
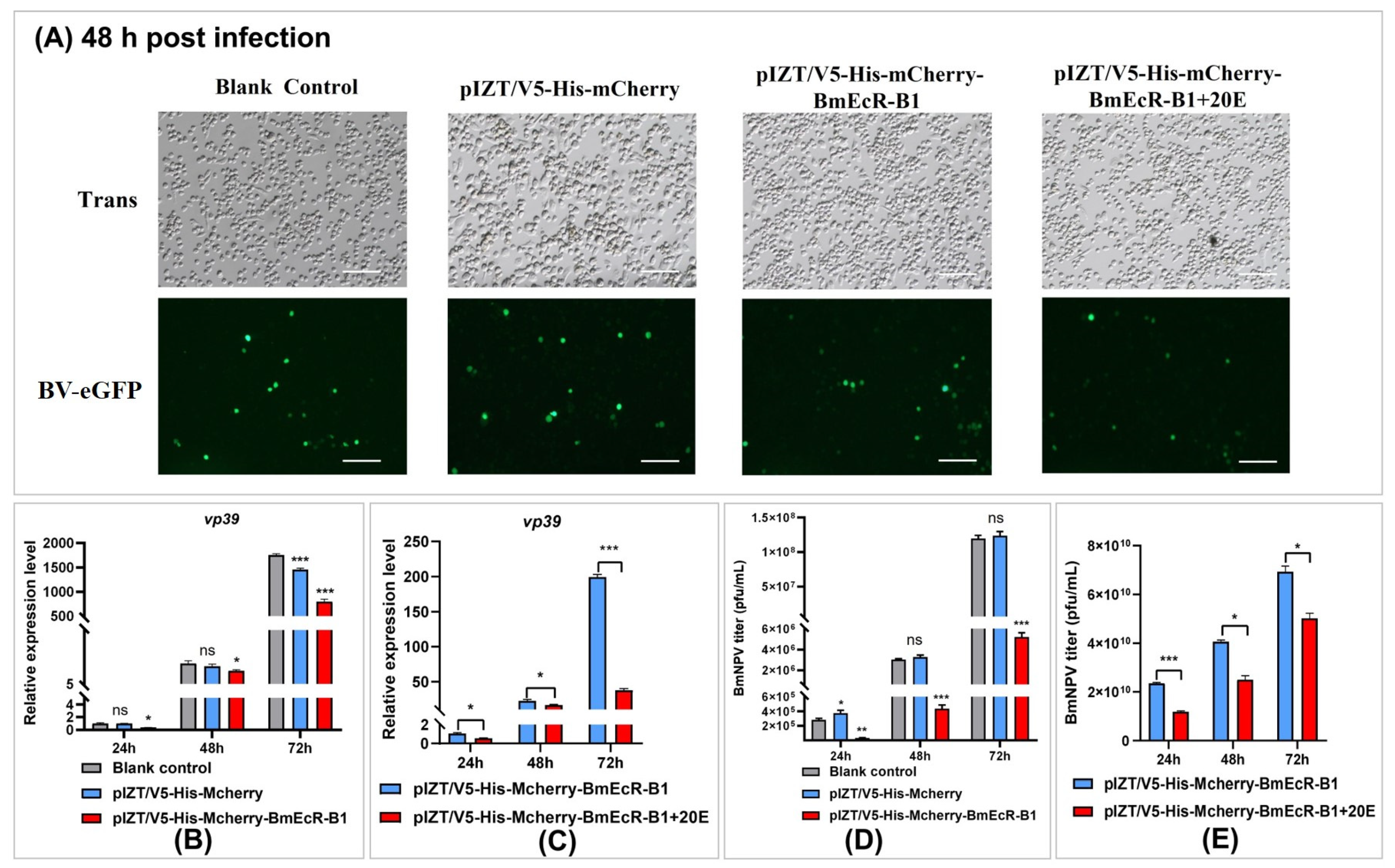
3.5. BmEcR-B1 Plays a Vital Role in 20E Inhibition of BmNPV In Vitro
3.6. Overexpression of BmEcR-B1 Inhibits BmNPV Infection in BmN Cells
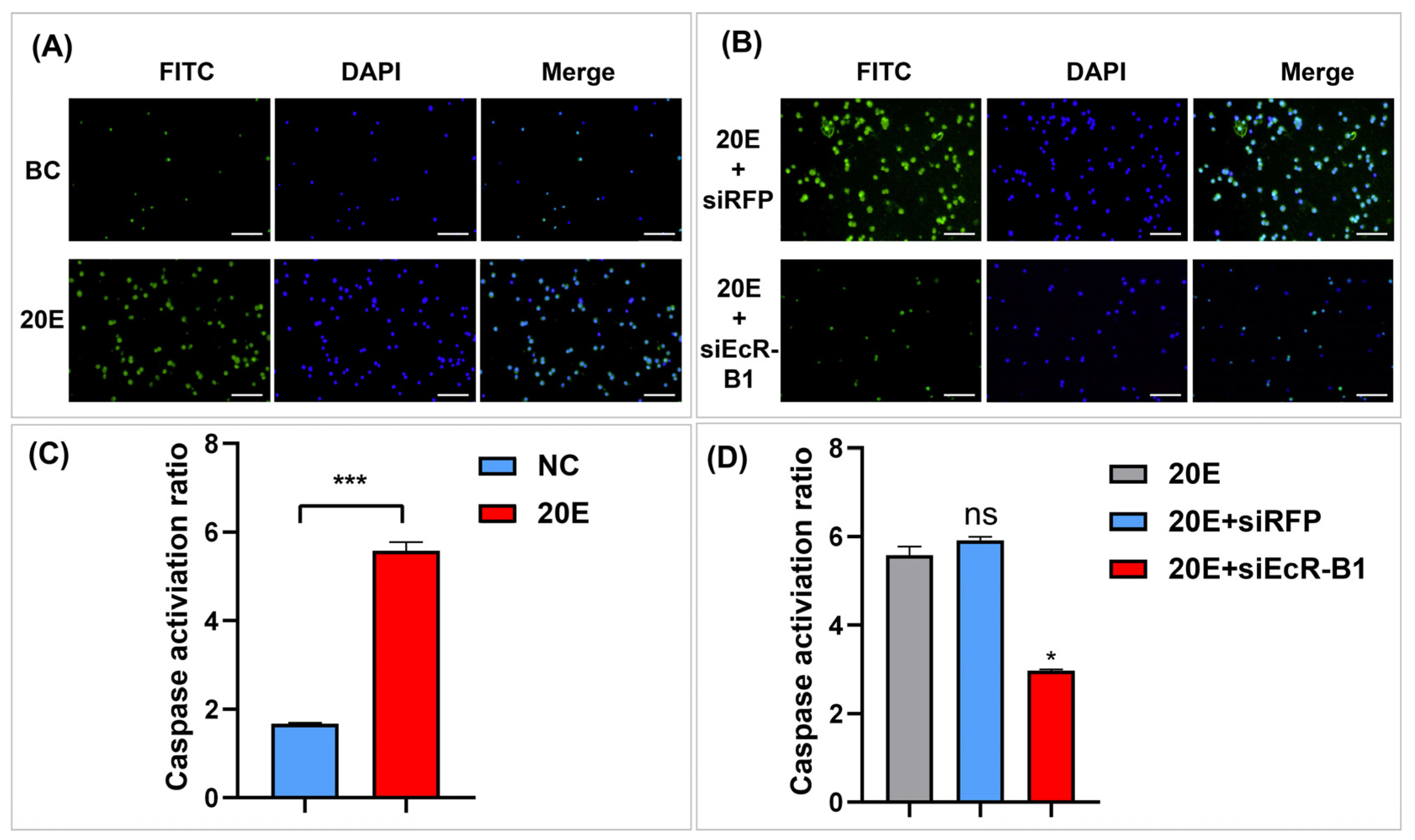
3.7. Inhibition of Viral Infection by BmEcR-B1 Is Associated with the Activation of Apoptosis
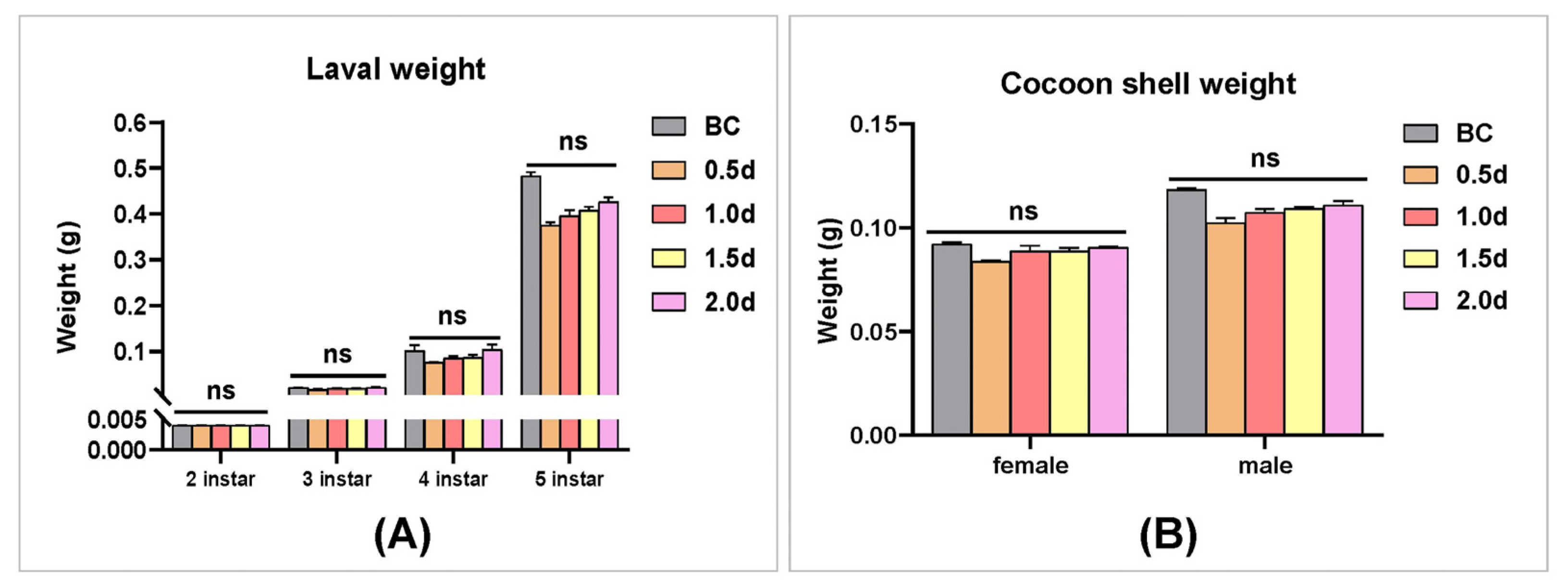
3.8. The Analysis of the Effect of 20E on Silkworm Larvae Development and Cocoon Shell
4. Discussion
4.1. BmEcR-B1 Is More Critical Than BmEcR-A in Silkworm
4.2. BmEcR-B1 Plays a Vital Role in Inhibiting BmNPV, Which Needs the Presence of 20E
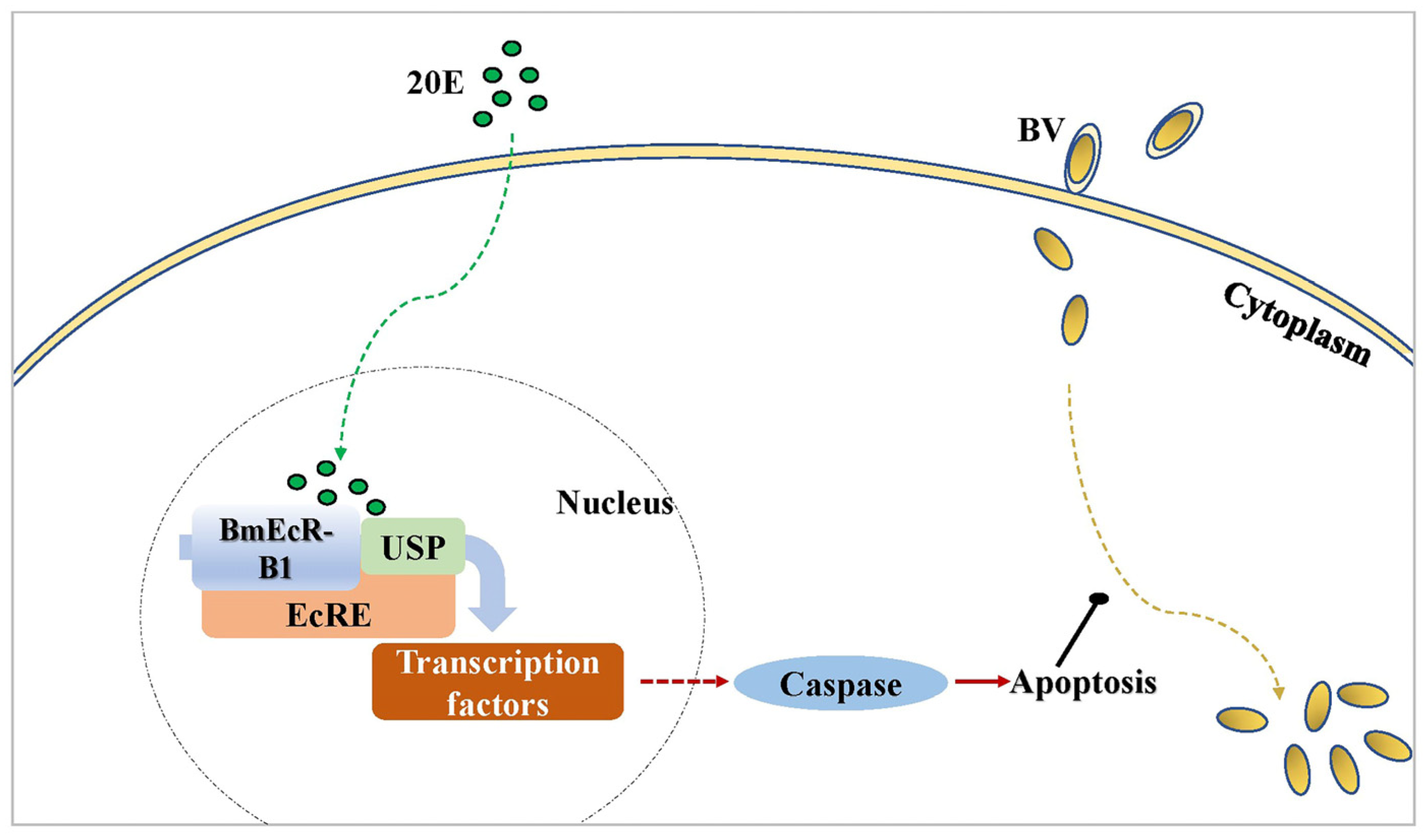
4.3. BmEcR-B1 Likely Inhibits Virus Infection in BmN Cells via the Activation of Apoptosis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jiang, L.; Goldsmith, M.R.; Xia, Q. Advances in the Arms Race Between Silkworm and Baculovirus. Front. Immunol. 2021, 12, 628151. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.Y.; Zhu, F.F.; Chen, K.P. The Mechanisms of Silkworm Resistance to the Baculovirus and Antiviral Breeding. Annu. Rev. Entomol. 2023, 68, 381–399. [Google Scholar] [CrossRef]
- Pan, J.; Di, Y.Q.; Li, Y.B.; Chen, C.H.; Wang, J.X.; Zhao, X.F. Insulin and 20-hydroxyecdysone oppose each other in the regulation of phosphoinositide-dependent kinase-1 expression during insect pupation. J. Biol. Chem. 2018, 293, 18613–18623. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.L.; Thummel, C.S. The RXR homolog ultraspiracle is an essential component of the Drosophila ecdysone receptor. Development 1998, 125, 4709–4717. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yao, B.; Li, X.; Jing, T.; Zhang, S.; Zou, H.; Zhang, G.; Zou, C. E74 knockdown represses larval development and chitin synthesis in Hyphantria cunea. Pestic. Biochem. Physiol. 2022, 187, 105216. [Google Scholar] [CrossRef]
- Xu, X.B.; Lai, Y.Y.; Hua, Z.C. Apoptosis and apoptotic body: Disease message and therapeutic target potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef]
- Ishikawa, H.; Ikeda, M.; Yanagimoto, K.; Alves, C.A.F.; Katou, Y.; Lavina-Caoili, B.A.; Kobayashi, M. Induction of apoptosis in an insect cell line, IPLB-Ld652Y, infected with nucleopolyhedroviruses. J. Gen. Virol. 2003, 84, 705–714. [Google Scholar] [CrossRef]
- Clarke, T.E.; Clem, R.J. In vivo induction of apoptosis correlating with reduced infectivity during baculovirus infection. J. Virol. 2003, 77, 2227–2232. [Google Scholar] [CrossRef]
- Xu, T.; Jiang, X.; Denton, D.; Kumar, S. Ecdysone controlled cell and tissue deletion. Cell. Death Differ. 2020, 27, 1–14. [Google Scholar] [CrossRef]
- Li, T.; Bender, M. A conditional rescue system reveals essential functions for the ecdysone receptor (EcR) gene during molting and metamorphosis in Drosophila. Development 2000, 127, 2897–2905. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Wang, S.; Li, S. Sumoylation modulates 20-hydroxyecdysone signaling by maintaining USP protein levels in Drosophila. Insect Biochem. Mol. Biol. 2014, 54, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Mansilla, A.; Martin, F.A.; Martin, D.; Ferrus, A. Ligand-independent requirements of steroid receptors EcR and USP for cell survival. Cell. Death Differ. 2016, 23, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Xu, S.; Dong, Y.Z.; Quan, L.F.; Chen, B.X. Ecdysone Receptor (EcR) and Ultraspiracle Protein (USP) Genes From Conopomorpha sinensisBradley Eggs: Identification and Expression in Response to Insecticides. Front. Physiol. 2020, 11, 851. [Google Scholar] [CrossRef]
- Kamimura, M.; Tomita, S.; Kiuchi, M.; Fujiwara, H. Tissue-specific and stage-specific expression of two silkworm ecdysone receptor isoforms—ecdysteroid-dependent transcription in cultured anterior silk glands. Eur. J. Biochem. 1997, 248, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Goncu, E.; Parlak, O. Morphological changes and patterns of ecdysone receptor B1 immunolocalization in the anterior silk gland undergoing programmed cell death in the silkworm, Bombyx mori. Acta Histochem. 2009, 111, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Sonobe, H.; Ito, Y. Phosphoconjugation and dephosphorylation reactions of steroid hormone in insects. Mol. Cell. Endocrinol. 2009, 307, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Shao, Z.M.; Chen, Q.Y.; Xu, J.P.; Sun, X.; Xu, Z.P.; Li, M.W.; Wu, Y.C. Knockdown of BmTCP-1 beta Delays BmNPV Infection in vitro. Front. Microbiol. 2019, 10, 578. [Google Scholar] [CrossRef]
- Guo, H.Z.; Jiang, L.; Xia, Q.Y. Selection of reference genes for analysis of stress-responsive genes after challenge with viruses and temperature changes in the silkworm Bombyx mori. Mol. Genet. Genom. 2016, 291, 999–1004. [Google Scholar] [CrossRef]
- Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.; Li, B.; Zhao, P.; Zha, X.; Cheng, T.; Chai, C.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar]
- Wang, M.; Wang, Y.; Chang, M.; Wang, X.; Shi, Z.; Raikhel, A.S.; Zou, Z. Ecdysone signaling mediates the trade-off between immunity and reproduction via suppression of amyloids in the mosquito Aedes aegypti. PLoS Pathog. 2022, 18, e1010837. [Google Scholar] [CrossRef]
- Wang, X.Y.; Yu, H.Z.; Geng, L.; Xu, J.P.; Yu, D.; Zhang, S.Z.; Ma, Y.; Fei, D.Q. Comparative Transcriptome Analysis of Bombyx mori (Lepidoptera) Larval Midgut Response to BmNPV in Susceptible and Near-Isogenic Resistant Strains. PLoS ONE 2016, 11, e0155341. [Google Scholar] [CrossRef]
- Nunes, C.; Sucena, E.; Koyama, T. Endocrine regulation of immunity in insects. FEBS J. 2021, 288, 3928–3947. [Google Scholar] [CrossRef] [PubMed]
- Bender, M.; Imam, F.B.; Talbot, W.S.; Ganetzky, B.; Hogness, D.S. Drosophila ecdysone receptor mutations reveal functional differences among receptor isoforms. Cell 1997, 91, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Truman, J.W. The evolution of insect metamorphosis. Curr. Biol. 2019, 29, R1252–R1268. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, S. 20-hydroxyecdysone enhances the expression of the chitinase 5 via Broad-Complex Zinc-Finger 4 during metamorphosis in silkworm, Bombyx mori. Insect Mol. Biol. 2017, 26, 243–253. [Google Scholar] [CrossRef]
- Sun, W.; Shen, Y.H.; Zhou, L.X.; Zhang, Z. Ecdysone Titer determined by 3DE-3 beta-reductase enhances the immune response in the silkworm. J. Immunol. 2016, 196, 1646–1654. [Google Scholar] [CrossRef]
- Cheng, Y.B.; Lee, J.C.; Lo, I.W.; Chen, S.R.; Hu, H.C.; Wu, Y.H.; Wu, Y.C.; Chang, F.R. Ecdysones from Zoanthus spp. with inhibitory activity against dengue virus 2. Bioorganic Med. Chem. Lett. 2016, 26, 2344–2348. [Google Scholar] [CrossRef] [PubMed]
- Jayachandran, B.; Hussain, M.; Asgari, S. Regulation of Helicoverpa armigera ecdysone receptor by miR-14 and its potential link to baculovirus infection. J. Invertebr. Pathol. 2013, 114, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.B.; Li, X.R.; Yang, T.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone promotes switching from autophagy to apoptosis by increasing intracellular calcium levels. Insect Biochem. Mol. Biol. 2016, 79, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.B.; Pei, X.Y.; Wang, D.; Chen, C.H.; Cai, M.J.; Wang, J.X.; Zhao, X.F. The steroid hormone 20-hydroxyecdysone upregulates calcium release-activated calcium channel modulator 1 expression to induce apoptosis in the midgut of Helicoverpa armigera. Cell Calcium. 2017, 68, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pei, X.Y.; Zhao, W.L.; Zhao, X.F. Steroid hormone 20-hydroxyecdysone promotes higher calcium mobilization to induce apoptosis. Cell Calcium. 2016, 60, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Song, H.Y.; Kim, J.H.; Kim, B.Y.; Park, S.W.; Sung, D.K.; Park, H.H.; Jeon, S.H.; Chung, I.S.; Lee, B.H. Silkworm 30k protein inhibits ecdysone-induced apoptosis by blocking the binding of ultraspiracle to ecdysone receptor-b1 in cultured Bm5 cells. Arch. Insect Biochem. 2012, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Note |
|---|---|---|---|
| BmEcR-B1-O | CGGAATTCATGGACAGAGCCTCCGGATAC | GCTCTAGAGATGCACATGTTGGAGTTTTG | ORF amplification |
| BmEcR-B1 | GCCTCCCACAACACCGAAATCA | TCTGGCGTCAGCATCAGCACT | 98.30%, 106 bp |
| BmEcR-A | ACTCGTCGCCACTATCCTCAGG | TGAAGCCGGCGAAAGTTCCT | 98.48%, 110 bp |
| BmGAPDH | CCGCGTCCCTGTTGCTAAT | CTGCCTCCTTGACCTTTTGC | 116.00%, 98 bp |
| vp39 | CAACTTTTTGCGAAACGACTT | GGCTACACCTCCACTTGCTT | 100.71%, 125 bp |
| Primer Names | Sequences (5′-3′) |
|---|---|
| EcR-Olig1-1 | GATCACTAATACGACTCACTATAGGGAATTTGGACATGCCTGTGAAATT |
| EcR-Olig1-2 | AATTTCACAGGCATGTCCAAATTCCCTATAGTGAGTCGTATTAGTGATC |
| EcR-Olig1-3 | AAAATTTGGACATGCCTGTGAAACCCTATAGTGAGTCGTATTAGTGATC |
| EcR-Olig1-4 | GATCACTAATACGACTCACTATAGGGTTTCACAGGCATGTCCAAATTTT |
| EcR-Olig2-1 | GATCACTAATACGACTCACTATAGGGAACACGTTGCGAATTTACATCTT |
| EcR-Olig2-2 | AAGATGTAAATTCGCAACGTGTTCCCTATAGTGAGTCGTATTAGTGATC |
| EcR-Olig2-3 | AAAACACGTTGCGAATTTACATCCCCTATAGTGAGTCGTATTAGTGATC |
| EcR-Olig2-4 | GATCACTAATACGACTCACTATAGGGGATGTAAATTCGCAACGTGTTTT |
| RFP-Olig-1 | GATCACTAATACGACTCACTATAGGGGCACCCAGACCATGAGAATTT |
| RFP-Olig-2 | AAATTCTCATGGTCTGGGTGCCCCTATAGTGAGTCGTATTAGTGATC |
| RFP-Olig-3 | AAGCACCCAGACCATGAGAATCCCTATAGTGAGTCGTATTAGTGATC |
| RFP-Olig-4 | GATCACTAATACGACTCACTATAGGGATTCTCATGGTCTGGGTGCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Z.; Zhao, C.; Huang, X.; Lv, J.; Zhao, Z.; Zheng, K.; Sun, X.; Qin, S.; Wang, X.; Jin, B.-R.; et al. Bombyx mori Ecdysone Receptor B1 May Inhibit BmNPV Infection by Triggering Apoptosis. Insects 2023, 14, 505. https://doi.org/10.3390/insects14060505
Su Z, Zhao C, Huang X, Lv J, Zhao Z, Zheng K, Sun X, Qin S, Wang X, Jin B-R, et al. Bombyx mori Ecdysone Receptor B1 May Inhibit BmNPV Infection by Triggering Apoptosis. Insects. 2023; 14(6):505. https://doi.org/10.3390/insects14060505
Chicago/Turabian StyleSu, Zhihao, Chunxiao Zhao, Xinming Huang, Junli Lv, Ziqin Zhao, Kaiyi Zheng, Xia Sun, Sheng Qin, Xueyang Wang, Byung-Rae Jin, and et al. 2023. "Bombyx mori Ecdysone Receptor B1 May Inhibit BmNPV Infection by Triggering Apoptosis" Insects 14, no. 6: 505. https://doi.org/10.3390/insects14060505
APA StyleSu, Z., Zhao, C., Huang, X., Lv, J., Zhao, Z., Zheng, K., Sun, X., Qin, S., Wang, X., Jin, B.-R., & Wu, Y. (2023). Bombyx mori Ecdysone Receptor B1 May Inhibit BmNPV Infection by Triggering Apoptosis. Insects, 14(6), 505. https://doi.org/10.3390/insects14060505