
A Survey of the Dung-Dwelling Arthropod Community in the Pastures of the Northern Plains


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling Procedure
2.3. Community Composition
2.4. Data Analysis
3. Results
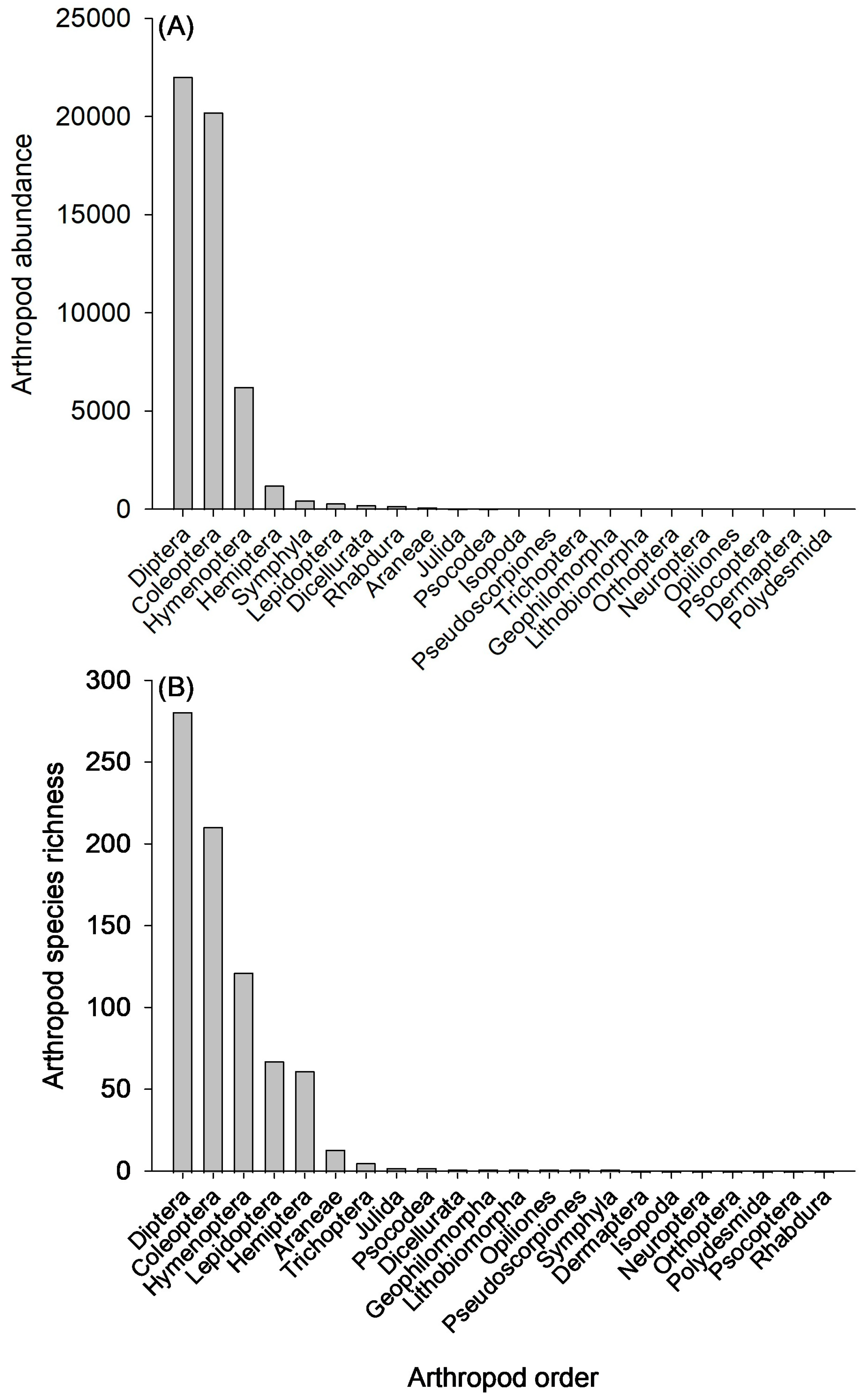
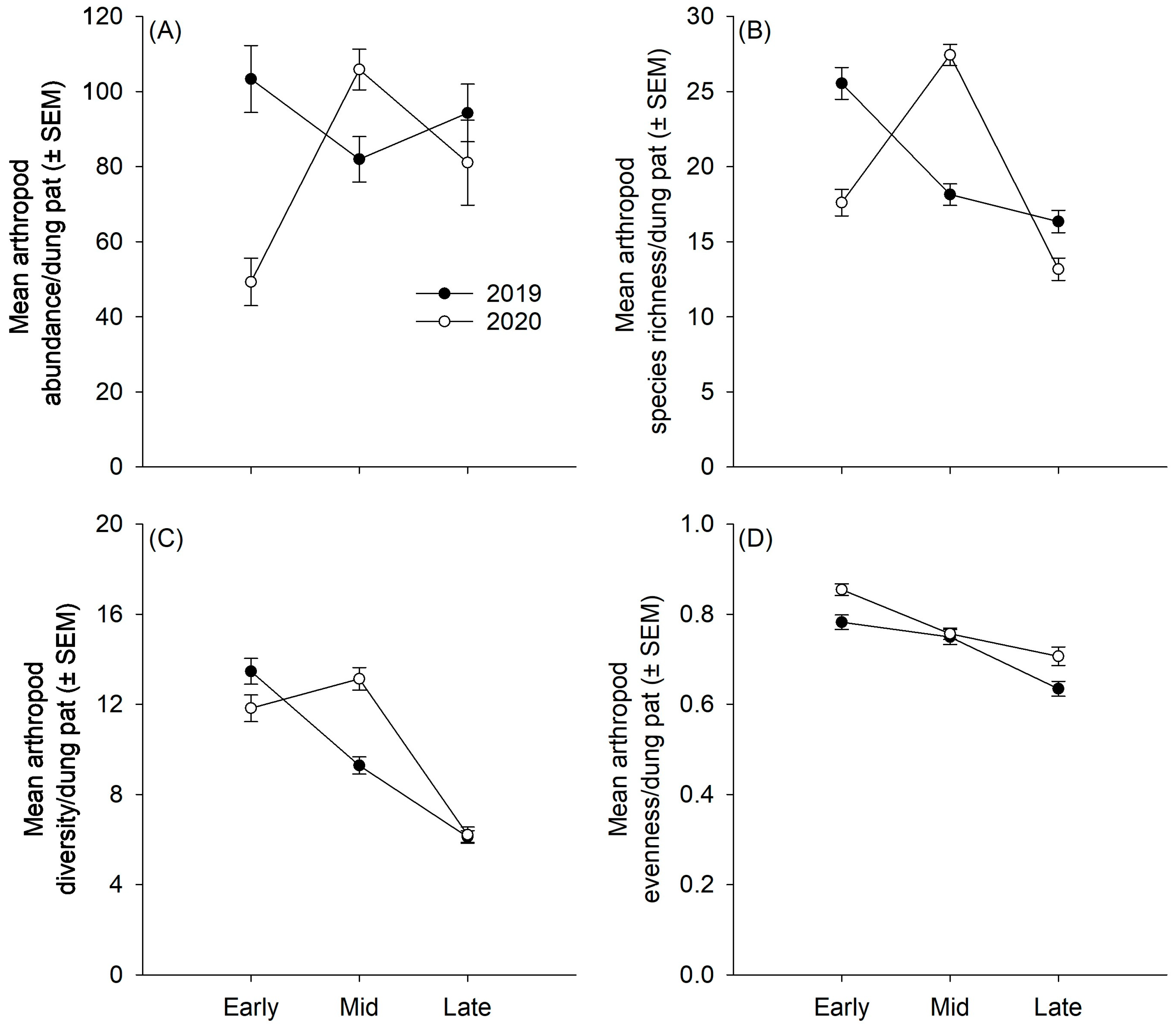
3.1. Arthropod Community
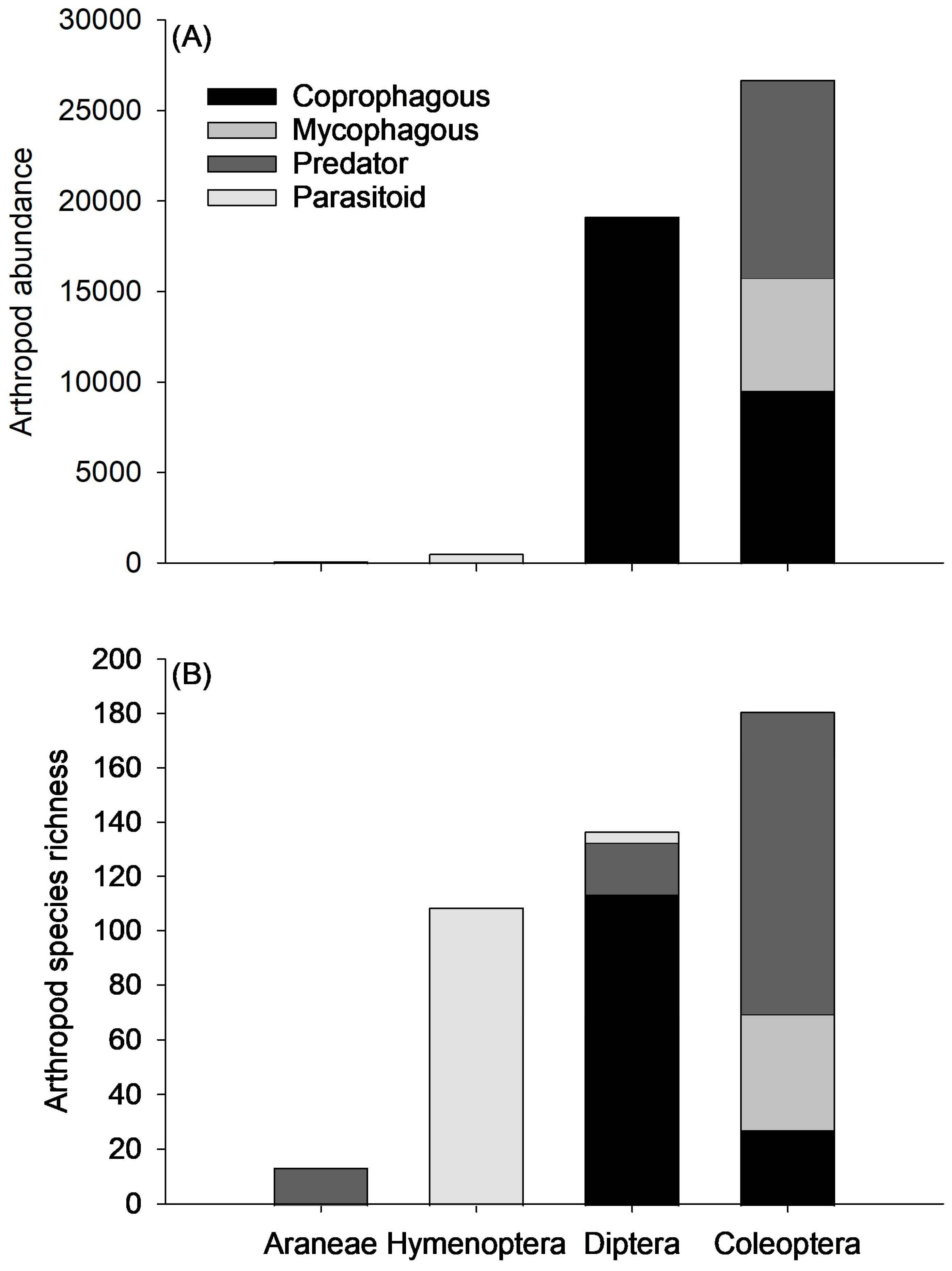
3.2. Functional Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartnett, D.C.; Steuter, A.A.; Hickman, K.R. Comparative Ecology of Native and Introduced Ungulates. In Ecology and Conservation of Great Plains Vertebrates; Volume Ecological; Springer: New York, NY, USA, 1997; pp. 77–101. [Google Scholar]
- Pokhrel, M.R.; Cairns, S.C.; Hemmings, Z.; Floate, K.D.; Andrew, N.R. A Review of Dung Beetle Introductions in the Antipodes and North America: Status, Opportunities, and Challenges. Environ. Entomol. 2021, 50, 762–780. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, Y.; Bouchard, P.; Davies, A.; Sikes, D. Checklist of Beetles (Coleoptera) of Canada and Alaska. Second Edition. ZooKeys 2013, 360, 1–44. [Google Scholar] [CrossRef]
- Bezanson, G.A.; Floate, K.D. An Updated Checklist of the Coleoptera Associated with Livestock Dung on Pastures in America North of Mexico. Coleopt. Bull. 2019, 73, 655–683. [Google Scholar] [CrossRef]
- Merritt, R.W.; Anderson, J.R. The Effects of Different Pasture and Rangeland Ecosystems on the Annual Dynamics of Insects in Cattle Droppings. Univ. Calif. Div. Agric. Sci. 1977, 45, 31–70. [Google Scholar] [CrossRef]
- Floate, K.D. Arthropods in Cattle Dung on Canada’s Grasslands. Arthropods Can. Grassl. 2011, 2, 71–88. [Google Scholar]
- Giménez Gómez, V.C.; Verdú, J.R.; Guerra Alonso, C.B.; Zurita, G.A. Relationship between Land Uses and Diversity of Dung Beetles (Coleoptera: Scarabaeinae) in the Southern Atlantic Forest of Argentina: Which Are the Key Factors? Biodivers. Conserv. 2018, 27, 3201–3213. [Google Scholar] [CrossRef]
- Pecenka, J.R.; Lundgren, J.G. Effects of Herd Management and the Use of Ivermectin on Dung Arthropod Communities in Grasslands. Basic Appl. Ecol. 2019, 40, 19–29. [Google Scholar] [CrossRef]
- Perrin, W.; Moretti, M.; Vergnes, A.; Borcard, D.; Jay-Robert, P. Response of Dung Beetle Assemblages to Grazing Intensity in Two Distinct Bioclimatic Contexts. Agric. Ecosyst. Environ. 2020, 289, 106740. [Google Scholar] [CrossRef]
- Pecenka, J.R.; Lundgren, J.G. The Importance of Dung Beetles and Arthropod Communities on Degradation of Cattle Dung Pats in Eastern South Dakota. PeerJ 2018, 6, e5220. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, M. Some Considerations on the Terminology Applied to Dung Beetle Functional Groups. Ecol. Entomol. 2021, 46, 772–776. [Google Scholar] [CrossRef]
- Zunino, M.; Barbero, E. Food Relocation and the Reproductive Biology of Aphodius fossor (L.) (Coleoptera Scarabaeidae Aphodiinae). Ethol. Ecol. Evol. 1990, 2, 334. [Google Scholar] [CrossRef]
- Gittings, T.; Giller, P.S. Life History Traits and Resource Utilisation in an Assemblage of North Temperate Aphodius Dung Beetles (Coleoptera: Scarabaeidae). Ecography 1997, 20, 55–66. [Google Scholar] [CrossRef]
- Hanski, I. Nutritional Ecology of Dung- and Carrion-Feeding Insects. In Nutritional Ecology of Insects, Mites, and Spiders; Wiley: New York, NY, USA, 1987; pp. 837–884. [Google Scholar]
- Wratten, S.D.; Forbes, A.B. Environmental Assessment of Veterinary Avermectins in Temperate Pastoral Ecosystems. Ann. Appl. Biol. 1996, 128, 329–348. [Google Scholar] [CrossRef]
- Stevenson, B.; Dindal, D. Insect Effects on Decomposition of Cow Dung in Microcosms. Pedobiologia 1987, 30, 81–92. [Google Scholar] [CrossRef]
- Hanski, I.; Cambefort, Y. Resource Partitioning. In Dung Beetle Ecology; Princeton University Press: Princeton, NJ, USA, 1991; pp. 330–349. [Google Scholar]
- Jargalsaikhan, P.; Altangerel, G.; Enkhchimeg, T.; Aibek, U.; Bayartogtokh, B. Variation in Dung Removal Rates by Dung Beetles (Coleoptera: Scarabaeoidea) in a Temperate, Dry Steppe Ecosystem. Diversity 2023, 15, 91. [Google Scholar] [CrossRef]
- Huerta, C.; Martínez, M.I.; de Oca, E.M.; Cruz-Rosales, M.; Favila, M.E. The Role of Dung Beetles in the Sustainability of Pasture and Grasslands. In WIT Transactions on State of the Art in Science and Engineering; Yáñez-Arancibia, A., Dávalos-Sotelo, R., Day, J.W., Reyes, E., Eds.; WIT Press: Southampton, UK, 2013; Volume 1, pp. 441–463. ISBN 978-1-84564-756-8. [Google Scholar]
- Nervo, B.; Tocco, C.; Caprio, E.; Palestrini, C.; Rolando, A. The Effects of Body Mass on Dung Removal Efficiency in Dung Beetles. PLoS ONE 2014, 9, e107699. [Google Scholar] [CrossRef] [PubMed]
- Wolcott, G.N. Insect Parasite Introduction in Porto Rico. JAUPR 1922, 6, 5–20. [Google Scholar] [CrossRef]
- Stanbrook, R.; King, J.R. Dung Beetle Community Composition Affects Dung Turnover in Subtropical US Grasslands. Ecol. Evol. 2022, 12, e8660. [Google Scholar] [CrossRef] [PubMed]
- Hanski, I. North Temperate Dung Beetles. In Dung Beetle Ecology; Princeton University Press: Princeton, NJ, USA, 1991; pp. 75–96. [Google Scholar]
- Kohlmann, B. Dung Beetles in Subtropical North America. In Dung Beetle Ecology; Princeton University Press: Princeton, NJ, USA, 1991; pp. 116–132. [Google Scholar]
- Claassen, R.; Cooper, J.C.; Carriazo, F. Crop Insurance, Disaster Payments, and Land Use Change: The Effect of Sodsaver on Incentives for Grassland Conversion. J. Agric. Appl. Econ. 2011, 43, 195–211. [Google Scholar] [CrossRef]
- Wimberly, M.C.; Janssen, L.L.; Hennessy, D.A.; Luri, M.; Chowdhury, N.M.; Feng, H. Cropland Expansion and Grassland Loss in the Eastern Dakotas: New Insights from a Farm-Level Survey. Land Use Policy 2017, 63, 160–173. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Season | Species Richness | Exponential Shannon-Wiener | Inverse Simpson | Inverse Berger-Parker |
|---|---|---|---|---|---|
| 2019 | Early | 25.5 ± 1.1 | 13.4 ± 0.6 | 10.7 ± 0.6 | 4.4 ± 0.2 |
| Mid | 18.1 ± 0.7 | 9.3 ± 0.4 | 7.3 ± 0.4 | 3.2 ± 0.1 | |
| Late | 16.3 ± 0.7 | 6.1 ± 0.3 | 3.8 ± 0.2 | 2.1 ± 0.1 | |
| 2020 | Early | 17.6 ± 0.9 | 11.8 ± 0.6 | 15.5 ± 1.8 | 4.4 ± 0.2 |
| Mid | 27.4 ± 0.7 | 13.1 ± 0.5 | 9.2 ± 0.6 | 3.9 ± 1.2 | |
| Late | 13.2 ± 0.7 | 6.2 ± 0.3 | 5.3 ± 0.5 | 2.7 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmid, R.B.; Welch, K.D.; Lundgren, J.G. A Survey of the Dung-Dwelling Arthropod Community in the Pastures of the Northern Plains. Insects 2024, 15, 38. https://doi.org/10.3390/insects15010038
Schmid RB, Welch KD, Lundgren JG. A Survey of the Dung-Dwelling Arthropod Community in the Pastures of the Northern Plains. Insects. 2024; 15(1):38. https://doi.org/10.3390/insects15010038
Chicago/Turabian StyleSchmid, Ryan B., Kelton D. Welch, and Jonathan G. Lundgren. 2024. "A Survey of the Dung-Dwelling Arthropod Community in the Pastures of the Northern Plains" Insects 15, no. 1: 38. https://doi.org/10.3390/insects15010038
APA StyleSchmid, R. B., Welch, K. D., & Lundgren, J. G. (2024). A Survey of the Dung-Dwelling Arthropod Community in the Pastures of the Northern Plains. Insects, 15(1), 38. https://doi.org/10.3390/insects15010038