
A Host Tree and Its Specialist Insects: Black Locust (Robinia pseudoacacia) Availability Largely Determines the Future Range Dynamics of Its Specialist Insects in Europe


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
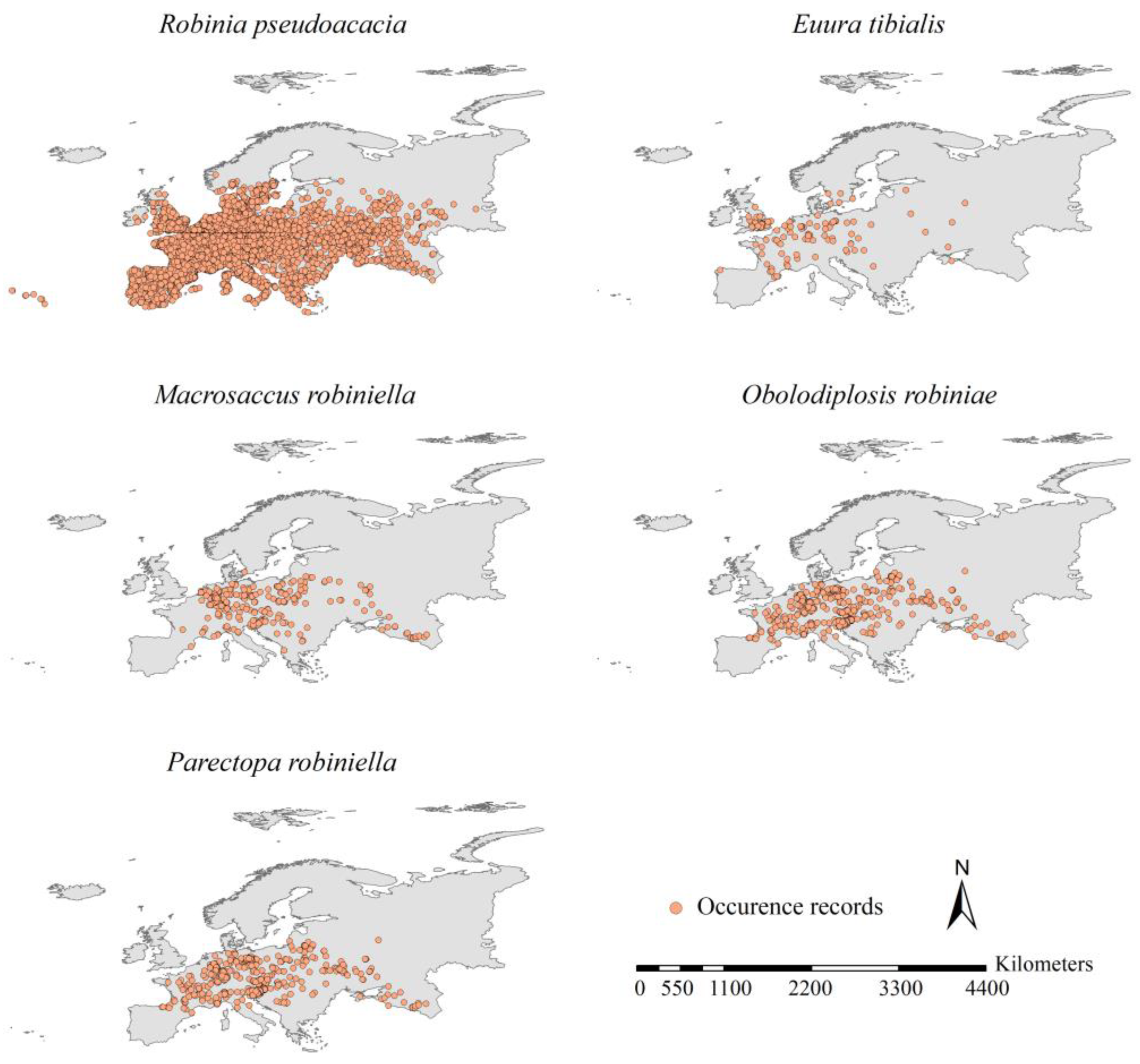
2.1. Retrieving Occurrence Records
2.2. Preparing Predictors
2.3. Predictor Selection
2.4. SDM Development
2.5. Range Dynamics
2.6. Statistical Analysis
3. Results
3.1. Model Reliability
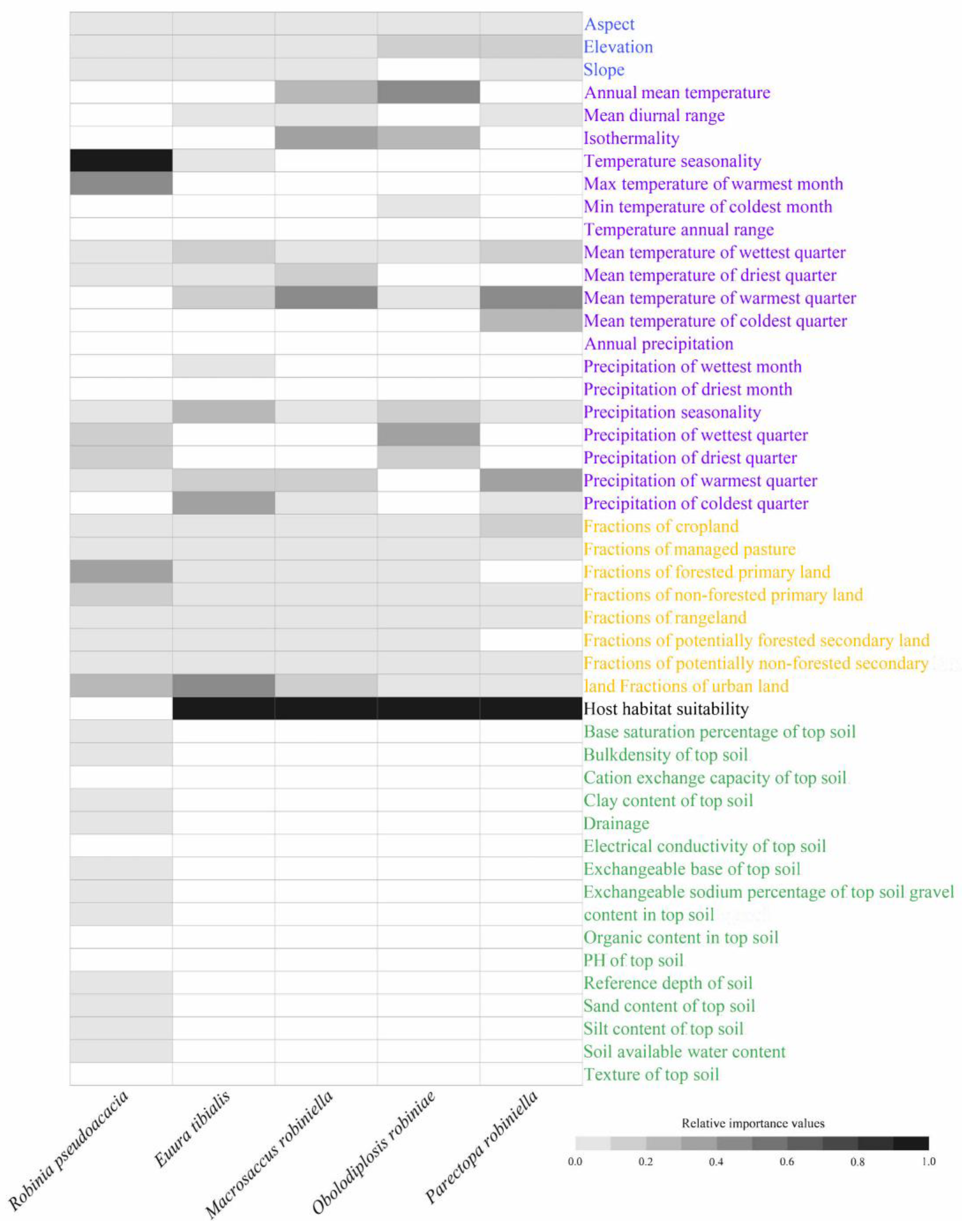
3.2. Predictors Affecting Range Dynamics
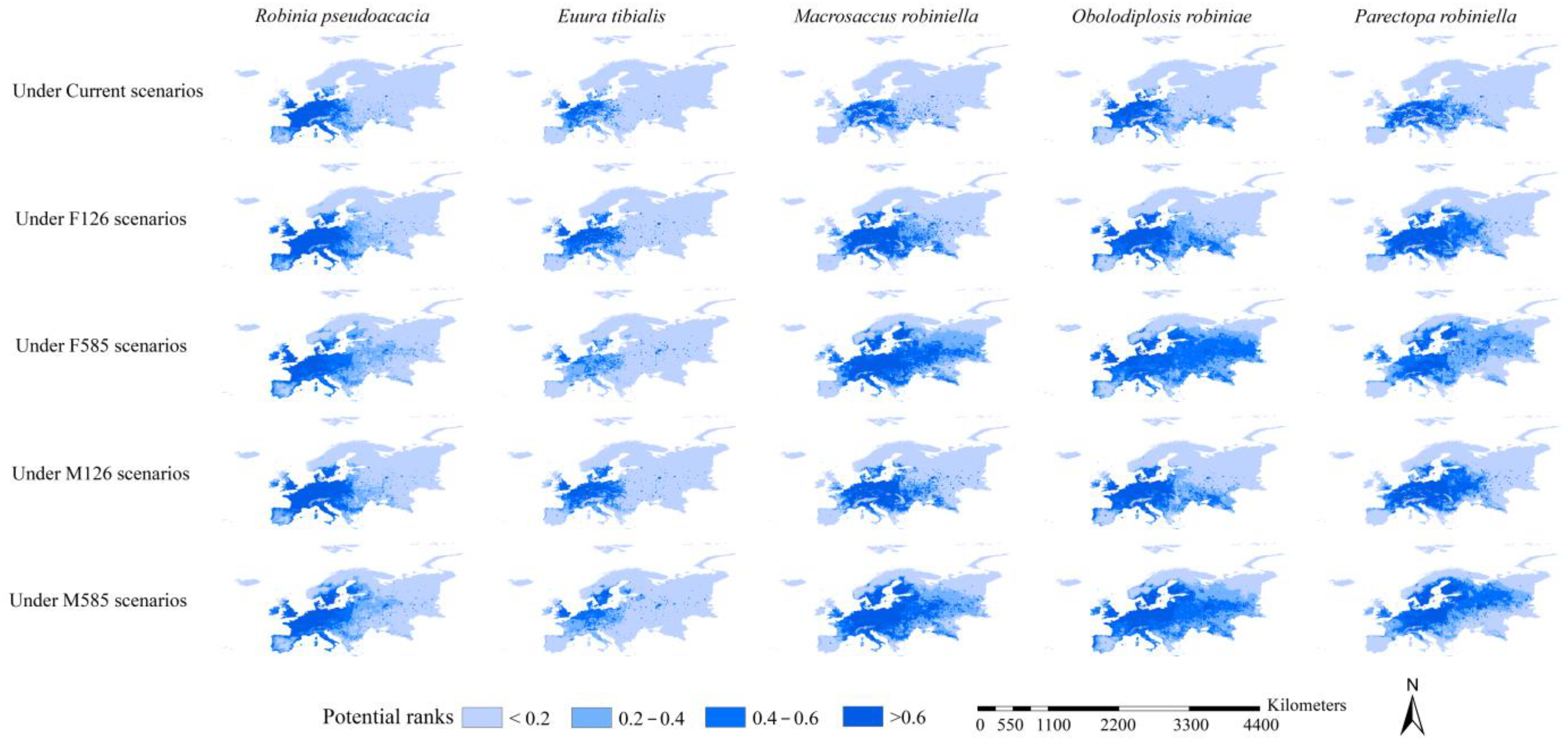
3.3. Habitat Suitability of Black Locust and Its Specialist Insects
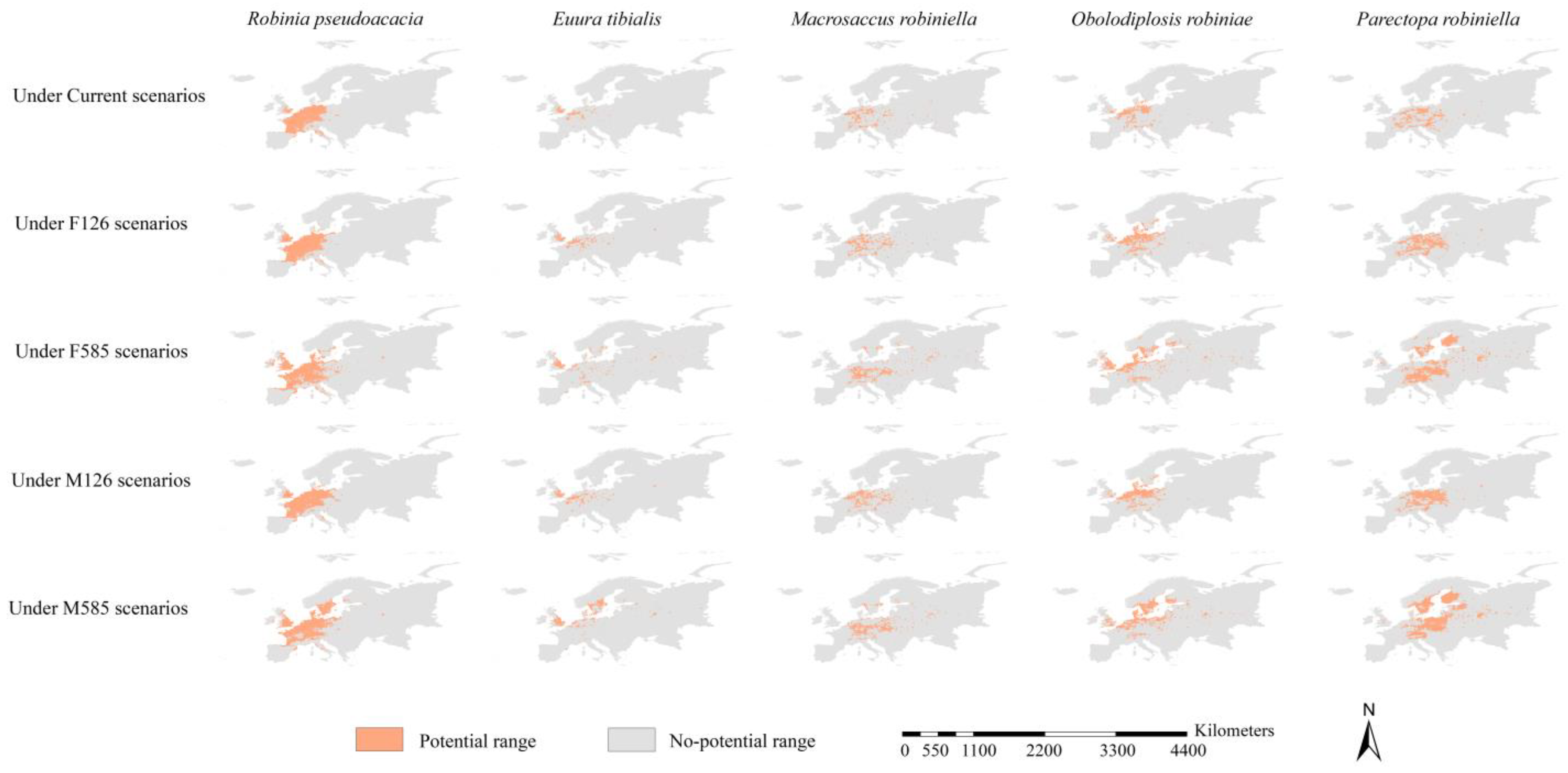
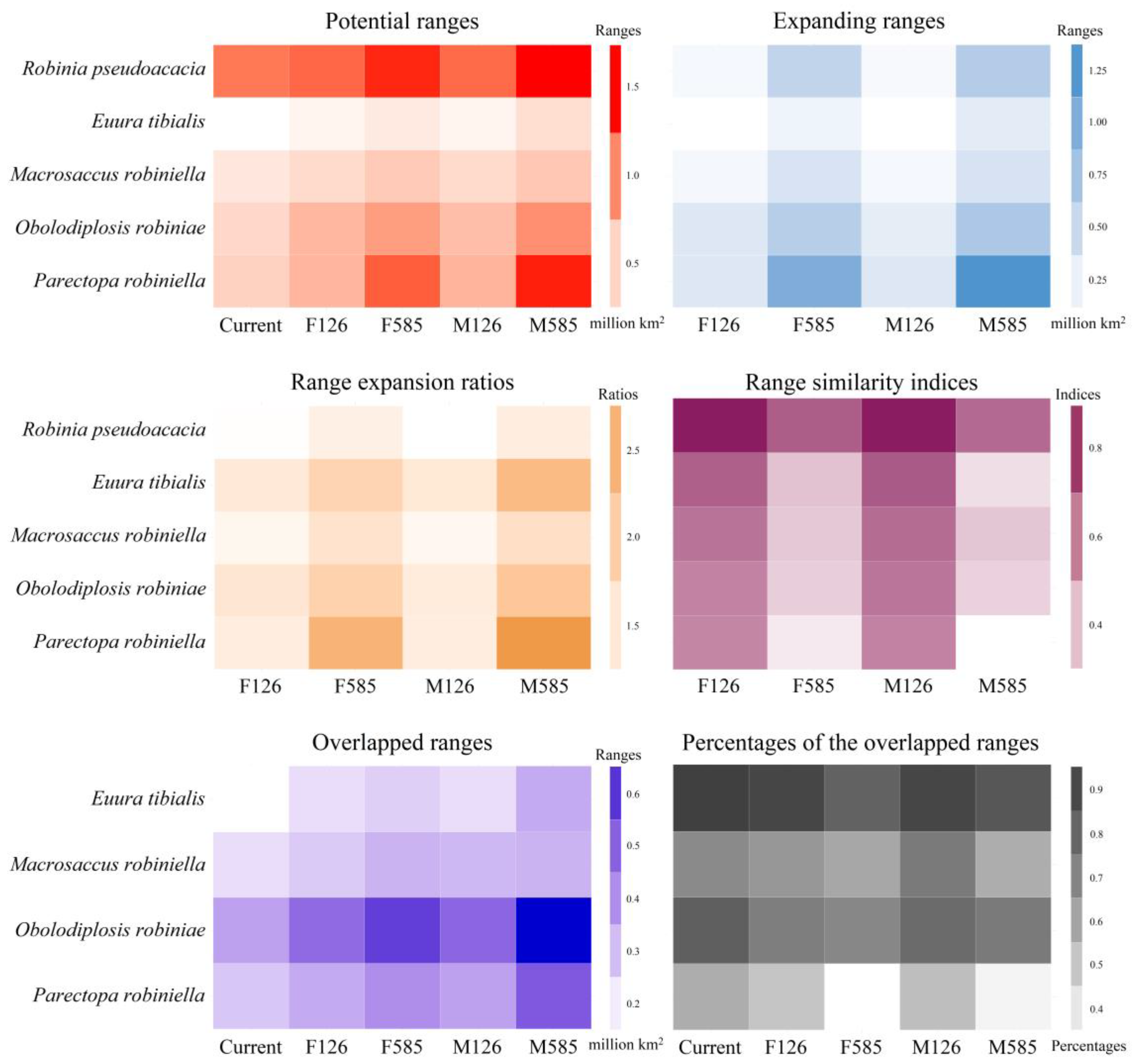
3.4. Potential Ranges of Black Locust and Its Specialist Insects
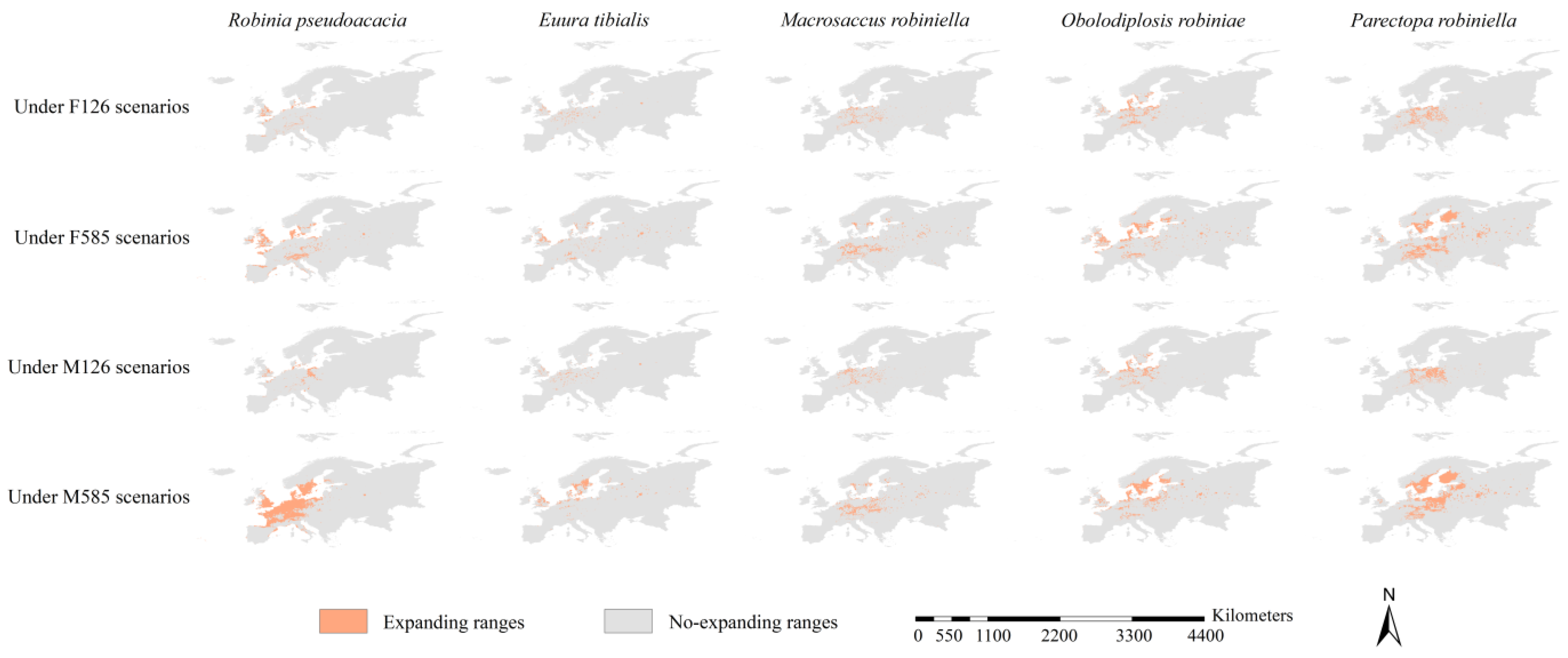
3.5. Range Dynamics of the Black Locust and Its Specialist Insects
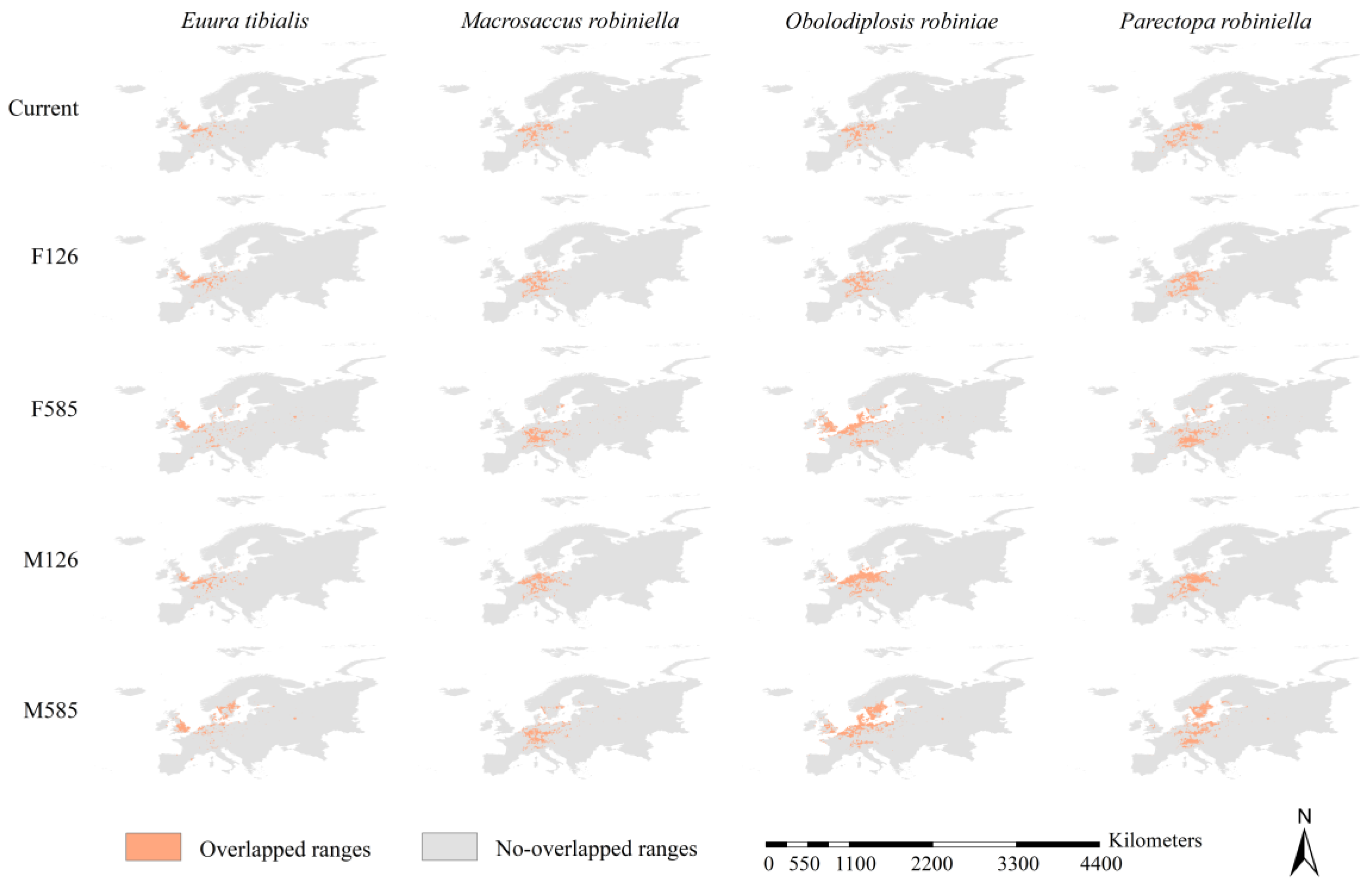
3.6. Range Overlap between Black Locust and Its Specialist Insects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, V.-N.; Hernea, C.; Bakti, B.; Keserű, Z.; Antal, B.; Rédei, K. Black locust (Robinia pseudoacacia L.) as a multi-purpose tree species in Hungary and Romania: A review. J. For. Res. 2018, 29, 1449–1463. [Google Scholar] [CrossRef]
- Nicolescu, V.N.; Rédei, K.; Mason, W.L.; Vor, T.; Pöetzelsberger, E.; Bastien, J.C.; Brus, R.; Bencat, T.; Dodan, M.; Cvjetkovic, B.; et al. Ecology, growth and management of black locust (Robinia pseudoacacia L.), a non-native species integrated into European forests. J. For. Res. 2020, 31, 1081–1101. [Google Scholar] [CrossRef]
- Li, G.; Xu, G.; Guo, K.; Du, S. Mapping the global potential geographical distribution of black locust (Robinia pseudoacacia L.) using herbarium data and a maximum entropy model. Forests 2014, 5, 2773–2792. [Google Scholar] [CrossRef]
- Martin, G.D. Addressing geographical bias: A review of Robinia pseudoacacia (black locust) in the southern hemisphere. S. Afr. J. Bot. 2019, 125, 481–490. [Google Scholar] [CrossRef]
- Benesperi, R.; Giuliani, C.; Zanetti, S.; Gennai, M.; Lippi, M.M.; Guidi, T.; Nascimbene, J.; Foggi, B. Forest plant diversity is threatened by Robinia pseudoacacia (black-locust) invasion. Biodivers. Conserv. 2012, 21, 3555–3568. [Google Scholar] [CrossRef]
- Rumlerová, Z.; Vilà, M.; Pergl, J.; Nentwig, W.; Pyšek, P. Scoring environmental and socioeconomic impacts of alien plants invasive in Europe. Biol. Invasions 2016, 18, 3697–3711. [Google Scholar] [CrossRef]
- Kroftová, M.; Reif, J. Management implications of bird responses to variation in non-native/native tree ratios within central European forest stands. For. Ecol. Manag. 2017, 391, 330–337. [Google Scholar] [CrossRef]
- Lazzaro, L.; Mazza, G.; d’Errico, G.; Fabiani, A.; Giuliani, C.; Inghilesi, A.F.; Lagomarsino, A.; Landi, S.; Lastrucci, L.; Pastorelli, R.; et al. How ecosystems change following invasion by Robinia pseudoacacia: Insights from soil chemical properties and soil microbial, nematode, microarthropod and plant communities. Sci. Total Environ. 2018, 622, 1509–1518. [Google Scholar] [CrossRef]
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2018, 20, 1611–1621. [Google Scholar] [CrossRef]
- Nascimbene, J.; Benesperi, R.; Casazza, G.; Chiarucci, A.; Giordani, P. Range shifts of native and invasive trees exacerbate the impact of climate change on epiphyte distribution: The case of lung lichen and black locust in Italy. Sci. Total Environ. 2020, 735, 139537. [Google Scholar] [CrossRef]
- Mally, R.; Ward, S.F.; Trombik, J.; Buszko, J.; Medzihorsky, V.; Liebhold, A.M. Non-native plant drives the spatial dynamics of its herbivores: The case of black locust (Robinia pseudoacacia) in Europe. NeoBiota 2021, 69, 155–175. [Google Scholar] [CrossRef]
- Puchałka, R.; Dyderski, M.K.; Vítková, M.; Sádlo, J.; Klisz, M.; Netsvetov, M.; Prokopuk, Y.; Matisons, R.; Mionskowski, M.; Wojda, T.; et al. Black locust (Robinia pseudoacacia L.) range contraction and expansion in Europe under changing climate. Glob. Chang. Biol. 2021, 27, 1587–1600. [Google Scholar] [CrossRef] [PubMed]
- Tytar, V.; Nekrasova, O.; Marushchak, O.; Pupins, M.; Skute, A.; Čeirāns, A.; Kozynenko, I. The spread of the invasive locust digitate leafminer Parectopa robiniella Clemens, 1863 (Lepidoptera: Gracillariidae) in Europe, with special reference to Ukraine. Diversity 2022, 14, 605. [Google Scholar] [CrossRef]
- Moshki, A.; Lamersdorf, N.P. Growth and Nutrient Status of Introduced Black Locust (Robinia pseudoacacia L.) Afforestation in arid and semi arid areas of Iran. Res. J. Environ. Sci. 2011, 5, 259–268. [Google Scholar] [CrossRef]
- Cierjacks, A.; Kowarik, I.; Joshi, J.; Hempel, S.; Ristow, M.; von der Lippe, M.; Weber, E. Biological flora of the British Isles: Robinia pseudoacacia. J. Ecol. 2013, 101, 1623–1640. [Google Scholar] [CrossRef]
- Liu, X.; Fan, Y.; Long, J.; Wei, R.; Kjelgren, R.; Gong, C.; Zhao, J. Effects of soil water and nitrogen availability on photosynthesis and water use efficiency of Robinia pseudoacacia seedlings. J. Environ. Sci. 2013, 25, 585–595. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, W.; Xu, M.; Deng, J.; Han, X.; Yang, G.; Feng, Y.Z.; Ren, G.X. Response of forest growth to C: N: P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 2019, 337, 280–289. [Google Scholar] [CrossRef]
- Rédei, K.; Csiha, I.; Keserű, Z.; Kamandiné Végh, Á.; Győri, J. The silviculture of black locust (Robinia pseudoacacia L.) in Hungary: A review. SEEFOR-South-East Eur. For. 2011, 2, 101–107. [Google Scholar] [CrossRef]
- Usta, A.; Yilmaz, M. Topographic controls in the distributions of tree species on the Karadağ Massif, NE Turkey. Balt. For. 2022, 28, 33–39. [Google Scholar] [CrossRef]
- Dyakov, N.R. Spatial distribution of some alien plants across a restricted mountainous area. Contemp. Probl. Ecol. 2016, 9, 663–673. [Google Scholar] [CrossRef]
- Vítková, M.; Tonika, J.; Müllerová, J. Black locust–successful invader of a wide range of soil conditions. Sci. Total Environ. 2015, 505, 315–328. [Google Scholar] [CrossRef]
- Sádlo, J.; Vítková, M.; Pergl, J.; Pyšek, P. Towards site-specific management of invasive alien trees based on the assessment of their impacts: The case of Robinia pseudoacacia. NeoBiota 2017, 35, 1–34. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Fodor, E.; Hâruţa, O. Niche partition of two invasive insect species, Parectopa robiniella (Lepidoptera; Gracillariidae) and Phyllonorycter robiniella (Clem.) (Lepidoptera: Gracillariidae). Res. J. Agric. Sci. 2009, 41, 261–269. [Google Scholar]
- Medzihorský, V.; Trombik, J.; Parectopa, R.; Turčáni, M.; Liebhold, A.M. Insect invasions track a tree invasion: Global distribution of black locust herbivores. J. Biogeogr. 2023, 50, 1285–1298. [Google Scholar] [CrossRef]
- Staszak, A.M.; Ratajczak, E.; Leśniewska, J.; Piotrowska-Niczyporuk, A.; Kostro-Ambroziak, A. A broad spectrum of host plant responses to the actions of the gall midge: Case study of Robinia pseudoacacia L. and Obolodiplosis robiniae (Haldeman). BMC Plant Biol. 2023, 23, 19. [Google Scholar] [CrossRef] [PubMed]
- Huemer, P.; Mayr, T. Chrysaster ostensackenella (Fitch, 1859), a potentially invasive species newly recorded from Europe (Lepidoptera, Gracillariidae). Check List. 2022, 18, 1237–1242. [Google Scholar] [CrossRef]
- Ivinskis, P.; Rimšaitė, J. Records of Phyllonorycter robiniella (Clemens, 1859) and Parectopa robiniella Clemens, 1863 (Lepidoptera, Gracillariidae) in Lithuania. Acta Zool Litu. 2008, 18, 130–133. [Google Scholar] [CrossRef]
- Tóth, P.; Váňová, M.; Lukáš, J. Impact of natural enemies on Obolodiplosis robiniae invasion. Biologia 2011, 66, 870–876. [Google Scholar] [CrossRef]
- Netoiu, C.; Tomescu, R. Moliile miniere ale salcâmului (Parectopa robiniella Clemens, 1863 si Phyllonorycter robiniella Clemens 1859, Lepidoptera, Gracillariidae). Analele ICAS 2006, 49, 119–131. [Google Scholar]
- Zhao, J.Q.; Gao, T.; Du, J.J.; Shi, J. Future trends in Obolodiplosis robiniae distribution across Eurasian continent under global climate change. Insects 2023, 14, 48. [Google Scholar] [CrossRef] [PubMed]
- Olenici, N.; BĂLĂCenoiu, F.; Tomescu, R.; NeȚOiu, C.; Buzatu, A.; Alexandru, A. Invasive alien forest insect species in south-eastern Romania. Not. Bot. Horti Agrobot. 2022, 50, 12618. [Google Scholar] [CrossRef]
- Chardon, N.I.; Cornwell, W.K.; Flint, L.E.; Flint, A.; Ackerly, D. Topographic, latitudinal and climatic distribution of Pinus coulteri: Geographic range limits are not at the edge of the climate envelope. Ecography 2015, 38, 590–601. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Sandel, B.; Blank, D.; Liu, Z.T.; Liu, X. Climate and topography explain range sizes of terrestrial vertebrates. Nat. Clim. Chang. 2016, 6, 498–502. [Google Scholar] [CrossRef]
- Michalak, J.L.; Lawler, J.J.; Roberts, D.R.; Carroll, C. Distribution and protection of climatic refugia in North America. Conserv. Biol. 2018, 32, 1414–1425. [Google Scholar] [CrossRef]
- Bartlett, J.C.; Convey, P.; Pertierra, L.R.; Hayward, S.A. An insect invasion of Antarctica: The past, present and future distribution of Eretmoptera murphyi (Diptera, Chironomidae) on Signy Island. Insect Conserv. Divers. 2020, 13, 77–90. [Google Scholar] [CrossRef]
- Oliver, T.H.; Morecroft, M.D. Interactions between climate change and land-use change on biodiversity: Attribution problems, risks, and opportunities. Wires Clim. Chang. 2014, 5, 317–335. [Google Scholar] [CrossRef]
- Bessa, A.S.; Carvalho, J.; Gomes, A.; Santarem, F. Climate and land-use drivers of invasion: Predicting the expansion of Vespa velutina nigrithorax into the Iberian Peninsula. Insect Conserv. Divers. 2016, 9, 27–37. [Google Scholar] [CrossRef]
- Hill, M.P.; Gallardo, B.; Terblanche, J.S. A global assessment of climatic niche shifts and human influence in insect invasions. Glob. Ecol. Biogeogr. 2017, 26, 679–689. [Google Scholar] [CrossRef]
- Guo, Q.; Fei, S.; Potter, K.M.; Liebhold, A.M.; Wen, J. Tree diversity regulates forest pest invasion. Proc. Natl. Acad. Sci. USA 2019, 116, 7382–7386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Nie, P.; Hu, X.; Feng, J. Future range expansions of invasive wasps suggest their increasing impacts on global apiculture. Insects 2024, 15, 546. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Xu, Z.; Han, Y.; Guo, W. Evaluation of CMIP6 models toward dynamical downscaling over 14 CORDEX domains. Clim. Dyn. 2022, 62, 4475–4489. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Nie, P.; Feng, J. Niche and Range Shifts of Aedes aegypti and Ae. albopictus Suggest That the Latecomer Shows a Greater Invasiveness. Insects 2023, 14, 810. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef]
- Ciuvăț, A.L.; Abrudan, I.V.; Ciuvăț, C.G.; Marcu, C.; Lorenț, A.; Dincă, L.; Szilard, B. Black locust (Robinia pseudoacacia L.) in Romanian forestry. Diversity 2022, 14, 780. [Google Scholar] [CrossRef]
- Canelles, Q.; Bassols, E.; Vayreda, J.; Brotons, L. Predicting the potential distribution and forest impact of the invasive species Cydalima perspectalis in Europe. Ecol. Evol. 2021, 11, 5713–5727. [Google Scholar] [CrossRef]
- Lach, L. Invasive ant establishment, spread, and management with changing climate. Curr. Opin. Insect Sci. 2021, 47, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.J.; Wang, S.J.; Yu, C.M.; Wang, X.T.; Wang, R.; Wan, J.Z. Habitat heterogeneity and topographic variation as the drivers of insect pest distributions in alpine landscapes. Acta Ecol. Sin. 2023, 43, 596–603. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, F.X.; Li, L.P.; Wang, C.J.; Wan, J.Z. Effects of habitat heterogeneity and topographic variation on insect pest risks in alpine regions. Land 2023, 12, 1314. [Google Scholar] [CrossRef]
- Baidya, P.; Bagchi, S. Influence of human land use and invasive species on beta diversity of tropical ant assemblages. Insect Conserv. Divers. 2022, 15, 73–85. [Google Scholar] [CrossRef]
- Della Rocca, F.; Milanesi, P. The new dominator of the world: Modeling the global distribution of the Japanese beetle under land use and climate change scenarios. Land 2022, 11, 567. [Google Scholar] [CrossRef]
- Montgomery, K.; Walden-Schreiner, C.; Saffer, A.; Jones, C.; Seliger, B.J.; Worm, T.; Tateosian, L.; Shukunobe, M.; Kumar, S.; Meentemeyer, R.K. Forecasting global spread of invasive pests and pathogens through international trade. Ecosphere 2023, 14, e4740. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, C.; Nie, P.; Feng, J.; Hu, X. invasive pest and invasive host: Where might spotted-wing drosophila (Drosophila suzukii) and American black cherry (Prunus serotina) cross paths in Europe? Forests 2024, 15, 206. [Google Scholar] [CrossRef]
- Acosta, A.N.C.; Mooney, K. Effects of geographic variation in host plant resources for a specialist herbivore’s contemporary and future distribution. Ecosphere 2021, 12, e03822. [Google Scholar] [CrossRef]
- Liu, T.M.; Wang, J.M.; Hu, X.K.; Feng, J.M. Land-use change drives present and future distributions of Fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Sci. Total Environ. 2020, 706, 135872. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Yamanaka, T.; Roques, A.; Augustin, S.; Chown, S.L.; Brockerhoff, E.G.; Pyšek, P. Plant diversity drives global patterns of insect invasions. Sci. Rep. 2018, 8, 12095. [Google Scholar] [CrossRef]
- Sirami, C.; Caplat, P.; Popy, S.; Clamens, A.; Arlettaz, R.; Jiguet, F.; Brotons, L.; Martin, J.L. Impacts of global change on species distributions: Obstacles and solutions to integrate climate and land use. Glob. Ecol. Biogeogr. 2017, 26, 385–394. [Google Scholar] [CrossRef]
- Begum, S.; Nakaba, S.; Yamagishi, Y.; Oribe, Y.; Funada, R. Regulation of cambial activity in relation to environmental conditions: Understanding the role of temperature in wood formation of trees. Physiol. Plant. 2013, 147, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Hacket-Pain, A.J.; Ascoli, D.; Vacchiano, G.; Biondi, F.; Cavin, L.; Conedera, M.; Drobyshev, I.; Liñán, I.D.; Friend, A.D.; Grabner, M.; et al. Climatically controlled reproduction drives interannual growth variability in a temperate tree species. Ecol. Lett. 2018, 21, 1833–1844. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.K.; Julio Camarero, J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–182. [Google Scholar] [CrossRef]
- Friend, A.D.; Eckes-Shephard, A.H.; Fonti, P.; Rademacher, T.T.; Rathgeber, C.B.K.; Richardson, A.D.; Turton, R.H. On the need to consider wood formation processes in global vegetation models and a suggested approach. Ann. For. Sci. 2019, 76, 49. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Nie, P.; Hu, X.; Feng, J. A Host Tree and Its Specialist Insects: Black Locust (Robinia pseudoacacia) Availability Largely Determines the Future Range Dynamics of Its Specialist Insects in Europe. Insects 2024, 15, 765. https://doi.org/10.3390/insects15100765
Zhang X, Nie P, Hu X, Feng J. A Host Tree and Its Specialist Insects: Black Locust (Robinia pseudoacacia) Availability Largely Determines the Future Range Dynamics of Its Specialist Insects in Europe. Insects. 2024; 15(10):765. https://doi.org/10.3390/insects15100765
Chicago/Turabian StyleZhang, Xueyou, Peixiao Nie, Xiaokang Hu, and Jianmeng Feng. 2024. "A Host Tree and Its Specialist Insects: Black Locust (Robinia pseudoacacia) Availability Largely Determines the Future Range Dynamics of Its Specialist Insects in Europe" Insects 15, no. 10: 765. https://doi.org/10.3390/insects15100765
APA StyleZhang, X., Nie, P., Hu, X., & Feng, J. (2024). A Host Tree and Its Specialist Insects: Black Locust (Robinia pseudoacacia) Availability Largely Determines the Future Range Dynamics of Its Specialist Insects in Europe. Insects, 15(10), 765. https://doi.org/10.3390/insects15100765