
Optimizing the Release Pattern of Telenomus podisi for Effective Biological Control of Euschistus heros in Soybean
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Laboratory Rearing of E. heros and T. podisi
2.2. Evaluation of Dispersal in the Field
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bueno, R.C.O.F.; Parra, J.R.P.; de Freitas Bueno, A. Trichogramma pretiosum parasitism and dispersal capacity: A basis for developing biological control programs for soybean caterpillars. Bull. Entomol. Res. 2012, 102, 1–8. [Google Scholar] [CrossRef]
- de Freitas Bueno, A.; Braz, É.C.; Favetti, B.M.; de Barros França-Neto, J.; Silva, G.V. Release of the egg parasitoid Telenomus podisi to manage the neotropical brown stink bug, Euschistus heros, in soybean production. Crop Prot. 2020, 137, 105310. [Google Scholar] [CrossRef]
- Michereff, M.F.F.; Michereff Filho, M.; Blassioli-Moraes, M.C.; Laumann, R.A.; Diniz, I.R.; Borges, M. Effect of resistant and susceptible soybean cultivars on the attraction of egg parasitoids under field conditions. J. Appl. Entomol. 2015, 139, 207–216. [Google Scholar] [CrossRef]
- Mapa Ato No 82, De 25 de Novembro de 2019, Diário Oficial da União. 2019, 229. Available online: https://www.in.gov.br/web/dou/-/ato-n-82-de-25-de-novembro-de-2019-229899956 (accessed on 1 January 2024).
- Naranjo-Guevara, N.; Santos, L.A.O.; Barbosa, N.C.C.P.; Castro, A.C.M.C.; Fernandes, O.A. Long-term mass rearing impacts performance of the egg parasitoid Telenomus remus (Hymenoptera: Platygastridae). J. Entomol. Sci. 2020, 55, 69–86. [Google Scholar] [CrossRef]
- Grande, M.L.M.; de Queiroz, A.P.; Gonçalves, J.; Hayashida, R.; Ventura, M.U.; de Freitas Bueno, A. Impact of environmental variables on parasitism and emergence of Trichogramma pretiosum, Telenomus remus and Telenomus podisi. Neotrop. Entomol. 2021, 50, 605–614. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.C.; Ikuno, P.H.P.; Pratissoli, D.; de Carvalho, J.R.; Hoback, W.W.; Salamina, B.A.Z. Biological characteristics and thermal requirements of Telenomus podisi and Trissolcus basalis (Hymenoptera: Scelionidae) in fresh and cryopreserved eggs of Euschistus heros and Nezara viridula (Hemiptera: Pentatomidae). Agronomy 2024, 14, 170. [Google Scholar] [CrossRef]
- Parra, L.M.; de Carvalho, J.R.; Hoback, W.W.; de Oliveira, R.C. Optimizing mass rearing of the egg parasitoid, Telenomus podisi, for control of the brown stink bug, Euschistus heros. Insects 2023, 14, 435. [Google Scholar] [CrossRef]
- Doetzer, A.K.; Foerster, L.A. Storage of pentatomid eggs in liquid nitrogen and dormancy of Trissolcus basalis (Wollaston) and Telenomus podisi Ashmead (Hymenoptera: Platygastridae) adults as a method of mass production. Neotrop. Entomol. 2013, 42, 534–538. [Google Scholar] [CrossRef]
- Hayashida, R.; de Freitas Bueno, A.; Hermel, A.O.; Hirakuri, M.H.; Silva, F.A.C.; Roggia, S. Euschistus heros (Hemiptera: Pentatomidae) fitness on artificial diets: An approach to optimize mass rearing of Telenomus podisi (Hymenoptera: Platygastridae) for augmentative biological control. J. Econ. Entomol. 2018, 111, 1605–1613. [Google Scholar] [CrossRef]
- Silva, C.C.; Laumann, R.A.; Blassioli, M.C.; Pareja, M.; Borges, M. Euschistus heros mass rearing technique for the multiplication of Telenomus podisi. Pesqui. Agropecuária Bras. 2008, 43, 575–580. [Google Scholar] [CrossRef]
- Queiroz, A.P.; Favetti, B.M.; Hayashida, R.; Grande, M.L.M.; Neiva, M.M.; Panizzi, A.R.; de Freitas Bueno, A. Effect of the ages of parasitoid and hoste eggs on Telenomus podisi (Hymenoptera: Platygastridae) parasitism. Neotrop. Entomol. 2019, 48, 974–982. [Google Scholar] [CrossRef]
- Golin, V. Incidência Natural, Biologia, Seletividade e Efeito de Liberações Inoculativas de Parasitoides de Ovos (Hymenoptera: Platygastridae) no Controle de Euschistus heros (Fabricius) (Hemiptera: Pentatomidae) no Mato Grosso. 2014. Available online: https://acervodigital.ufpr.br/handle/1884/36836 (accessed on 1 January 2024).
- Li, D.; Yuan, X.; Zhang, B.; Zhao, Y.; Song, Z.; Zuo, C. Report of using unmanned aerial vehicle to release Trichogramma. Chin. J. Biol. Control 2013, 29, 455. [Google Scholar]
- Borges, M.; Laumann, R.A.; Da Silva, C.C.A.; Moraes, M.C.B.; Dos Santos, H.M.; Ribeiro, D.T. Metodologias de criação e manejo de colônias de percevejos da soja (Hemiptera-Pentatomidae) para estudos de comportamento e ecologia química. In Embrapa Recursos Genéticos e Biotecnologia. Documentos; Embrapa Recursos Genéticos e Biotecnologia: Brasília, Brazil, 2006; Volume 182. [Google Scholar]
- Peres, W.A.A.; Corrêa-Ferreira, B.S. Methodology of mass multiplication of Telenomus podisi Ash. and Trissolcus basalis (Woll.) (Hymenoptera: Scelionidae) on eggs of Euschistus heros (Fab.) (Hemiptera: Pentatomidae). Neotrop. Entomol. 2004, 33, 457–462. [Google Scholar] [CrossRef]
- Matei, G.; Benin, G.; Woyann, L.G.; Dalló, S.C.; Milioli, A.S.; Zdziarski, A.D. Agronomic performance of modern soybean cultivars in multi-environment trials. Pesqui. Agropecu. Bras. 2017, 52, 500–511. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Shimizu, G.D.; Marubayashi, R.Y.P.; Goncalves, L.S.A. Experimental Statistics and Graphics for Agricultural Sciences; Londrina. 2022. Available online: https://rdrr.io/cran/AgroR/ (accessed on 1 January 2024).
- Wickham, H. Programming with Ggplot2. In ggplot2; Springer: Berlin/Heidelberg, Germany, 2016; pp. 241–253. [Google Scholar]
- Burr, I.W.; Foster, L.A. A Test for Equality of Variances; University of Purdue: West Lafayette, IN, USA, 1972. [Google Scholar]
- Beer, T. Beaufort Wind Scale. In Encyclopedia of Natural Hazards; Bobrowsky, P.T., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 42–45. ISBN 978-1-4020-4399-4. [Google Scholar]
- Da Fonseca, P.R.B.; Fernandes, M.G.; Justiniano, W.; Cavada, L.H.; da Silva, J.A.N. Spatial distribution of adults and nymphs of Euschistus heros (F.) (Hemiptera: Pentatomidae) on bt and non-bt soybean. J. Agric. Sci. 2014, 6, 131–142. [Google Scholar] [CrossRef]
- Orr, D.B.; Entomologist, F.; Orr, D.B. Scelionid wasps as biological control agents: A review. Fla. Entomol. 1988, 71, 506–528. [Google Scholar] [CrossRef]
- Weber, I.D.; Garcia, A.G.; de Freitas Bueno, A.; Oliveira, R.C.; Godoy, W.A.C. Release strategies of Telenomus podisi for control of Euschistus heros: A computational modeling approach. Pest Manag. Sci. 2022, 78, 4544–4556. [Google Scholar] [CrossRef] [PubMed]
- Hayashida, R.; Godoy, C.V.; Hoback, W.W.; de Freitas Bueno, A. Are economic thresholds for IPM decisions the same for low LAI soybean cultivars in Brazil? Pest Manag. Sci. 2021, 77, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Moraes, M.C.B.; Laumann, R.A.; Pareja, M.; Sereno, F.T.P.S.; Michereff, M.F.F.; Birkett, M.A.; Pickett, J.A.; Borges, M. Attraction of the stink bug egg parasitoid Telenomus podisi to defence signals from soybean activated by treatment with cis-jasmone. Entomol. Exp. Appl. 2009, 131, 178–188. [Google Scholar] [CrossRef]
- Dias, A.M.; Pareja, M.; Laia, M.; Blassioli-Moraes, M.C.; Borges, M.; Laumann, R.A. Attraction of Telenomus podisi to volatiles induced by Euschistus heros in three different plant species. Arthropod. Plant. Interact. 2016, 10, 419–428. [Google Scholar] [CrossRef]
- Havill, N.P.; Raffa, K.F. Compound effects of induced plant responses on insect herbivores and parasitoids: Implications for tritrophic interactions. Ecol. Entomol. 2000, 25, 171–179. [Google Scholar] [CrossRef]
- Heimpel, G.E. Linking parasitoid nectar feeding and dispersal in conservation biological control. Biol. Control 2019, 132, 36–41. [Google Scholar] [CrossRef]
- Tognon, R.; Sant’Ana, J.; Michereff, M.F.F.; Laumann, R.A.; Borges, M.; Blassioli-Moraes, M.C.; Redaelli, L.R. Kairomones from Euschistus heros egg masses and their potential use for Telenomus podisi parasitism improvement. Bull. Entomol. Res. 2020, 110, 638–644. [Google Scholar] [CrossRef]
- Favetti, B.M.; Butnariu, A.R.; Doetzer, A.K. Storage of Euschistus heros eggs (Fabricius) (Hemiptera: Pentatomidae) in liquid nitrogen for parasitization by Telenomus podisi Ashmead (Hymenoptera: Platygastridae). Neotrop. Entomol. 2014, 43, 291–293. [Google Scholar] [CrossRef]
- Manzano, C.; Fernandez, P.; Hill, J.; Luft Albarracin, E.; Virla, E.; Coll Aráoz, M. Chemical ecology of the host searching behavior in an egg parasitoid: Are common chemical cues exploited to locate hosts in taxonomically distant plant species? J. Chem. Ecol. 2022, 48, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Barrufaldi, A.P.F.; Hayashida, R.; Hoback, W.W.; Higley, L.G.; De Carvalho, J.R.; De Oliveira, R.C. Trade-offs between temperature and fitness in Euschistus heros (Fabricius) (Hemiptera: Pentatomidae): Implications for mass rearing and field management. Insects 2023, 14, 448. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, D.J.P.; Corrêa-Ferreira, B.S. Parasitismo de Telenomus podisi Ashmead (Hymenoptera: Scelionidae) em populações de percevejos pragas da soja. An. Soc. Entomol. Bras. 2000, 29, 295–302. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Dieckhoff, C.; Vinyard, B.T.; Hoelmer, K.A. Parasitism and predation on sentinel egg masses of the brown marmorated stink bug (Hemiptera: Pentatomidae) in three vegetable crops: Importance of dissections for evaluating the impact of native parasitoids on an exotic pest. Environ. Entomol. 2016, 45, 1536–1542. [Google Scholar] [CrossRef]
- Pomari, A.F.; de Freitas Bueno, A.; de Freitas Bueno, R.C.O.; de Oliveiras Menezes Junior, A.; Fonseca, A.C.P.F. Releasing number of Telenomus remus (Nixon)(Hymenoptera: Platygastridae) against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in corn, cotton and soybean. Ciência Rural 2013, 43, 377–382. [Google Scholar] [CrossRef]
- Asplen, M.K.; Chacón, J.M.; Heimpel, G.E. Sex-specific dispersal by a parasitoid wasp in the field. Entomol. Exp. Appl. 2016, 159, 252–259. [Google Scholar] [CrossRef]
- Zhou, J.; Dong, Q.; Zhang, T.; Duan, L.; Ning, S.; Liu, Q.; Li, Y.; Li, C.; Dong, H. Effect of wind time on the dispersal capacity of Trichogramma dendrolimi Matsumura (Hymenoptera Trichogrammatidae). J. Asia Pac. Entomol. 2019, 22, 742–749. [Google Scholar] [CrossRef]
- Salazar-Mendoza, P.; Rodriguez-Saona, C.; Aparecido Fernandes, O. Release density, dispersal capacity, and optimal rearing conditions for Telenomus remus, an egg parasitoid of Spodoptera frugiperda, in maize. Biocontrol Sci. Technol. 2020, 30, 1040–1059. [Google Scholar] [CrossRef]
- Fernandes Cordeiro, J.P.; de Freitas Bueno, A. Trissolcus teretis (Johnson, 1987) (Hymenoptera: Scelionidae) parasitism on Euschistus heros (Fabricius, 1798) and Diceraeus melacanthus Dallas, 1851 (Hemiptera: Pentatomidae) eggs at different temperatures. Rev. Bras. Entomol. 2021, 65, e20200073. [Google Scholar] [CrossRef]
- Nunes, M.C.; Corrêa-Ferreira, B.S. Danos causados à soja por adultos de Euschistus heros (Fabricius) (Hemiptera: Pentatomidae), sadios e parasitados por Hexacladia smithii Ashmead (Hymenoptera: Encyrtidae). Neotrop. Entomol. 2002, 31, 109–113. [Google Scholar] [CrossRef]
- Corrêa-Ferreira, B.S.; Nunes, M.C.; Uguccioni, L.D. Ocorrência do parasitóide Hexacladia smithii Ashmead em adultos de Euschistus heros (F.) no Brasil. An. Soc. Entomol. Bras. 1998, 27, 495–498. [Google Scholar] [CrossRef]
- Zerbino, M.S.; Panizzi, A.R. The underestimated role of pest pentatomid parasitoids in Southern South America. Arthropod. Plant. Interact. 2019, 13, 703–718. [Google Scholar] [CrossRef]
- Tuelher, E.S.; Silva, É.H.; Hirose, E.; Guedes, R.N.C.; Oliveira, E.E. Competition between the phytophagous stink bugs Euschistus heros and Piezodorus guildinii in soybeans. Pest Manag. Sci. 2016, 72, 1837–1843. [Google Scholar] [CrossRef]
- de Freitas Bueno, A.; Paula-Moraes, S.V.; Gazzoni, D.L.; Pomari, A.F. Economic thresholds in soybean-integrated pest management: Old concepts, current adoption, and adequacy. Neotrop. Entomol. 2013, 42, 439–447. [Google Scholar] [CrossRef]
- Magano, D.A.; Machado, M.R.R.; Jerónimo, J.A.; Guedes, J.V.C.; Doberstein, A.P.S. Modelagem geostatistica aplicada a distribuição espacial de lagartas presentes na cultura da soja: Geostatistic modeling applied to the spatial distribution of caterpillars present in soybean crop. Braz. J. Anim. Environ. Res. 2023, 6, 20–29. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
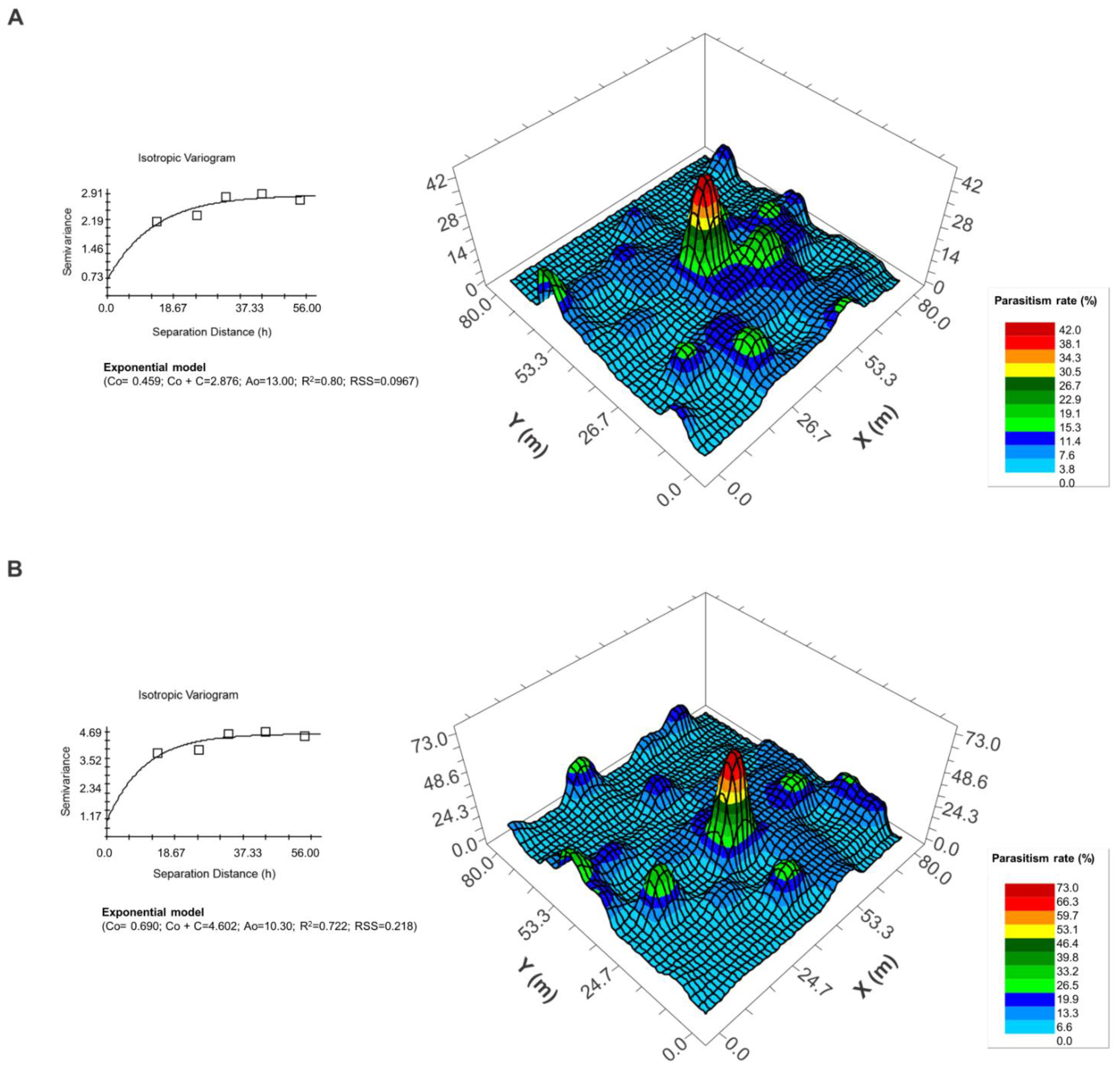{kind=link}
| Matrix | T °C | Wind (m/s) | Direction | RH % |
|---|---|---|---|---|
| Vn–R1 | ||||
| M1 | 23.3 | 0.9 | east | 71 |
| M2 | 22.3 | 1.0 | east | 68 |
| M3 | 22.7 | 1.5 | west | 70 |
| M4 | 22.1 | 1.5 | east | 71 |
| M5 | 22.0 | 1.8 | east | 79 |
| M6 | 22.5 | 2.1 | east | 83 |
| R5–R6 | ||||
| M1 | 23.3 | 1.5 | west | 77 |
| M2 | 22.3 | 1.0 | east | 86 |
| M3 | 22.7 | 2.0 | west | 72 |
| M4 | 22.1 | 1.0 | east | 82 |
| M5 | 22.0 | 1.0 | east | 59 |
| M6 | 22.5 | 1.2 | west | 64 |
| Stage | T °C | Wind (m/s) | RH (%) | PAtm (hPa) | Ppt mm | Date |
|---|---|---|---|---|---|---|
| Vn–R1 | 23.86 | 1.08 | 87.97 | 941.77 | - | 6 January 2020 |
| R5–R6 | 27.35 | 0.97 | 84.84 | 947.30 | 0.64 | 18 February 2020 |
| Stage | Parasitism Rate (%) | ||
|---|---|---|---|
| Inner Box | Middle Box | Outer Box | |
| Vn–R1 | 6.10 ± 1.39 aA (n = 54) | 2.67 ± 0.39 bA (n = 96) | 1.84 ± 0.24 bA (n = 228) |
| R5–R6 | 6.81 ± 1.66 aA (n = 54) | 5.16 ± 0.69 aB (n = 96) | 4.56 ± 0.46 aB (n = 228) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoback, W.W.; Ramos, G.; Hayashida, R.; Santos, D.M.; Alvarez, D.d.L.; Oliveira, R.C.d. Optimizing the Release Pattern of Telenomus podisi for Effective Biological Control of Euschistus heros in Soybean. Insects 2024, 15, 192. https://doi.org/10.3390/insects15030192
Hoback WW, Ramos G, Hayashida R, Santos DM, Alvarez DdL, Oliveira RCd. Optimizing the Release Pattern of Telenomus podisi for Effective Biological Control of Euschistus heros in Soybean. Insects. 2024; 15(3):192. https://doi.org/10.3390/insects15030192
Chicago/Turabian StyleHoback, William Wyatt, Gabryele Ramos, Rafael Hayashida, Daniel Mariano Santos, Daniel de Lima Alvarez, and Regiane Cristina de Oliveira. 2024. "Optimizing the Release Pattern of Telenomus podisi for Effective Biological Control of Euschistus heros in Soybean" Insects 15, no. 3: 192. https://doi.org/10.3390/insects15030192