Presence of Multiple Genetic Mutations Related to Insecticide Resistance in Chinese Field Samples of Two Phthorimaea Pest Species


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Insects
2.2. Genomic DNA Extraction
2.3. PCR Amplification of Target Gene Fragments and Sanger Sequencing
2.4. DNA Sequence Analysis and Phylogenetic Analysis
2.5. Molecular Clock Estimates
2.6. Syntenic Relationship of the Insecticide Target Gene Loci
3. Results
3.1. Distribution and Frequency of Insecticide Resistance-Conferring Mutations
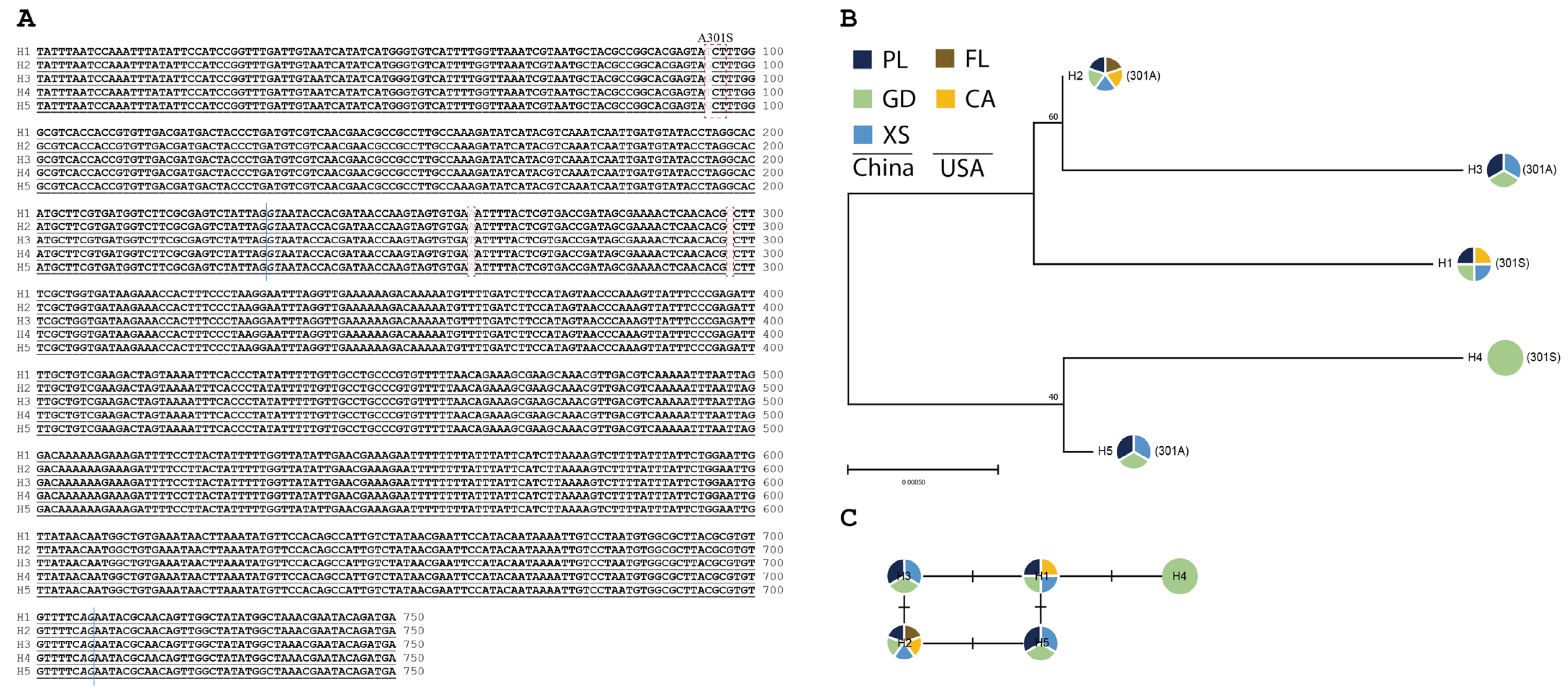
3.2. Haplotype Identification and Phylogeny
3.3. The Divergence Time among Gelechiidae
3.4. The Collinearity of Specific Genomic Regions in P. operculella and P. absoluta
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Si, H.J.; Zhang, N.; Tang, X.; Yang, J.W.; Wen, Y.K.; Wang, L.; Zhou, X.Y. Transgenic Research in Tuber and Root Crops: A Review. In Genetic Engineering of Horticultural Crops; Rout, G.R., Peter, K.V., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 225–248. [Google Scholar] [CrossRef]
- Wang, Q.B.; Zhang, W. China’s potato industry and potential impacts on the global market. Am. J. Potato Res. 2004, 81, 101–109. [Google Scholar] [CrossRef]
- Amiri, A.N.; Bakhsh, A. An effective pest management approach in potato to combat insect pests and herbicide. 3 Biotech. 2019, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Rondon, S.I. The potato tuberworm: A literature review of its biology, ecology, and control. Am. J. Potato Res. 2010, 87, 149–166. [Google Scholar] [CrossRef]
- Rondon, S.I.; Gao, Y.L. Potato tuberworm. In Insect Pests of Potato: Global Perspectives on Biology and Management; Alyokhin, A., Rondon, S.I., Gao, Y.L., Eds.; Academic Press: Oxford, UK, 2022; pp. 149–160. Available online: https://www.sciencedirect.com/book/9780123868954/insect-pests-of-potato (accessed on 2 February 2024).
- Du, L.T.; Li, Z.Y.; Zhou, L.M.; Li, C.M.; Tang, X. Comparsion of five chemicals in three groups for their effects controlling larva of potato moth. China Potato J. 2006, 20, 92–93. [Google Scholar] [CrossRef]
- Gao, Y.L. Green control techniques for potato tuberworm (Phthorimaea operculella). Sci. Agric. Sin. 2021, 54, 533–535. [Google Scholar] [CrossRef]
- Yan, J.J.; Guo, W.C.; Li, G.Q.; Pan, H.P.; Chen, B.; Zhou, W.W.; Li, S.H.; Gao, Y.L. Current status and prospects of the management of important insect pests on potato in China. Plant Protect. 2023, 49, 190–195. [Google Scholar] [CrossRef]
- Yang, J.; Wang, F.G.; Gao, S.H.; Ma, H.M. Identification and integrated prevention and control of tomato leaf miner moth, Tuta absoluta. Appl. Tech. Rural Area 2021, 3, 88–89. [Google Scholar]
- Desneux, N.; Wajnberg, E.; Wyckhuys, K.A.G.; Burgio, G.; Arpaia, S.; Narváez-Vasquez, C.A.; Gonzalez-Cabrera, J.; Ruescas, D.C.; Tabone, E.; Frandon, J.; et al. Biological invasion of European tomato crops by Tuta absoluta: Ecology, geographic expansion and prospects for biological control. J. Pest. Sci. 2010, 83, 197–215. [Google Scholar] [CrossRef]
- Desneux, N.; Luna, M.G.; Guillemaud, T.; Urbaneja, A. The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: The new threat to tomato world production. J. Pest Sci. 2011, 84, 403–408. [Google Scholar] [CrossRef]
- Brévault, T.; Bernadas, G.; Sylla, S.; Diatte, M.; Diarra, K. Tuta absoluta Meyrick (Lepidoptera: Gelechiidae): A new threat to tomato production in sub-Saharan Africa. Afr. Entomol. 2014, 22, 441–444. [Google Scholar] [CrossRef]
- Zhang, G.F.; Ma, D.Y.; Wang, Y.S.; Gao, Y.H.; Liu, W.X.; Zhang, R.; Fu, W.J.; Xian, X.Q.; Wang, J.; Kuang, M.; et al. First report of the South American tomato leafminer, Tuta absoluta (Meyrick), in China. J. Integr. Agr. 2020, 19, 1912–1917. [Google Scholar] [CrossRef]
- Desneux, N.; Han, P.; Mansour, R.; Arnó, J.; Brévault, T.; Campos, M.R.; Chailleux, A.; Guedes, R.N.C.; Karimi, J.; Konan, K.A.J.; et al. Integrated pest management of Tuta absoluta: Practical implementations across different world regions. J. Pest Sci. 2022, 95, 17–39. [Google Scholar] [CrossRef]
- Zhang, G.F.; Ma, D.Y.; Liu, W.X.; Wang, Y.S.; Fu, W.J.; Wang, J.; Gao, Y.H.; Wan, F.H. The arrival of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae), in China. J. Biosaf. 2019, 28, 200–203. [Google Scholar] [CrossRef]
- Lu, Y.Y. Invasion alert: Rapid spread of the tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae), an invasive pest in China. J. Environ. Entomol. 2021, 43, 526–528. [Google Scholar] [CrossRef]
- Zhang, G.F.; Zhang, Y.B.; Xian, X.Q.; Liu, W.X.; Li, P.; Liu, W.C.; Liu, H.; Fen, X.D.; Lv, Z.C.; Wang, Y.S.; et al. Damage of an important and newly invaded agricultural pest, Phthorimaea absoluta, and its prevention and management measures. Plant Protect. 2022, 48, 51–58. [Google Scholar]
- Zhang, M.D.; Yan, J.J.; Ali, A.; Gao, Y.L. Potato plant variety affects the performance and oviposition preference of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae). Pest Manag. Sci. 2022, 78, 3912–3919. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.J.; Zhang, M.D.; Ali, A.; Du, X.; Mei, X.D.; Gao, Y.L. Optimization and field evaluation of sex-pheromone of potato tuber moth, Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae). Pest Manag. Sci. 2022, 78, 3903–3911. [Google Scholar] [CrossRef]
- Yan, J.J.; Zhang, M.D.; Gao, Y.L. Biology, ecology and integrated management of potato tuber moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). Acta Entomol. Sin. 2019, 62, 1469–1482. [Google Scholar] [CrossRef]
- Raymond, M.; Berticat, C.; Weill, M.; Pasteur, N.; Chevillon, C. Insecticide resistance in the mosquito culex pipiens: What have we learned about adaptation? Genetica 2001, 8, 287–296. [Google Scholar] [CrossRef]
- Feyereisen, R.; Dermauw, W.; Van-Leeuwen, T. Genotype to phenotype, the molecular and physiological dimensions of resistance in arthropods. Pestic Biochem. Physiol. 2015, 121, 61–77. [Google Scholar] [CrossRef]
- Mugenzi, L.M.J.; Menze, B.D.; Tchouakui, M.; Wondji, M.J.; Irving, H.; Tchoupo, M.; Hearn, J.; Weedall, G.D.; Riveron, J.M.; Wondji, C.S. Cis-regulatory CYP6P9b P450 variants associated with loss of insecticide-treated bed net efficacy against Anopheles funestus. Nat. Commun. 2019, 10, 4652. [Google Scholar] [CrossRef] [PubMed]
- Nauen, R.; Bass, C.; Feyereisen, R.; Vontas, J. The role of cytochrome P450s in insect toxicology and resistance. Annu. Rev. Entomol. 2022, 67, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Dermauw, W.; Van-Leeuwen, T.; Feyereisen, R. Diversity and evolution of the P450 family in arthropods. Insect Biochem. Mol. Biol. 2020, 127, 103490. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect CYP genes and P450 enzymes. In Insect Molecular Biology and Biochemistry; Gilbert, L., Ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 236–316. [Google Scholar] [CrossRef]
- Boaventura, D.; Martin, M.; Pozzebon, A.; Mota-Sanchez, D.; Nauen, R. Monitoring of target-site mutations conferring insecticide resistance in Spodoptera frugiperda. Insects 2020, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Engdahl, C.; Knutsson, S.; Fredriksson, S.A.; Linusson, A.; Bucht, G.; Ekström, F. Acetylcholinesterases from the disease vectors Aedes aegypti and Anopheles gambiae: Functional characterization and comparisons with vertebrate orthologues. PLoS ONE 2015, 10, e0138598. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.C.; Callaghan, A.; Field, L.M.; Williamson, M.S.; Moores, G.D. Identification of mutations conferring insecticide-insensitive AChE in the cotton-melon aphid, Aphis gossypii Glover. Insect Mol. Biol. 2004, 13, 555–561. [Google Scholar] [CrossRef]
- Chang, C.; Cheng, X.; Huang, X.Y.; Dai, S.M. Amino acid substitutions of acetylcholinesterase associated with carbofuran resistance in Chilo suppressalis. Pest Manag. Sci. 2014, 70, 1930–1935. [Google Scholar] [CrossRef]
- Ilias, A.; Vontas, J.; Tsagkarakou, A. Global distribution and origin of target site insecticide resistance mutations in Tetranychus urticae. Insect Biochem. Mol. Biol. 2014, 48, 17–28. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Campbell, C.L.; Lozano, S.; Penilla-Navarro, P.; Lopez-Solis, A.; Solis-Santoyo, F. Rodriguez, A.D.; Perera, R.; Black Lv, W.C. Permethrin resistance in Aedes aegypti: Genomic variants that confer knockdown resistance, recovery, and death. PLoS Genet. 2021, 17, e1009606. [Google Scholar] [CrossRef]
- Zhao, Y.H.; Feng, X.Y.; Li, M.; Qiu, X.H. The double-mutation (M918I + L1014F) kdr allele is fixed in Cimex hemipterus populations in Guangxi, China. Bull Entomol. Res. 2020, 110, 506–511. [Google Scholar] [CrossRef]
- Garrood, W.T.; Zimmer, C.T.; Gutbrod, O.; Lüke, B.; Williamson, M.S.; Bass, C.; Nauen, R.; Davies, T.G.E. Influence of the RDL A301S mutation in the brown planthopper Nilaparvata lugens on the activity of phenylpyrazole insecticides. Pestic Biochem. Physiol. 2017, 142, 1–8. [Google Scholar] [CrossRef]
- Doğramacı, M.; Tingey, W.M. Comparison of insecticide resistance in a North American field population and a laboratory colony of potato tuberworm (Lepidoptera: Gelechiidae). J. Pest Sci. 2008, 81, 17–22. [Google Scholar] [CrossRef]
- El-Kady, H. Insecticide resistance in potato tuber moth Phthorimaea operculella Zeller in Egypt. J. Am. Sci. 2011, 7, 263–266. Available online: http://www.americanscience.org (accessed on 2 February 2024).
- Roditakis, E.; Steinbach, D.; Moritz, G.; Vasakis, E.; Stavrakaki, M.; Ilias, A.; García-Vidal, L.; Aguirre, M.R.M.; Bielza, P.; Morou, E.; et al. Ryanodine receptor point mutations confer diamide insecticide resistance in tomato leafminer, Tuta absoluta (Lepidoptera: Gelechiidae). Insect Biochem. Mol. Biol. 2017, 80, 11–20. [Google Scholar] [CrossRef]
- Campos, M.R.; Silva, T.B.; Silva, W.M.; Silva, J.E.; Siqueira, H.A. Spinosyn resistance in the tomato borer Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). J. Pest Sci. 2015, 88, 405–412. [Google Scholar] [CrossRef]
- Silva, J.E.; Ribeiro, L.M.D.S.; Vinasco, N.; Guedes, R.N.C.; Siqueira, H.Á.A. Field-evolved resistance to chlorantraniliprole in the tomato pinworm Tuta absoluta: Inheritance, cross-resistance profile, and metabolism. J. Pest Sci. 2019, 92, 1421–1431. [Google Scholar] [CrossRef]
- Haddi, K.; Berger, M.; Bielza, P.; Cifuentes, D.; Field, L.M.; Gorman, K.; Rapisarda, C.; Williamson, M.S.; Bass, C. Identification of mutations associated with pyrethroid resistance in the voltage-gated sodium channel of the tomato leaf miner (Tuta absoluta). Insect Biochem. Mol. Biol. 2012, 42, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Haddi, K.; Berger, M.; Bielza, P.; Rapisarda, C.; Williamson, M.S.; Moores, G.; Bass, C. Mutation in the ace-1 gene of the tomato leaf miner (Tuta absoluta) associated with organophosphates resistance. J. Appl. Entomol. 2017, 141, 612–619. [Google Scholar] [CrossRef]
- Kwon, M.; Kim, J.; Maharjan, R.; Choi, J.Y.; Kim, G.H. Change in the distribution of the potato tuber moth, Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae), in Korea. J. Asia-Pac. Entomol. 2017, 20, 1249–1253. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Zhang, L.; Hamm, R.L.; Brady, S.G.; Lazzaro, B.P.; Scott, J.G. Frequencies of the pyrethroid resistance alleles of Vssc1 and CYP6D1 in house flies from the eastern United States. Insect Mol. Biol. 2006, 15, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Paradis, E. Analysis of haplotype networks: The randomized minimum spanning tree method. Methods Ecol. Evol. 2018, 9, 1308–1317. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bielza, P. Insecticide resistance in natural enemies. In Advances in Insect Control and Resistance Management; Horowitz, A.R., Ishaaya, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 313–329. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, Y.H.; Kwon, D.H.; Cha, D.J.; Kim, J.H. Mutation and duplication of arthropod acetylcholinesterase: Implications for pesticide resistance and tolerance. Pestic Biochem. Physiol. 2015, 120, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Zhen, C.; Miao, L.; Liang, P.; Gao, X.W. Survey of organophosphate resistance and an Ala216Ser substitution of acetylcholinesterase-1 gene associated with chlorpyrifos resistance in Apolygus lucorum (Meyer-Dür) collected from the transgenic Bt cotton fields in China. Pestic Biochem. Physiol. 2016, 132, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Khajehali, J.; Van-Leeuwen, T.; Grispou, M.; Morou, E.; Alout, H.; Weill, M.; Tirry, L.; Vontas, J.; Tsagkarakou, A. Acetylcholinesterase point mutations in European strains of Tetranychus urticae (Acari: Tetranychidae) resistant to organophosphates. Pest Manag. Sci. 2010, 66, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yoon, K.A.; Lee, S.H. Short communication: Screening of proof-of-concept mutations of honey bee acetylcholinesterase 2 conferring resistance to organophosphorus and carbamate insecticides. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2023, 264, 109524. [Google Scholar] [CrossRef]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.X.; Sliver, K.; Zhorov, B. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lynd, A.; Oruni, A.; Van’t, H.A.E.; Morgan, J.C.; Naego, L.B.; Pipini, D.; O’Kines, K.A.; Bobanga, T.L.; Donnelly, M.J.; Weetman, D. Insecticide resistance in Anopheles gambiae from the northern Democratic Republic of Congo, with extreme knockdown resistance (kdr) mutation frequencies revealed by a new diagnostic assay. Malar. J. 2018, 17, 412. [Google Scholar] [CrossRef]
- Tian, X.R.; Liu, J.D.; Guo, Z.M.; Hu, B.; Kibe, M.D.; Wang, S.Y.; Wei, Q.; Su, J.Y. The characteristics of voltage-gated sodium channel and the association with lambda cyhalothrin resistance in Spodoptera exigua. J. Asia-Pac. Entomol. 2018, 21, 1020–1027. [Google Scholar] [CrossRef]
- Ghavami, M.B.; Haghi, F.P.; Alibabaei, Z.; Enayati, A.A.; Vatandoost, H. First report of target site insensitivity to pyrethroids in human flea, Pulex irritans (Siphonaptera: Pulicidae). Pestic Biochem. Physiol. 2018, 146, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Schuler, T.H.; Martinez-Torres, D.; Thompson, A.J.; Denholm, I.; Devonshire, A.L.; Duce, I.R.; Williamson, M.S. Toxicological, electrophysiological, and molecular characterisation of knockdown resistance to pyrethroid insecticides in the diamondback moth, Plutella xylostella (L.). Pestic Biochem Phys. 1998, 59, 169–182. [Google Scholar] [CrossRef]
- Usherwood, P.N.; Davies, T.G.; Mellor, I.R.; O’Reilly, A.O.; Peng, F.; Vais, H.; Khambay, B.P.S.; Field, L.M.; Williamson, M.S. Mutations in DIIS5 and the DIIS4-S5 linker of Drosophila melanogaster sodium channel define binding domains for pyrethroids and DDT. FEBS Lett. 2007, 581, 5485–5492. [Google Scholar] [CrossRef]
- Samantsidis, G.R.; Panteleri, R.; Denecke, S.; Kounadi, S.; Christou, I.; Nauen, R.; Douris, V.; Vontas, J. ‘What I cannot create, I do not understand’: Functionally validated synergism of metabolic and target site insecticide resistance. Proc. Biol. Sci. 2020, 287, 20200838. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Populations | n | F158Y | A201S | L231V | F290V | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | F/Y | Y | A | A/S | S | L | L/V | V | F | F/V | V | |||
| P. operculella | LL | 30 | 83 | 10 | 7 | 87 | 13 | 0 | 0 | 0 | 100 | 100 | 0 | 0 |
| ML | 30 | 86 | 7 | 7 | 90 | 7 | 3 | 0 | 0 | 100 | 100 | 0 | 0 | |
| XW | 30 | 90 | 10 | 0 | 100 | 0 | 0 | 0 | 0 | 100 | 100 | 0 | 0 | |
| P. absoluta | PL | 30 | 100 | 0 | 0 | 0 | 77 | 23 | 77 | 23 | 0 | 40 | 60 | 0 |
| GD | 30 | 100 | 0 | 0 | 0 | 67 | 33 | 63 | 37 | 0 | 73 | 27 | 0 | |
| XS | 30 | 100 | 0 | 0 | 0 | 63 | 37 | 97 | 3 | 0 | 70 | 30 | 0 | |
| Species | Populations | n | M918T | L925M | T929I | L1014F | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | M/T | T | L | L/M | M | T | T/I | I | L | L/F | F | |||
| P. operculella | LL | 30 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 84 | 13 | 3 |
| ML | 30 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 87 | 10 | 3 | |
| XW | 30 | 100 | 0 | 0 | 100 | 0 | 0 | 100 | 0 | 0 | 77 | 20 | 3 | |
| P. absoluta | PL | 30 | 30 | 47 | 23 | 93 | 7 | 0 | 23 | 47 | 30 | 0 | 0 | 100 |
| GD | 30 | 53 | 40 | 7 | 90 | 10 | 0 | 10 | 47 | 43 | 0 | 0 | 100 | |
| XS | 30 | 50 | 37 | 13 | 93 | 7 | 0 | 13 | 40 | 47 | 0 | 0 | 100 | |
| Species | Populations | n | A301S | ||
|---|---|---|---|---|---|
| A/A | A/S | S/S | |||
| P. operculella | LL | 30 | 100 | 0 | 0 |
| ML | 30 | 100 | 0 | 0 | |
| XW | 30 | 100 | 0 | 0 | |
| P. absoluta | PL | 30 | 63 | 20 | 17 |
| GD | 30 | 66 | 17 | 17 | |
| XS | 30 | 54 | 23 | 23 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Chen, R.; Liu, J.; Lin, W.; Liang, J.; Nauen, R.; Li, S.; Gao, Y. Presence of Multiple Genetic Mutations Related to Insecticide Resistance in Chinese Field Samples of Two Phthorimaea Pest Species. Insects 2024, 15, 194. https://doi.org/10.3390/insects15030194
Zhu J, Chen R, Liu J, Lin W, Liang J, Nauen R, Li S, Gao Y. Presence of Multiple Genetic Mutations Related to Insecticide Resistance in Chinese Field Samples of Two Phthorimaea Pest Species. Insects. 2024; 15(3):194. https://doi.org/10.3390/insects15030194
Chicago/Turabian StyleZhu, Jiang, Ruipeng Chen, Juan Liu, Weichao Lin, Jiaxin Liang, Ralf Nauen, Suhua Li, and Yulin Gao. 2024. "Presence of Multiple Genetic Mutations Related to Insecticide Resistance in Chinese Field Samples of Two Phthorimaea Pest Species" Insects 15, no. 3: 194. https://doi.org/10.3390/insects15030194
APA StyleZhu, J., Chen, R., Liu, J., Lin, W., Liang, J., Nauen, R., Li, S., & Gao, Y. (2024). Presence of Multiple Genetic Mutations Related to Insecticide Resistance in Chinese Field Samples of Two Phthorimaea Pest Species. Insects, 15(3), 194. https://doi.org/10.3390/insects15030194