
Revealing a Novel Potential Pest of Plum Trees in the Caucasus: A Species Resembling the European Leaf-Mining Stigmella plagicolella, Nepticulidae #

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
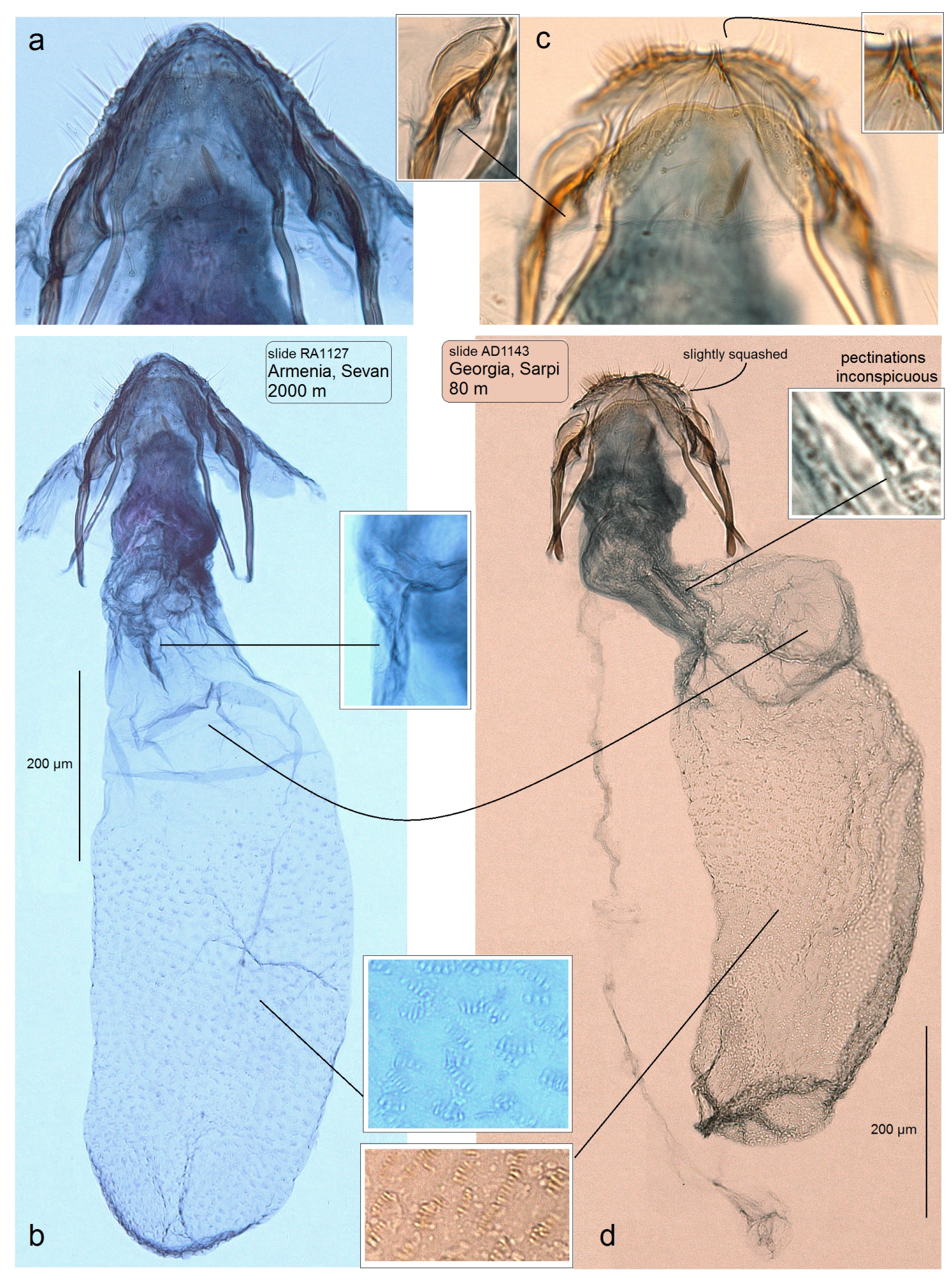
3.1. Taxonomic Account: A Description of a New Species
3.2. Molecular Account: The Caucasian Stigmella colchica sp. nov. vs. the European S. plagicolella
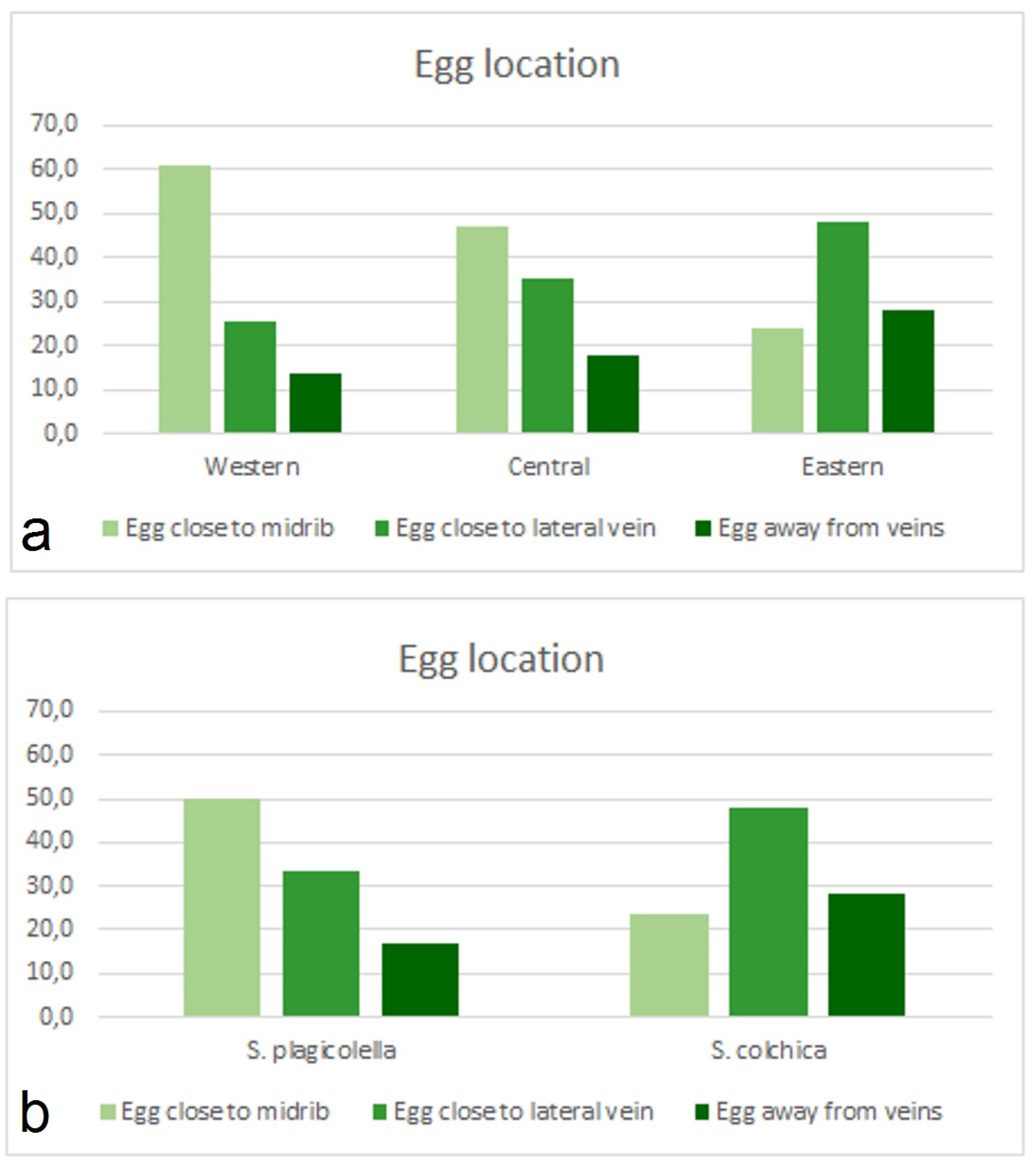
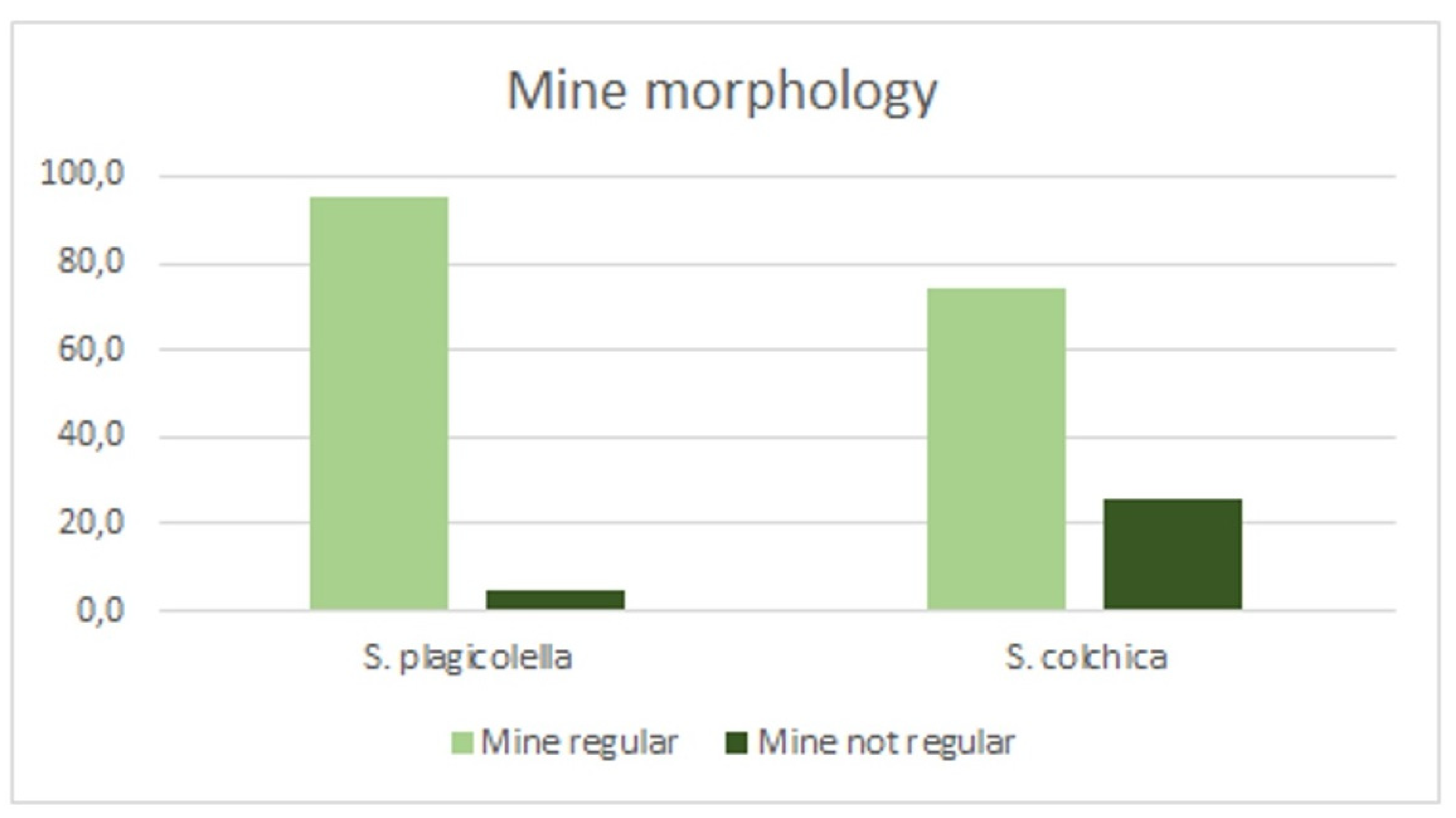
3.3. Statistical Analysis of Leaf-Mine Samples
- (1)
- Stigmella plagicolella, a sample of 246 leaf mines from GREAT BRITAIN, London (the type locality of S. plagicolella), 51°30′27.2″ N 0°25′39.3″ W, 8 August 2023, leg. J.R. Stonis & S.R. Hill;
- (2)
- S. plagicolella, a sample of 52 leaf mines from CROATIA, Rovinj, 45°04′27.1″ N 13°39′05.2″ E, 11 August 2023, leg. J.R. Stonis & S.R. Hill;
- (3)
- S. plagicolella, a sample of 318 leaf mines from CROATIA, Zagreb (Jarun), 45°46′38.9″ N 15°55′47.2″ E, 19–20 August 2023, leg. J.R. Stonis & S.R. Hill;
- (4)
- S. plagicolella, a sample of 366 leaf mines from BULGARIA, Sofia, 42°40′40.7″ N 23°18′52.1″ E, 23 August 2023, leg. S.R. Hill & J.R. Stonis;
- (5)
- S. plagicolella, a sample of 273 leaf mines from BULGARIA, 43°12′45.1″ N 27°56′23.4″ E, 24–30 August 2023, leg. S.R. Hill & J.R. Stonis.
- (6)
- S. colchica sp. nov., a sample of 279 leaf mines from GEORGIA, Kutaisi, 42°16′32.6″ N 42°42′36.5″ E–42°17′03.1″ N 42°43′42.1″ E, 25 July 2023, leg. S.R. Hill & J.R. Stonis;
- (7)
- S. colchica sp. nov., a sample of 60 leaf mines from GEORGIA, Sairme, 41°54′19.2″ N 42°44′56.5″ E, ca 1050 m, 15 July 2023, leg. J.R. Stonis;
- (8)
- S. colchica sp. nov., a sample of 333 leaf mines from GEORGIA, Samtredia, 42°09′47.6″ N 42°20′34.2″ E, 24 July 2023, leg. S.R. Hill & J.R. Stonis;
- (9)
- S. colchica sp. nov., a sample of 350 leaf mines from GEORGIA, Adjara AR, Kobuleti, 41°49′47.8″ N 41°46′43.9″ E, ca. 10 m, 23 July 2023, leg. S.R. Hill & J.R. Stonis;
- (10)
- S. colchica sp. nov., a sample of 141 leaf mines from GEORGIA, Batumi, 41°38′21.9″ N 41°37′44.0″ E–41°42′38.7″ N 41°43′42.1″ E, ca. 25 m, 17–23 July 2023, leg. S.R. Hill & J.R. Stonis;
- (11)
- S. colchica sp. nov., a sample of 51 leaf mines from GEORGIA, Adjara AR, SE of Batumi, Mirveti, 41°31′36.4″ N 41°42′58.6″ E, ca. 150 m and Machakhela N.P., Zeda Chkhutuneti, 41°29′05.9″ N 41°51′28.4″ E, ca. 600 m, 18 July 2023, leg. J.R. Stonis;
- (12)
- S. colchica sp. nov., a sample of 396 leaf mines from GEORGIA, Adjara AR, Sarpi (type locality of S. colchica sp. nov.), 41°31′26.2″ N 41°33′02.1″ E, ca. 80 m, 19–20 July 2023, leg. S.R. Hill & J.R. Stonis;
- (13)
- S. colchica sp. nov., a sample of 111 leaf mines from TÜRKIYE, Karadeniz Bölgesi, Artvin Province, Hopa, 41°23′39.4″ N 41°25′02.6″ E, ca. 5 m, and Üçkardeş, 41°29′35.7″ N 41°32′24.2″ E, ca. 70 m, 20 July 2023, leg. J.R. Stonis & S.R. Hill.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wheeler, Q.; Bourgoin, T.; Coddington, J.; Gostony, T.; Hamilton, A.; Larimer, R.; Polaszek, A.; Schauff, M.; Solis, M.A. Nomenclatural benchmarking: The role of digital typification and telemicroscopy. ZooKeys 2012, 209, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, Q.D.; Knapp, S.; Stevenson, D.W.; Stevenson, J.; Blum, S.D.; Boom, B.M.; Borisy, G.G.; Buizer, J.L.; de Carvalho, M.R.; Cibrian, A.; et al. Mapping the biosphere: Exploring species to understand the origin, organization, and sustainability of biodiversity. Syst. Biodivers. 2012, 10, 1–20. [Google Scholar] [CrossRef]
- Johansson, R.; Nielsen, E.S.; van Nieukerken, E.J.; Gustafsson, B. The Nepticulidae and Opostegidae (Lepidoptera) of north west Europe. Fauna Entomol. Scand. 1990, 23, 1–739. [Google Scholar]
- Regier, J.C.; Mitter, C.; Kristensen, N.P.; Davis, D.R.; van Nieukerken, E.J.; Rota, J.; Simonsen, T.J.; Mitter, K.T.; Kawahara, A.Y.; Yen, S.-H.; et al. A molecular phylogeny for the oldest (nonditrysian) lineages of extant Lepidoptera, with implications for classification, comparative morphology and life-history evolution. Syst. Entomol. 2015, 40, 671–704. [Google Scholar] [CrossRef]
- Doorenweerd, C.; van Nieukerken, E.J.; Hoare, R.J.B. Phylogeny, classification and divergence times of pygmy leaf-mining moths (Lepidoptera: Nepticulidae): The earliest lepidopteran radiation on Angiosperms? Syst. Entomol. 2017, 42, 267–287. [Google Scholar] [CrossRef]
- Stonis, J.R.; Remeikis, A.; Diškus, A. Neotropical Nepticulidae (a Pictorial Monograph Introducing an Electronic Identification Tool); Nature Research Centre: Vilnius, Lithuania, 2022; 363p. [Google Scholar]
- Dobrynina, V.; Stonis, J.R.; Diškus, A.; Solis, M.A.; Baryshnikova, S.V.; Young-Min, S. Global Nepticulidae, Opostegidae, and Tischeriidae (Lepidoptera): Temporal dynamics of species descriptions and their authors. Zootaxa 2022, 5099, 450–474. [Google Scholar] [CrossRef]
- Van Nieukerken, E.J.; Doorenweerd, C.; Hoare, J.R.; Davis, D.R. Revised classification and catalogue of global Nepticulidae and Opostegidae (Lepidoptera, Nepticuloidea). ZooKeys 2016, 628, 65–246. [Google Scholar] [CrossRef]
- Stonis, J.R.; Remeikis, A.; Dobrynina, V.; Mey, W. Collecting in the Northern Andean Páramo revealed purple iridescent pygmy moths of the little known Andean endemic Brachinepticula (Nepticulidae). Zootaxa 2023, 5227, 328–340. [Google Scholar] [CrossRef]
- Scoble, M.J. A revised cladistic classification of the Nepticulidae (Lepidoptera) with descriptions of new taxa mainly from South Africa. Transvaal Mus. Monogr. 1983, 2, 1–105. Available online: https://hdl.handle.net/10520/AJA090799001_84 (accessed on 2 September 2023).
- Puplesis, R. The Nepticulidae of Eastern Europe and Asia: Western, Central and Eastern Parts; Backhuys Publishers: Leiden, The Netherlands, 1994; 291p. [Google Scholar]
- Puplesis, R.; Robinson, G.S. A review of the Central and South American Nepticulidae (Lepidoptera) with special reference to Belize. Bull. Nat. Hist. Mus. Lond. 2000, 69, 3–114. Available online: https://www.biodiversitylibrary.org/page/40885664#page/5/mode/1up (accessed on 6 January 2024).
- Puplesis, R.; Diškus, A. The Nepticuloidea & Tischerioidea (Lepidoptera)—A Global Review, with Strategic Regional Revisions; Lututė Publishers: Kaunas, Lithuania, 2003; 512p. [Google Scholar]
- Stonis, J.R.; Remeikis, A.; Diškus, A.; Navickaitė, A. Documenting new and little known leaf-mining Nepticulidae from middle and southwestern areas of the Asian continent. Zootaxa 2020, 4881, 401–452. [Google Scholar] [CrossRef] [PubMed]
- Stonis, J.R.; Diškus, A. Diagnostics of Mines of the Lithuanian Nepticulidae. An Identification Tool. 2022. (In Lithuanian). Available online: https://leafmines.info (accessed on 10 January 2024).
- Van Nieukerken, E.J. A taxonomic revision of the Western Palaearctic species of the subgenera Zimmermannia Hering and Ectoedemia Busck s. str. (Lepidoptera, Nepticulidae), with notes on their phylogeny. Tijdschr. Entomol. 1985, 128, 1–164. Available online: https://biostor.org/reference/61076 (accessed on 2 September 2023).
- Van Nieukerken, E.J.; Doorenweerd, C.; Nishida, K.; Snyers, C. New taxa, including three new genera show uniqueness of Neotropical Nepticulidae (Lepidoptera). ZooKeys 2016, 628, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Van Nieukerken, E.J. A provisional phylogenetic check-list of the western Palaearctic Nepticulidae, with data on hostplants (Lepidoptera). Entomol. Scand. 1986, 17, 1–27. [Google Scholar] [CrossRef]
- Remeikis, A.; Diškus, A.; Stonis, J.R. Preliminary trophic analysis of leaf-mining Nepticulidae: Pronounced feeding strategies in Middle and South America? Biologija 2016, 62, 207–214. [Google Scholar] [CrossRef]
- Stonis, J.R.; Remeikis, A.; Diškus, A.; Baryshnikova, S.V.; Solis, M.A. What are the smallest moths (Lepidoptera) in the world? Zootaxa 2021, 4942, 269–289. [Google Scholar] [CrossRef]
- Kuznetzov, V.I.; Puplesis, R. Fam. Nepticulidae (Stigmellidae)—Pygmy Moths, Lepidoptera, 3. In Insects and Mites—Pests of Agricultural Plants; Kuznetzov, V.I., Ed.; Nauka Publishers: St. Petersburg, Russia, 1994; pp. 12–23. (In Russian) [Google Scholar]
- Stonis, J.R.; Remeikis, A.; Diškus, A.; Orlovskytė, S.; Vargas, S.A.; Solis, M.A. A new leafmining pest of guava: Hesperolyra guajavifoliae sp. nov., with comments on the diagnostics of the endemic Neotropical genus Hesperolyra van Nieukerken (Lepidoptera, Nepticulidae). ZooKeys 2019, 900, 87–110. [Google Scholar] [CrossRef]
- Stonis, J.R.; Diškus, A.; Fernández-Alonso, J.L.; Remeikis, A.; Solis, M.A. New leaf-mining Nepticulidae (Lepidoptera): Potential pests of aromatic Lamiaceae plants from equatorial Andes. Zootaxa 2021, 4926, 363–383. [Google Scholar] [CrossRef]
- Puplesis, R.K. New species of the nepticulid moths (Lepidoptera, Nepticulidae) from southern Far East and Tadzhikistan. Proc. Zool. Inst. Russ. Acad. Sci. 1985, 134, 59–72. (In Russian) [Google Scholar]
- Heyerdahl, R.; Dutcher, J.D. Management of the pecan serpentine leafminer (Lepidoptera: Nepticulidae). J. Econ. Entomol. 1985, 78, 1121–1124. [Google Scholar] [CrossRef]
- Alford, D.V. Pests of Ornamental Trees, Shrubs and Flowers. A Colour Handbook; CRC Press: London, UK, 2012; 480p. [Google Scholar] [CrossRef]
- Laštůvka, A.; Laštůvka, Z. Nepticulidae Mitteleuropas. Ein illustrierter Begleiter (Lepidoptera); Konvoj Publishers: Brno, Czechia, 1997; 230p. [Google Scholar]
- Stonis, J.R.; Dobrynina, V.; Remeikis, A.; Diškus, A.; Orlovskytė, S.; Kalashian, M.Y. The First Attempt to Assess the Taxonomic Diversity of the Nepticulidae of Armenia Resulted in the Discovery of New Species and Cryptic Taxa in the Caucasus. Diversity 2024, 16, 60. [Google Scholar] [CrossRef]
- Navickaitė, A.; Diškus, A.; Stonis, J.R. An updated checklist of Nepticulidae (Lepidoptera) of the Crimea, Sub-Mediterranean SE Europe. Zootaxa 2014, 3847, 151–202. [Google Scholar] [CrossRef] [PubMed]
- Rajaei, H.; Aarvik, L.; Arnscheid, W.R.; Baldizzone, G.; Bartsch, D.; Bengtsson, B.Å.; Bidzilya, O.; Buchner, P.; Buchsbaum, U.; Buszko, J.; et al. Catalogue of the Lepidoptera of Iran. Integr. Syst. Stutt. Contrib. Nat. Hist. 2023, 6, 121–459. [Google Scholar] [CrossRef]
- Stainton, H.T. Insecta Britannica. Lepidoptera: Tineina; Lovell Reeve: London, UK, 1854; 313p. [Google Scholar]
- Orlovskytė, S.; Dobrynina, V.; Stonis, J.R. Unexpected mitotype diversity of Simplimorpha promissa (Lepidoptera: Nepticulidae) in Ukraine and Armenia revealing a possible cryptic taxon. Zootaxa 2023, 5336, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2020, 21, 609–620. [Google Scholar] [CrossRef]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP v6: DNA Sequence Polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Clement, M.; Snell, Q.; Walke, P.; Posada, D.; Crandall, K. TCS: Estimating gene genealogies. In Proceedings of the 16th International Parallel and Distributed Processing Symposium, Ft. Lauderdale, FL, USA, 15–19 April 2002; p. 7. [Google Scholar] [CrossRef]
- Dean, A.G.; Sullivan, K.M.; Soe, M.M. OpenEpi: Open Source Epidemiologic Statistics for Public Health. Available online: http://OpenEpi.com (accessed on 2 September 2023).
- Fleiss, J.L.; Levin, B.; Paik, M.C. Statistical Methods for Rates and Proportions, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; 800p. [Google Scholar]
- G-Test Calculator. Available online: https://elem.com/~btilly/effective-ab-testing/g-test-calculator.html (accessed on 12 August 2023).
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data system (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Li, M.; Zhu, J.; Lu, W.; Li, J.; Zheng, B. Descriptions of Pedesta wangi sp. nov. and the female of P. xiaoqingae (Huang & Zhan, 2004) from China (Lepidoptera: Hesperiidae). Zootaxa 2019, 4700, 356–364. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; 514p. [Google Scholar]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef] [PubMed]
- Khomphimai, N.; Samranthin, C.; Chaianunporn, K.; Kutanan, W.; Chaianunporn, T. Mitochondrial DNA diversity of the lemon emigrant butterfly Catopsilia pomona Fabricius (Lepidoptera: Pieridae) in Khon Kaen Province, Thailand. Songklanakarin J. Sci. Technol. 2020, 42, 573–584. [Google Scholar] [CrossRef]
- Mayoke, A.; Muya, S.M.; Bateta, R.; Mireji, P.O.; Okoth, S.O.; Onyoyo, S.G.; Auma, J.E.; Ouma, J.O. Genetic diversity and phylogenetic relationships of tsetse fies of the palpalis group in Congo Brazzaville based on mitochondrial cox1 gene sequences. Parasit. Vectors 2020, 13, 253. [Google Scholar] [CrossRef]
- Radžiutė, S.; Orlovskytė, S.; Būdienė, J.; Blažytė-Čereškienė, L.; Bumbulytė, G.; Būda, V. Stored product pest Nemapogon variatella (Clemens): Identification of sex pheromone, putative antagonist and mitotype characterization. J. Appl. Entomol. 2023, 147, 857–867. [Google Scholar] [CrossRef]
- Teodoro, S.S.A.; da Silva Cortinhas, M.C.; Proietti, M.C.; Costa, R.C.; Dumont, L.F.C. High genetic connectivity among pink shrimp Farfantepenaeus paulensis (Pérez-Farfante, 1967) groups along the south-southeastern coast of Brazil. Estuar. Coast. Shelf Sci. 2020, 232, 106488. [Google Scholar] [CrossRef]
- Dou, C.; Muhammad, F.; Liu, L.; Gong, L.; Chen, Y.; Guo, B.; Lü, Z. Population diversity of Cistopus indicus inferred from mitochondrial DNA (cytochrome b) variation from China and Vietnam. Molluscan Res. 2019, 40, 1–7. [Google Scholar] [CrossRef]
- Garg, R.K.; Mishra, V. Molecular insights into the genetic and haplotype diversity among four populations of Catla catla from Madhya Pradesh revealed through mtDNA cyto b gene sequences. J. Genet. Eng. Biotechnol. 2018, 16, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Mohd Yasin, N.A.; Kasim, N.S.; Mat Jaafar, T.N.A.; Mat Piah, R.; Mohd Arshaad, W.; Mohd Nor, S.A.; Habib, A.; Tan, M.P. Population genetics of longtail tuna (Thunnus tonggol) (Bleeker, 1851) from East Malaysia based on mitochondrial DNA D-loop marker. UMT JUR 2020, 2, 49–56. [Google Scholar] [CrossRef]
- Suzuki, N.; Abramov, A.V.; Amaike, Y.; Nishita, Y.; Masuda, R. Phylogeography of the Altai weasel (Carnivora: Mustelidae: Mustela altaica) based on an analysis of mitochondrial control-region haplotypes. Biol. J. Linn. Soc. 2023, 138, 274–281. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000; pp. 59–60. [Google Scholar]
- Walker, T.J. Migrating Lepidoptera: Are butterflies better than moths? Fla. Entomol. 1980, 63, 79–98. [Google Scholar] [CrossRef]



















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stonis, J.R.; Diškus, A.; Orlovskytė, S.; Dobrynina, V. Revealing a Novel Potential Pest of Plum Trees in the Caucasus: A Species Resembling the European Leaf-Mining Stigmella plagicolella, Nepticulidae. Insects 2024, 15, 198. https://doi.org/10.3390/insects15030198
Stonis JR, Diškus A, Orlovskytė S, Dobrynina V. Revealing a Novel Potential Pest of Plum Trees in the Caucasus: A Species Resembling the European Leaf-Mining Stigmella plagicolella, Nepticulidae. Insects. 2024; 15(3):198. https://doi.org/10.3390/insects15030198
Chicago/Turabian StyleStonis, Jonas R., Arūnas Diškus, Svetlana Orlovskytė, and Viktorija Dobrynina. 2024. "Revealing a Novel Potential Pest of Plum Trees in the Caucasus: A Species Resembling the European Leaf-Mining Stigmella plagicolella, Nepticulidae" Insects 15, no. 3: 198. https://doi.org/10.3390/insects15030198