
Thrips Species Composition in Ontario Greenhouse Floriculture: Innovative Identification Tools and Implications for Integrated Pest Management

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Thrips Collection
2.2. Thrips Identification and Development of a Simplified Key
2.3. Data Analyses
3. Results
3.1. Thrips Species Found in Floriculture Greenhouses in Ontario in Summer 2016 and 2017
3.2. Thrips Species Composition in Greenhouses Revisited in Summer 2017 and 2019
3.3. Thrips Species Composition in Greenhouses Revisited in Fall 2018
3.4. Thrips Populations or Outbreaks Identified after 2016 in Ontario
3.5. Simple Key to Thrips Species Found in Ontario Greenhouses for Growers
- a. Head and pronotum tan or yellow; abdomen tan, yellow, or light to medium brown (GO TO STEP 2).b. Head, pronotum, and abdomen brown to black in colour; head and pronotum may be lighter brown than abdomen, but not yellow (GO TO STEP 7).
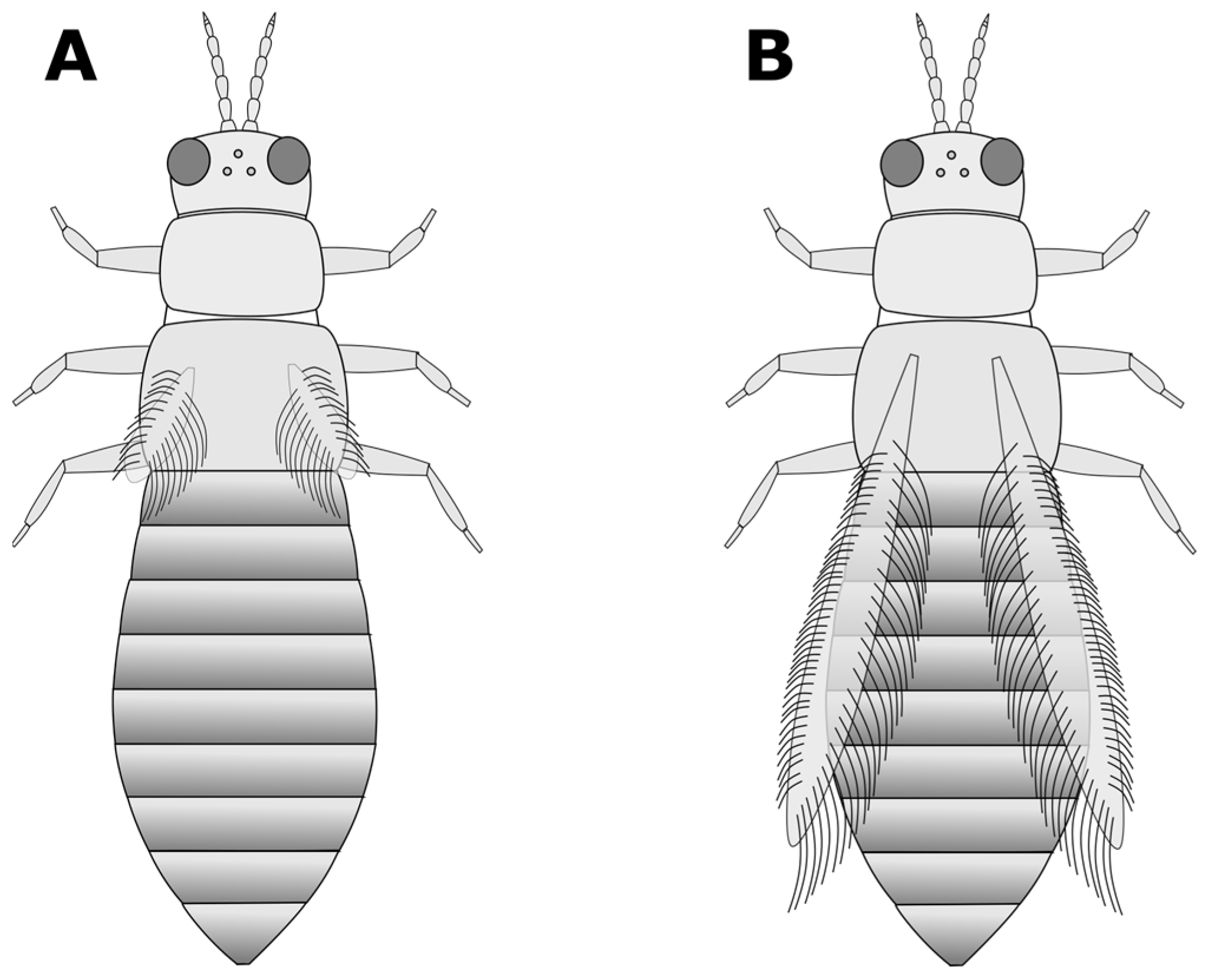
- a. Very short wings, shorter than the width of the body (Figure 3A); foliar feeding damage typically on lower leaves, usually only found on chrysanthemums and gloxinias:Thrips nigropilosus, “chrysanthemum thrips”, wingless form.(Note: both winged and wingless forms may be present in the same population).b. Long fringed wings extending nearly the full length of the body (Figure 3B) (GO TO STEP 3).
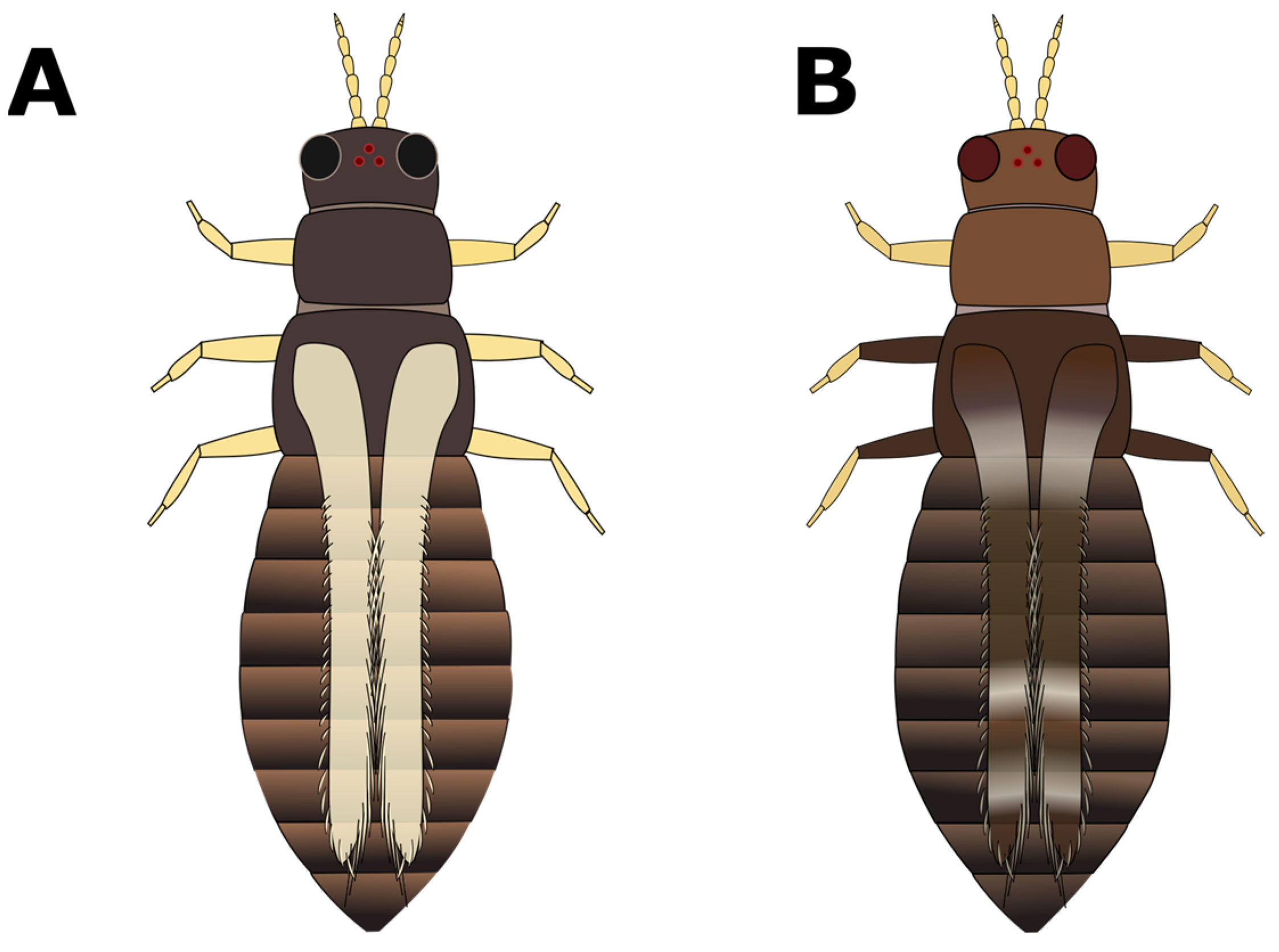
- a. Pronotum has two pairs of long coarse hairs on both the top and bottom of the pronotum (Figure 4A); ocelli are red:Frankliniella occidentalis “western flower thrips” (most common).(May also be other Frankliniella species, such as F. tritici, F. bispinosa, or F. schultzei (pale forms). Differentiating between Frankliniella species requires a compound microscope and advanced identification skills.)b. No long coarse hairs on the top of the pronotum. There may be hairs on the bottom edge of the pronotum; ocelli may or may not be red (Figure 4B–D) (GO TO STEP 4).
- a. Ocelli are grey; two pairs of long coarse hairs on the bottom edge of the pronotum (Figure 4B):Thrips tabaci, “onion thrips”.b. Ocelli are red; coarse hairs on the bottom of the pronotum may be long or short (Figure 4B–D) (GO TO STEP 5).
- a. Three pairs of coarse hairs on the bottom of the pronotum, outer two pairs of hairs of equal length, which are distinctly longer than the inner pair (Figure 5A); wings are pale or absent; foliar feeding damage, typically on lower leaves; usually only found on chrysanthemums and gloxinias:Thrips nigropilosus, “chrysanthemum thrips”, winged form.(If found on crops other than chrysanthemum and gloxinia, it may be T. palmi, which is not present in Canada but may be intercepted on plant material imported from tropical regions, including Florida, Mexico, and Central America. Differentiating between T. nigropilosus and T. palmi requires a compound microscope and advanced identification skills. It could also be male T. parvispinus or T. setosus: see Step 10 for identification of females if dark-coloured thrips are also present.)b. Coarse dark hairs on the bottom of the pronotum are short and may be difficult to see (Figure 5B,C); small body size compared to other common thrips species; wings are grey or have black markings; usually found on tropicals (GO TO STEP 6).
- a. Head and body yellow; wings black with distinct pale band in the middle (Figure 6A); hairs on the bottom of the pronotum are short and fine and often difficult to see with a dissection microscope (Figure 5C); usually found on tropicals:Chaetanaphothrips orchidii, “orchid thrips”.(This species is not present in Canada but widespread in tropical and sub-tropical regions, including California and Florida; it may be intercepted on imported plant material.)b. Head and body yellow; wings are grey with some paler sections but not distinctly banded (Figure 6B); coarse dark hairs on the bottom of the pronotum are short, middle pair distinctly longer than the others (Figure 5B) (may be difficult to see depending on the quality of your microscope); usually found on tropicals; distinctive feeding damage causes distortions at growing points, resembling broad mite damage:Scirtothrips dorsalis, “chilli thrips”.(This species is not present in Canada, but present in Florida, Texas, Mexico, and the Caribbean; it may be intercepted on imported plant material.)
- a. Pronotum has long coarse hairs (Figure 4A–C). Note: hairs may be difficult to see on black thrips—may be easier to see in side-view (GO TO STEP 8).b. No long coarse hairs on the pronotum (Figure 4D); front legs entirely yellow (GO TO STEP 11).
- a. Long coarse hairs on both top and bottom of pronotum (Figure 4A); no red pigment visible between the segments; wings uniformly pale brown:Frankilinella fusca, “tobacco thrips”, winged form (most common).(May also be F. occidentalis, dark morph (usually seen in fall and winter), F. intonsa (not known from Ontario, but present in British Columbia), or F. schultzei (not present in Canada but present in Florida, Central America, and the Caribbean, may be intercepted on imported plant materials)).b. No long coarse hairs on top of pronotum, but bottom has two pairs of long coarse hairs (Figure 4B); red pigment may or may not be visible between segments; wings may be uniformly pale or dark with pale bands (GO TO STEP 9).
- a. Head and body light to medium brown, never black, uniform in colour; grey ocelli; wings uniformly pale in colour (Figure 7A):Thrips tabaci, “onion thrips”, dark morph.b. Abdomen dark brown to black; head and pronotum either medium brown or black; red ocelli; light patches at the top of the wings (visible on dry specimens) (Figure 7B,C) (GO TO STEP 10).
- a. Head and body black; red ocelli (may be difficult to see); red pigmentation often visible between body segments; back legs yellow with black femurs, femurs on front legs dusky at base but not distinctly black; abdomen widest at the top and narrowing toward tip (Figure 7B):Echinothrips americanus, “poinsettia thrips” (most common).(Dichromothrips corbetti is similar in appearance to Echinothrips, but is only present on orchid crops, and does not have hairs on the top or bottom of the pronotum.)b. Head and pronotum medium to dark brown, abdomen darker than head; ocelli bright red and easily visible; no red pigmentation between segments; abdomen widest in the middle (Figure 7C):Thrips parvispinus, “pepper thrips”, females, or Thrips setosus, “Japanese flower thrips”, females.(The males of both species are pale yellow. Neither species is established in Canada but present in some US states; they may be intercepted on imported plant material. In northern latitudes, T. parvispinus is typically intercepted on tropicals, and T. setosus has been found on Hydrangea. Differentiating between these two species requires a compound microscope and advanced identification skills.)
- a. All legs entirely yellow; head and pronotum as dark as or darker than the abdomen; wings uniform in colour and paler than the body (visible on dry specimens) (Figure 8A):Heliothrips haemorrhoidalis, “greenhouse thrips”b. Front legs yellow, brown femurs on back legs; head and pronotum often paler than abdomen; light bands at the top and tips of the wings (visible on dry specimens) (Figure 8B):Hercinothrips femoralis, “banded greenhouse thrips”.
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, T. Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxford, UK, 1997; ISBN 0851991785. [Google Scholar]
- van Dijken, F.R.; Dik, M.T.A.; Gebala, B.; de Jong, J.; Mollema, C. Western flower thrips (Thysanoptera: Thripidae) effects on chrysanthemum cultivars: Plant growth and leaf scarring in nonflowering plants. J. Econ. Entomol. 1994, 87, 1312–1317. [Google Scholar] [CrossRef]
- Lewis, T. Pest Thrips in Perspective. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxford, UK, 1997; pp. 1–13. [Google Scholar]
- Ullman, D.E.; Sherwood, J.L.; German, T.L. Thrips as Vectors of Plant Pathogens. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxford, UK, 1997; pp. 539–565. ISBN 0851991785. [Google Scholar]
- Okuda, M.; Fuji, S.; Okuda, S.; Sako, K.; Iwanami, T. Evaluation of the potential of thirty two weed species as infection sources of Impatiens necrotic spot virus. J. Plant Pathol. 2010, 92, 357–361. [Google Scholar] [CrossRef]
- Childers, C.C. Feeding and Oviposition Injuries to Plants. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: New York, NY, USA, 1997; pp. 505–538. [Google Scholar]
- Summerfield, A. Biocontrol thriving in Canadian greenhouses. Greenh. Can. 2019, 39, 28–30. [Google Scholar]
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agric. For. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Buitenhuis; Murphy, G.; Shipp, L. Aphis gossypii Glover, Melon/Cotton Aphid, Aulacorthum solani (Kaltenbach), Foxglove Aphid, and Other Arthropod Pests in Greenhouse Crops. In Biocontrol Programmes in Canada 2001–2012; Mason, P.G., Gillespie, D.R., Eds.; CAB International: Oxford, UK, 2013; pp. 98–107. [Google Scholar]
- Murphy, G.; Ontario Ministry of Agriculture, Food and Rural Affairs, Vineland Station, ON, Canada. Personal Communication, 2015.
- Frank, C.E.; Skinner, M. Common Greenhouse Pests: Aphids; University of Vermont, Entomology Research Laboratory, 2013. Available online: https://www.uvm.edu/~entlab/Greenhouse%20IPM/Pests&Beneficials/Aphids.pdf (accessed on 1 March 2024).
- White, J. Whiteflies in the Greenhouse | University of Kentucky Entomology; University of Kentucky Cooperative Extension Service, 2013. Available online: https://entomology.ca.uky.edu/ef456 (accessed on 1 March 2024).
- Buitenhuis, R. Grower Guide: Quality Assurance of Biocontrol Products; Vineland Research and Innovation Centre, 2017. Available online: https://www.vinelandresearch.com/wp-content/uploads/2020/02/Grower-Guide.pdf (accessed on 1 March 2024).
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef]
- MacIntyre-Allen, J.K.; Scott-Dupree, C.D.; Tolman, J.H.; Harris, C.R. Evaluation of sampling methodology for determining the population dynamics of onion thrips (Thysanoptera: Thripidae) in Ontario onion fields. J. Econ. Entomol. 2005, 98, 2272–2281. [Google Scholar] [CrossRef] [PubMed]
- Shipp, J.L.; Gillespie, D.R.; Fry, K.M.; Ferguson, G.M. Echinothrips Americanus (Morgan), Frankliniella occidentalis (Pergande), Western Flower Thrips, and Thrips tabaci Lindeman, Onion Thrips (Thysanoptera: Thripidae). In Biocontrol Programmes in Canada 1981–2000; Mason, P.G., Huber, J.T., Eds.; CAB International: Oxford, UK, 2002; pp. 115–119. [Google Scholar]
- Broadbent, A.B.; Allen, W.R.; Foottit, R.G. The association of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) with greenhouse crops and the tomato spotted wilt virus in Ontario. Can. Entomol. 1987, 119, 501–503. [Google Scholar] [CrossRef]
- Ontario Ministry of Agriculture Food and Rural Affairs. Vegetable Crop Protection Guide (Publication 838); Ontario Ministry of Agriculture Food and Rural Affairs, 2021; ISBN 1800265565. Available online: https://www.omafra.gov.on.ca/english/crops/pub838/pub838.pdf (accessed on 1 March 2024).
- Chittenden, F.H. Control of the Onion Thrips. Farmers’ Bull. United States Dep. Agric. 1919, 1–16. [Google Scholar]
- Bryant, T.; Reay-Jones, F.P.F. Integrated Pest Management: Concepts and Strategies Basics of IPM Steps In Developing an IPM Program; Land-Grant Press by Clemson Extension, 2020. Available online: https://lgpress.clemson.edu/publication/integrated-pest-management-concepts-and-strategies/ (accessed on 1 March 2024).
- Brown, W. The Ontario Greenhouse Floriculture Industry; Ontario Ministry of Agriculture, Food and Rural Affairs, 2003. Available online: https://omafra.gov.on.ca/english/crops/hort/greenhouse.html (accessed on 1 March 2024).
- Lewis, T. Movement, Migration and Dispersal. In Thrips, Their Biology, Ecology and Economic Importance; Academic Press: London, UK, 1973; pp. 131–166. ISBN 0-12-447160-9. [Google Scholar]
- Murphy, G.; Jandricic, S. Reacting to a sudden summer pest threat. Greenh. Canada 2016, 36, 44–48. [Google Scholar]
- Summerfield, A. Integrated Pest Management of Onion Thrips (Thrips tabaci), an Emerging Pest of Greenhouse Floriculture Crops in Ontario. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2022. [Google Scholar]
- Stannard, L.J. The Thrips, or Thysanoptera, of Illinois. Illinois Nat. Hist. Surv. Bull. 1968, 29, 215–552. [Google Scholar] [CrossRef]
- Mound, L.A.; Kibby, G. Thysanoptera an Identification Guide, 2nd ed.; CAB International: Oxford, UK, 1998. [Google Scholar]
- Cluever, J.D.; Smith, H.A. A photo-based key of thrips (Thysanoptera) associated with horticultural crops in Florida. Florida Entomol. 2017, 100, 454–467. [Google Scholar] [CrossRef]
- North Carolina Cooperative Extension Service. Key to Eleven Species of Thrips Found in Greenhouses. In Insect and Related Pests of Fowers and Foliage Plants (Agricultural Extension Publication 136); Baker, J.R., Ed.; North Carolina Cooperative Extension Service, 1994; pp. 79–91. ISBN 9788527729833. Available online: https://content.ces.ncsu.edu/insect-and-related-pests-of-flowers-and-foliage-plants/thrips-found-on-flowers-and-foliage (accessed on 1 March 2024).
- Frantz, G.; Fasulo, T.R. Thrips Knowledgebase. Glades Crop Care. Available online: https://www.gladescropcare.com/thrips/pg1.html (accessed on 22 January 2024).
- Summerfield, A.; Jandricic, S. Simple Key to Important Thrips Pests of Canadian Greenhouses, 2nd ed.; 2021; Available online: http://greenhouseipm.org/wp-content/uploads/2021/06/Key-to-important-thrips-pests-of-Ontario-greenhouses_2ndEd_2021_HighRes.pdf (accessed on 1 March 2024).
- Pearsall, I.A. Daily flight activity of the western flower thrips (Thysan., Thripidae) in nectarine orchards in British Columbia, Canada. J. Appl. Entomol. 2002, 126, 293–302. [Google Scholar] [CrossRef]
- Environment Canada Historical Data—Station Results—Vineland. Available online: https://climate.weather.gc.ca/historical_data/search_historic_data_stations_e.html?searchType=stnName&timeframe=1&txtStationName=Vineland&searchMethod=contains&optLimit=yearRange&StartYear=2016&EndYear=2019&Year=2024&Month=1&Day=22&selRowPerPage=25 (accessed on 22 January 2024).
- Gleason, J.E.; Maw, E.; Summerfield, A.; Jandricic, S.E.; Brunet, B.M.T.; Centre, D.; Canada, A. First records of invasive agricultural pests Thrips parvispinus (Karny, 1922) and Thrips setosus Moulton, 1928 (Thysanoptera: Thripidae) in Canada. J. Entomol. Soc. Ontario 2023, 154, 1–12. [Google Scholar]
- Pizzol, J.; Nammour, D.; Rabasse, M.; Desneux, N.; Poncet, C.; Reynaud, P. Species and population dynamics of thrips occurring inside and outside greenhouses cultivated with roses in southern France. Int. J. Agric. Pol. Res. 2014, 2, 141–153. [Google Scholar]
- Bout, A.; Marchand, A.; Ziegler, M.; Disdier, M.; Reynaud, P.; Robert, F.; Crochard, D.; Pizzol, J.; Ris, N.; Malausa, T. Improving knowledge on thrips diversity and ecology in French greenhouses: Inputs of barcoding approaches for a better pest management. Acta Hortic. 2015, 1104, 395–400. [Google Scholar] [CrossRef]
- Bennison, J.; Seymour, P. A review of thrips species (other than western flower thrips ) and their control on strawberry. Agric. Hortic. Dev. Board 2020, 1, 1–23. Available online: https://projectbluearchive.blob.core.windows.net/media/Default/Horticulture/Desk%20study%20-%20Alternative%20thrips%20in%20strawberry.pdf (accessed on 1 March 2024).
- Nielsen, H.; Sigsgaard, L.; Kobro, S.; Jensen, N.L.; Jacobsen, S.K. Species composition of thrips (Thysanoptera: Thripidae) in strawberry high tunnels in Denmark. Insects 2021, 12, 208. [Google Scholar] [CrossRef]
- Ganaha-Kikumura, T.; Ohno, S.; Kijima, K.; Masumoto, M.; Maekado, N. Species composition of thrips (Thysanoptera: Thripidae) and spider mites (Acari: Tetranychidae) on cultivated chrysanthemum (Asteraceae) in Okinawa, southwestern Japan. Entomol. Sci. 2012, 15, 232–237. [Google Scholar] [CrossRef]
- Morsello, S.C.; Groves, R.L.; Nault, B.A.; Kennedy, G.G. Temperature and precipitation affect seasonal patterns of dispersing tobacco thrips, Frankliniella fusca, and onion thrips, Thrips tabaci (Thysanoptera: Thripidae) caught on sticky traps. Environ. Entomol. 2008, 37, 79–86. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Chen, M. Studies of thrips migratory flights in Israel. Acta Phytopathol. Entomol. Hungarica 2008, 43, 243–248. [Google Scholar] [CrossRef]
- Veres, A.; Toth, F.; Orosz, S.; Kristof, D.; Fetyko, K. Spatial analysis of greenhouse density in relation to western flower thrips (Frankliniella occidentalis), onion thrips (Thrips tabaci) and minute pirate bug (Orius spp.) populations in greenhouses. IOBC-WPRS Bull. 2008, 34, 129–132. [Google Scholar]
- Lewis, T. Flight and Dispersal. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: Oxford, UK, 1997; pp. 175–196. [Google Scholar]
- Buitenhuis, R.; Lee, W.; Summerfield, A.; Smitley, D. Thrips IPM in floriculture: Cutting dips to start clean. IOBC-WPRS Bull. 2019, 147, 130–135. [Google Scholar]
- Lewis, T. (Ed.) Jacobson Integrated Pest Management (IPM) in Glasshouses. In Thrips as Crop Pests; CAB International: Oxford, UK, 1997; pp. 639–666. ISBN 0 85199 178 5. [Google Scholar]
- Jensen, S.E. Insecticide resistance in the western flower thrips, Frankliniella occidentalis. Integr. Pest Manag. Rev. 2000, 5, 131–146. [Google Scholar] [CrossRef]
- Wang, Z.H.; Gong, Y.J.; Jin, G.H.; Li, B.Y.; Chen, J.C.; Kang, Z.J.; Zhu, L.; Gao, Y.L.; Reitz, S.; Wei, S.E.J. Field-evolved resistance to insecticides in the invasive western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae) in China. Pest Manag. Sci. 2016, 72, 1440–1444. [Google Scholar] [CrossRef]
- Brodsgaard, H.F. Insecticide resistance in European and African strains of western flower thrips (Thysanoptera: Thripidae) tested in a new residue-on-glass test. J. Econ. Entomol. 1994, 87, 1141–1146. [Google Scholar] [CrossRef]
- Brodsgaard, H.F. Biological Control of Thrips on Ornamental Crops. In Biocontrol in Protected Culture; Heinz, K.M., Van Driesche, R.G., Parrella, M.P., Eds.; Ball Publishing: Batavia, NY, USA, 2004; pp. 253–264. [Google Scholar]
- Homem, R.A.; Buttery, B.; Richardson, E.; Tan, Y.; Field, L.M.; Williamson, M.S.; Emyr Davies, T.G. Evolutionary trade-offs of insecticide resistance—The fitness costs associated with target-site mutations in the nAChR of Drosophila melanogaster. Mol. Ecol. 2020, 29, 2661–2675. [Google Scholar] [CrossRef] [PubMed]
- Foster, S.P.; Denholm, I.; Poppy, G.M.; Thompson, R.; Powell, W. Fitness trade-off in peach-potato aphids (Myzus persicae) between insecticide resistance and vulnerability to parasitoid attack at several spatial scales. Bull. Entomol. Res. 2011, 101, 659–666. [Google Scholar] [CrossRef]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Acetamiprid resistance and fitness costs of melon aphid, Aphis gossypii: An age-stage, two-sex life table study. Pestic. Biochem. Physiol. 2021, 171, 104729. [Google Scholar] [CrossRef]
- Jackson, G.E.; Malloch, G.; McNamara, L.; Little, D. Grain aphids (Sitobion avenae) with knockdown resistance (kdr) to insecticide exhibit fitness trade-offs, including increased vulnerability to the natural enemy Aphidius ervi. PLoS ONE 2020, 15, e0230541. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef]
- Li, X.; Wan, Y.; Yuan, G.; Hussain, S.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Wu, Q. Fitness trade-off associated with spinosad resistance in Frankliniella occidentalis (Thysanoptera: Thripidae). J. Econ. Entomol. 2017, 110, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Reitz, S.R.; Yuan, H.; Lei, Z.; Paini, D.R.; Gao, Y. Pesticide-mediated interspecific competition between local and invasive thrips pests. Sci. Rep. 2017, 7, 40512. [Google Scholar] [CrossRef]
- Reitz, S.R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Florida Entomol. 2009, 92, 7–13. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, D.R. Biological control of thrips [Thysanoptera: Thripidae] on greenhouse cucumber by Amblyseius cucumeris. Entomophaga 1989, 34, 185–192. [Google Scholar] [CrossRef]
- Nielsen, M.C.; Davidson, M.M.; Butler, R.C. Predation rate of Thrips tabaci larvae by Neoseiulus cucumeris is influenced by prey density and presence of a host plant. New Zeal. Plant Prot. 2014, 67, 197–203. [Google Scholar] [CrossRef]
- Gulzar, S.; Wakil, W.; Shapiro-Ilan, D.I. Combined effect of entomopathogens against Thrips tabaci Lindeman (Thysanoptera: Thripidae): Laboratory, greenhouse and field trials. Insects 2021, 12, 456. [Google Scholar] [CrossRef] [PubMed]
- Cloyd, R.A.; Galle, C.L.; Keith, S.R. Compatibility of three miticides with the predatory mites Neoseiulus californicus McGregor and Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae). HortScience 2006, 41, 707–710. [Google Scholar] [CrossRef]
- You, Y.; Lin, T.; Wei, H.; Zeng, Z.; Fu, J.; Liu, X.; Lin, R.; Zhang, Y. Laboratory evaluation of the sublethal effects of four selective pesticides on the predatory mite Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae). Syst. Appl. Acarol. 2016, 21, 1506–1514. [Google Scholar] [CrossRef]
- Van Driesche, R.G.; Lyon, S.; Nunn, C. Compatibility of spinosad with predacious mites (Acari: Phytoseiidae) used to control western flower thrips (Thysanoptera: Thripidae) in greenhouse crops. Florida Entomol. 2006, 89, 396–401. [Google Scholar] [CrossRef]
- Herrick, N.J.; Cloyd, R.A. Direct and indirect effects of pesticides on the insidious flower bug (Hemiptera: Anthocoridae) under laboratory conditions. J. Econ. Entomol. 2017, 110, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Crops Sampled | 2016 | 2017 | 2018 | 2019 |
|---|---|---|---|---|---|
| 1 | Chrysanthemums | 5 | 3 | 3 | 6 |
| 2 | Gerbera | 4 | 1 | ||
| Mandevilla | 3 | ||||
| 3 | Chrysanthemum | 6 | 4 | ||
| Hibiscus | 7 | ||||
| 4 | Gerbera | 3 | |||
| 5 | Cut Chrysanthemums | 6 | 3 | ||
| 6 | Cut Chrysanthemums | 5 | |||
| Cut Gerbera | 4 | 3 | |||
| 7 | Ivy Geranium | 2 | |||
| New Guinea impatiens | 4 | ||||
| 8 | Chrysanthemums | 2 |
| Site | Crop | #of Dates | Total Thrips | WFT ± SE | OT ± SE | Other ± SE |
|---|---|---|---|---|---|---|
| 1 | Chrysanthemum (potted) | 5 | 258 | 97.0% ± 1.7 | 3.0% ± 1.7 | 0.0% ± 0.0 |
| 2 | Mandevilla | 3 | 310 | 97.2% ± 1.4 | 1.4% ± 1.4 | 1.4% ± 1.4 |
| Gerbera (potted) | 4 | 194 | 8.3% ± 2.5 | 89.8% ± 4.4 | 1.9% ± 1.9 | |
| 3 | Chrysanthemum (potted) | 6 | 207 | 72.6% ± 13.5 | 27.4% ± 13.5 | 0.0% ± 0.0 |
| Hibiscus | 7 | 299 | 99.4% ± 0.4 | 0.6% ± 0.4 | 0.0% ± 0.0 | |
| 4 | Gerbera (potted) | 3 | 423 | 17.5% ± 8.0 | 82.5% ± 8.0 | 0.0% ± 0.0 |
| 5 | Chrysanthemum (cut) | 6 | 292 | 86.4% ± 5.7 | 12.8% ± 5.8 | 0.8% ± 0.6 |
| 6 | Chrysanthemum (cut) | 5 | 177 | 61.8% ± 18.3 | 5.2% ± 3.2 | 33.0% ± 20.1 |
| Gerbera (cut) | 4 | 240 | 53.9% ± 14.3 | 45.1% ± 14.2 | 1.0% ± 0.6 | |
| 7 | Ivy Geranium | 2 | 36 | 90.0% ± 10.0 | 10.0% ± 10.0 | 0.0% ± 0.0 |
| New Guinea Impatiens | 4 | 165 | 83.4% ± 8.2 | 16.6% ± 8.2 | 0.0% ± 0.0 | |
| 8 | Chrysanthemum (potted) | 2 | 62 | 1.0% ± 1.0 | 99.0% ± 1.0 | 0.0% ± 0.0 |
| Total, all sites and crops | 2663 | 59.1% ± 10.8 | 37.9% ± 3.2 | 3.2% ± 2.6 |
| Species | Crop | Date |
|---|---|---|
| Bagnalliella yuccae (Hinds) | Yucca spp. * | December 2023 |
| Dichromothrips corbetti (Priesner) | Phalaenopsis orchid | July 2023 |
| Echinothrips americanus Morgan | Gerbera (cut) | November 2018 |
| Philodendron * | November 2022 | |
| Frankliniella fusca (Hinds) | Cyclamen | October 2018 |
| Cyclamen | October 2022 | |
| Frankliniella schultzei (Trybom) | Cactus * | November 2022 |
| Gynaikothrips uzeli (Zimmermann) | Ficus benjamina * | November 2022 |
| Hercinothrips femoralis (Reuter) | Unspecified ornamentals * | January 2017 |
| Gerbera (cut) | August 2018 | |
| Spring bedding plants | February 2019 | |
| Peperomia * | December 2022 | |
| Scirtothrips dorsalis Hood | Schefflera | December 2017 |
| Thrips nigropilosus Uzel | Cyclamen | October 2018 |
| Chrysanthemum (potted) | August 2020 | |
| Thrips parvispinus (Karny) | Mandevilla/Dipladenia * | October 2021 |
| Mandevilla/Dipladenia * | February 2022 | |
| Unspecified tropical ornamentals * | July 2022 | |
| Thrips setosus Moulton | Hydrangea * | April 2022 |
| Hydrangea * | October 2022 | |
| Thrips tabaci Lindeman | Osteospermum | May 2016 |
| Gerbera (potted) | July 2017 | |
| Chrysanthemum (cut) | July 2021 | |
| Cyclamen | October 2022 | |
| Cyclamen | November 2022 | |
| Anthurium * | November 2022 | |
| Primrose | January 2023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jandricic, S.E.; Summerfield, A.; Maw, H.E.L.; Brunet, B.M.T.; Buitenhuis, R. Thrips Species Composition in Ontario Greenhouse Floriculture: Innovative Identification Tools and Implications for Integrated Pest Management. Insects 2024, 15, 211. https://doi.org/10.3390/insects15030211
Jandricic SE, Summerfield A, Maw HEL, Brunet BMT, Buitenhuis R. Thrips Species Composition in Ontario Greenhouse Floriculture: Innovative Identification Tools and Implications for Integrated Pest Management. Insects. 2024; 15(3):211. https://doi.org/10.3390/insects15030211
Chicago/Turabian StyleJandricic, Sarah Elizabeth, Ashley Summerfield, H. Eric L. Maw, Bryan M. T. Brunet, and Rosemarije Buitenhuis. 2024. "Thrips Species Composition in Ontario Greenhouse Floriculture: Innovative Identification Tools and Implications for Integrated Pest Management" Insects 15, no. 3: 211. https://doi.org/10.3390/insects15030211
APA StyleJandricic, S. E., Summerfield, A., Maw, H. E. L., Brunet, B. M. T., & Buitenhuis, R. (2024). Thrips Species Composition in Ontario Greenhouse Floriculture: Innovative Identification Tools and Implications for Integrated Pest Management. Insects, 15(3), 211. https://doi.org/10.3390/insects15030211