
Bumblebee Foraging Dynamics and Pollination Outcomes for Cherry Tomato and Pear Varieties in Northern China

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Site
2.2. Bumblebee and Hormone Treatment for Cherry Tomatoes in Greenhouses
2.3. Monitoring of Temperature, Relative Humidity, and Bumblebee Foraging Behavior in Cherry Tomato Greenhouses
2.4. Investigation of the Effect of Bumblebee and Hormone Treatments on Cherry Tomatoes
2.5. Bumblebee Pollination, Hand Pollination, and Open Pollination in Pear Orchards
2.6. Monitoring of Temperature, Relative Humidity, and Bumblebee Foraging Behavior in Pear Orchards
2.7. Investigation of the Effects of the Three Pollination Methods on Pears
2.8. Data Analysis
3. Results
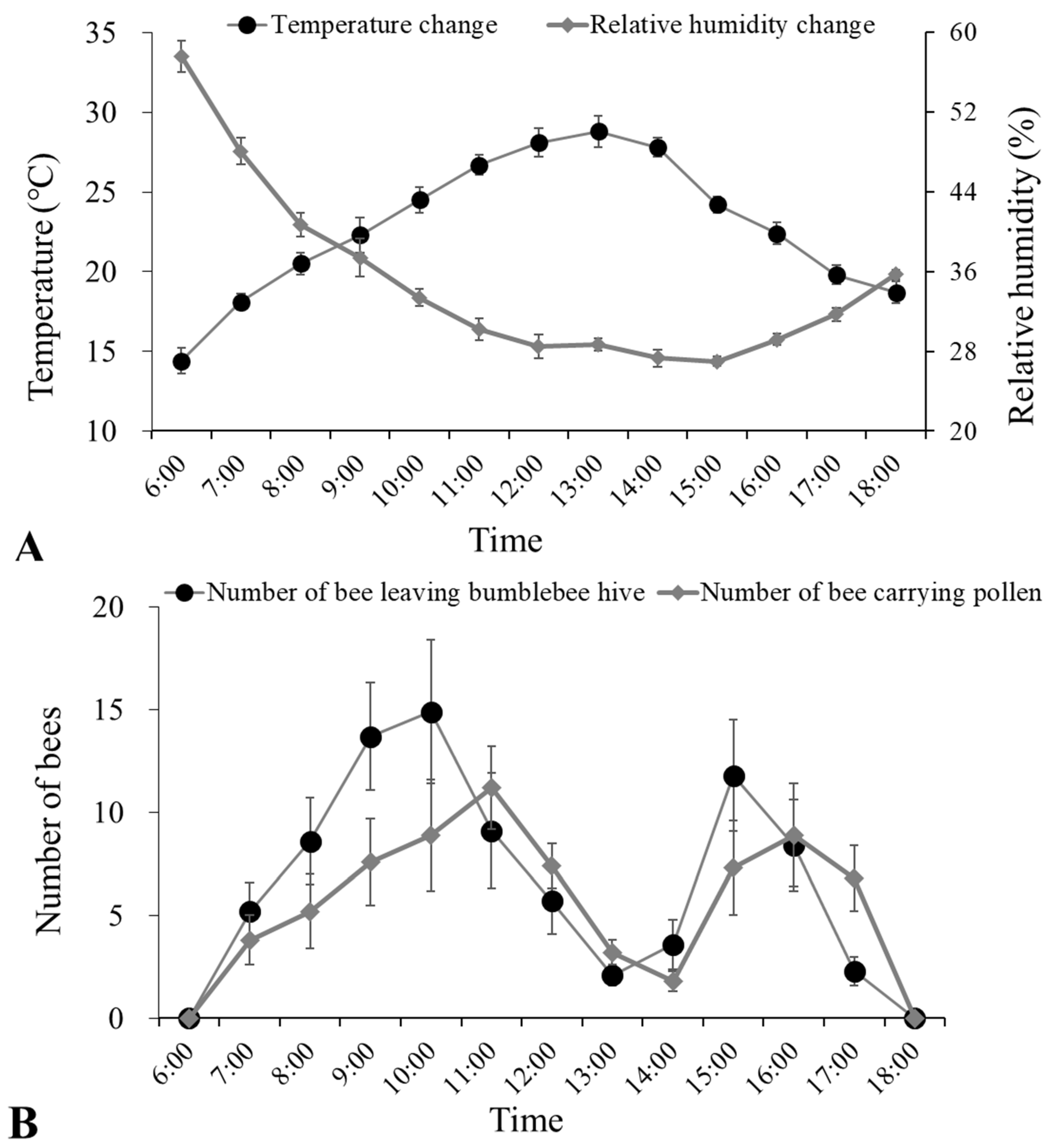
3.1. Bumblebee Foraging Activities in Greenhouses of Cherry Tomatoes
3.2. Bumblebee Foraging Activities in Pear Orchards
3.3. Foraging Behavior of Bumblebees Visiting Flowers
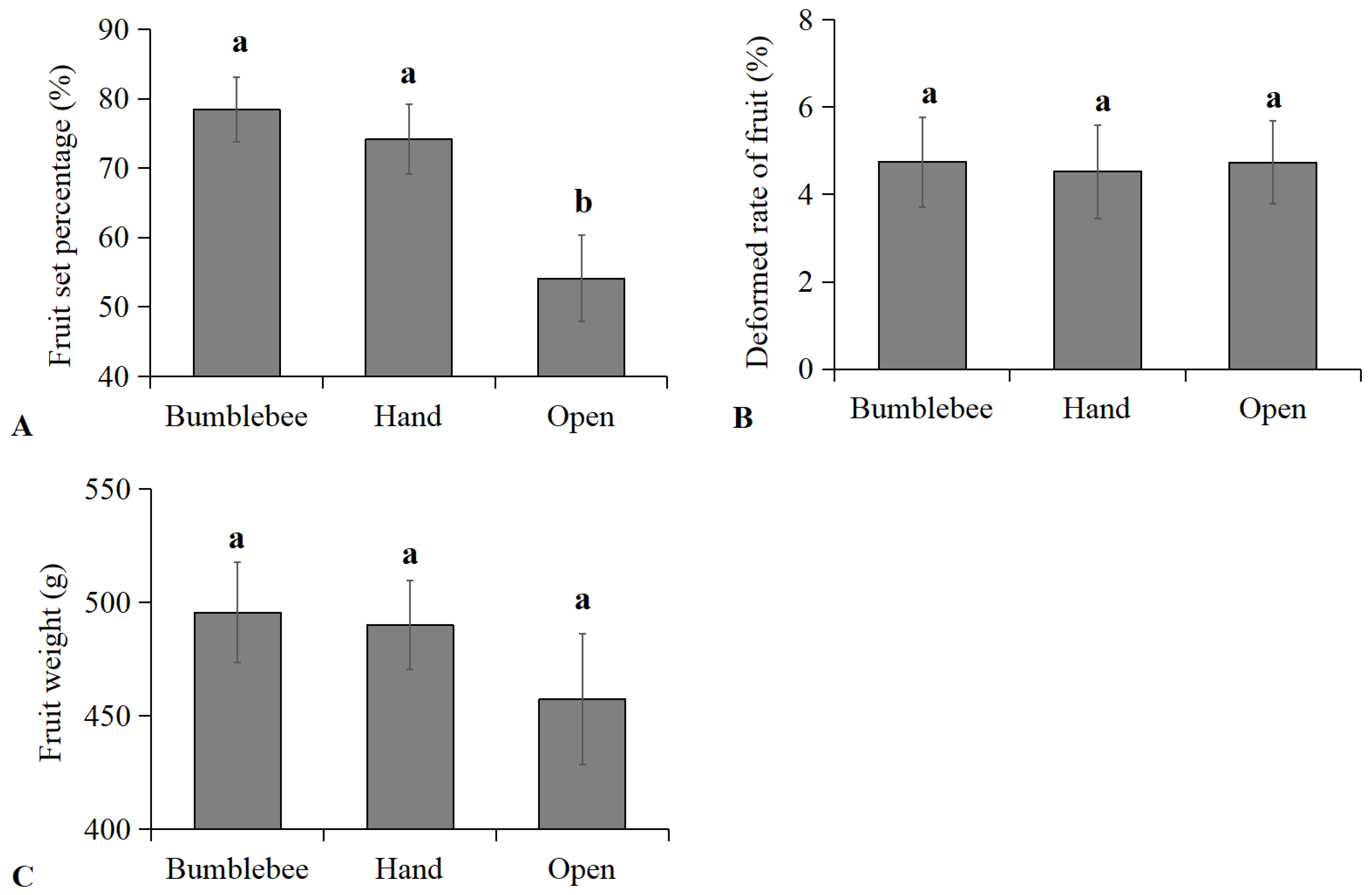
3.4. Effects of Bumblebee Pollination on Cherry Tomato Fruit
3.5. Effects of Bumblebee Pollination on Pear Fruit
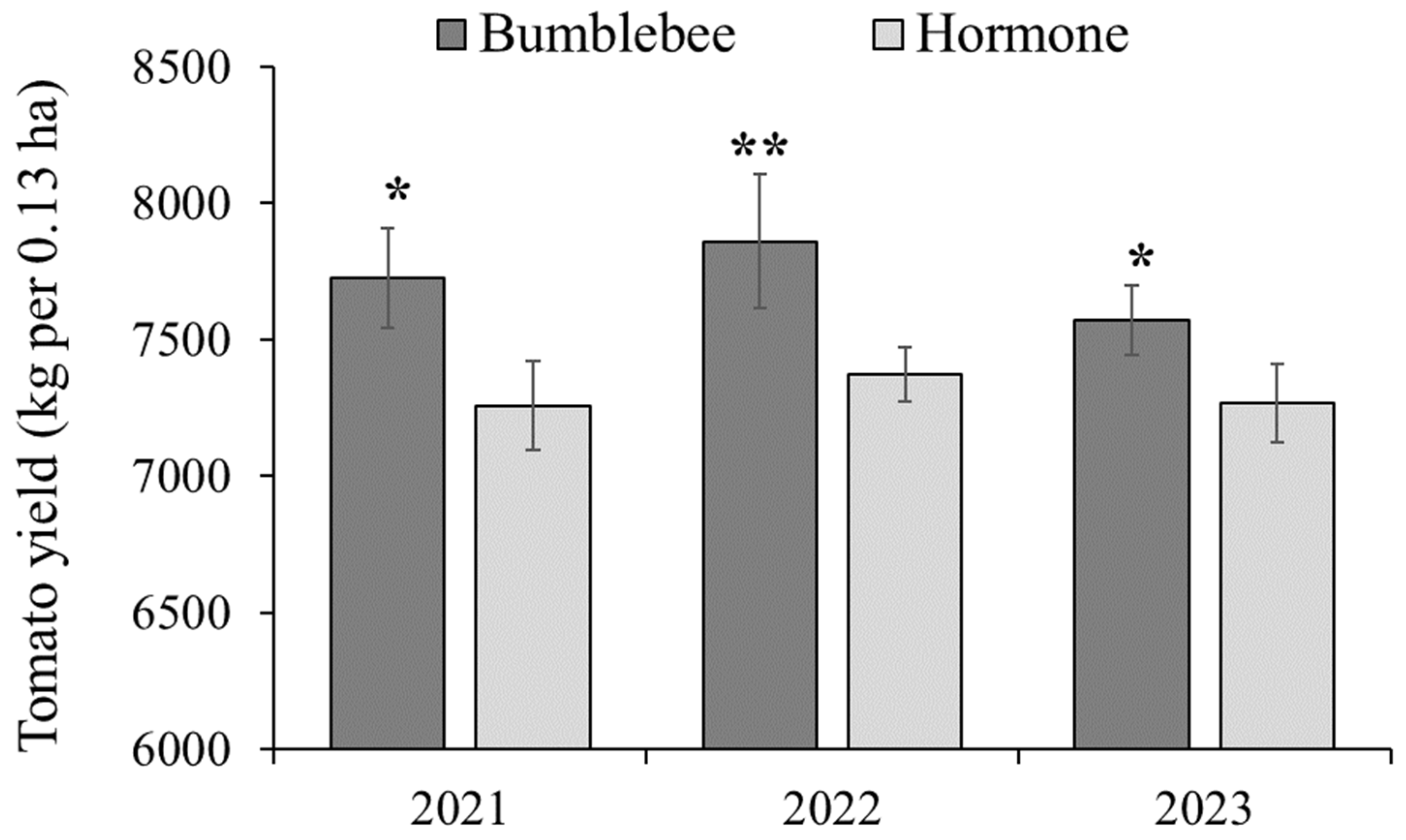
3.6. Yield Effects of Bumblebee Pollination on Cherry Tomatoes
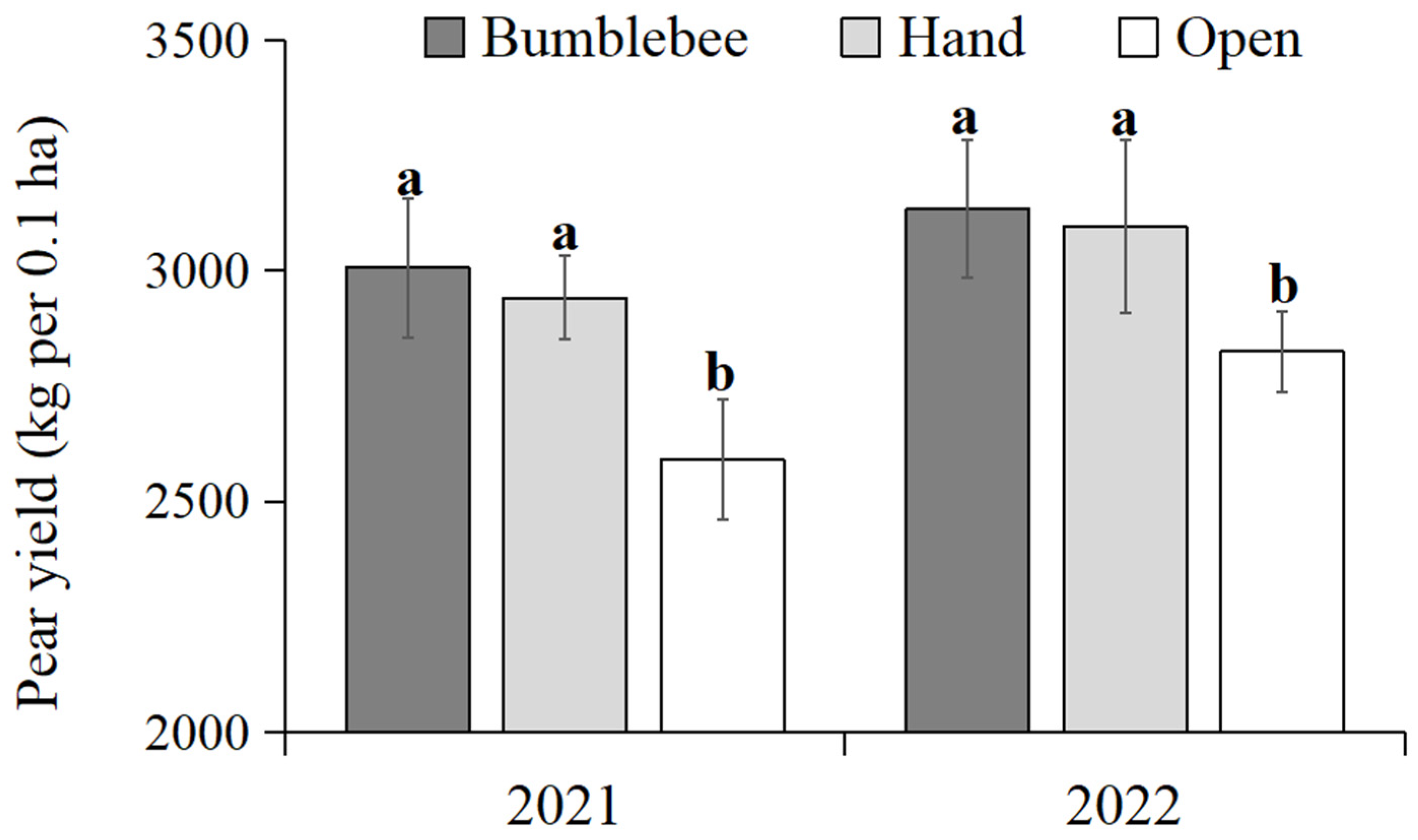
3.7. Yield Effects of Bumblebee Pollination on Pears
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cameron, S.A.; Sadd, B.M. Global trends in bumble bee health. Annu. Rev. Entomol. 2020, 65, 209–232. [Google Scholar] [CrossRef] [PubMed]
- Vinod, M.; Naik, A.K.; Biradar, R.; Natikar, P.K.; Balikai, R.A. Bee pollination under organic and conventional farming systems: A review. J. Exp. Zool. India 2016, 19, 643–652. [Google Scholar]
- Sladen, F.W.L. The Humblebee; MacMillan: London, UK, 1912. [Google Scholar]
- Kriesell, L.; Hilpert, A.; Leonhardt, S.D. Different but the same: Bumblebee species collect pollen of different plant sources but similar amino acid profiles. Apidologie 2017, 48, 102–116. [Google Scholar] [CrossRef]
- Stabler, D.; Paoli, P.P.; Nicolson, S.W.; Wright, G.A. Nutrient balancing of the adult worker bumblebee (Bombus terrestris) depends on the dietary source of essential amino acids. J. Exp. Biol. 2015, 218, 793–802. [Google Scholar] [CrossRef] [PubMed]
- Ghosha, S.; Jung, C. Changes in nutritional composition from bee pollen to pollen patty used in bumblebee rearing. J. Asia-Pac. Entomol. 2020, 23, 701–708. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z.Y.; An, J.D. Pollen release dynamics and daily patterns of pollen-collecting activity of honeybee Apis mellifera and bumblebee Bombus lantschouensis in solar greenhouse. Insects 2019, 10, 216. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Huang, J.; Wang, Y.; Zhao, X.; Su, L.; Thomas, G.W.C.; Zhao, M.; Zhang, X.; Jungreis, I.; Kellis, M.; et al. Genus-wide characterization of bumblebee genomes provides insights into their evolution and variation in ecological and behavioral traits. Mol. Biol. Evol. 2020, 38, 486–501. [Google Scholar] [CrossRef]
- Pashalidou, F.G.; Lambert, H.; Peybernes, T.; Mescher, M.C.; De Moraes, C.M. Bumble bees damage plant leaves and accelerate flower production when pollen is scarce. Science 2020, 368, 881–884. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; Van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Huang, J.; An, J.; Wu, J. Advantage of bumblebee as pollinator for solanum in greenhouse. Chin. Agric. Sci. Bul. 2007, 23, 5–9. [Google Scholar]
- Liu, X.Y.; Gao, C.D. Artificial Breeding and Pollination of Bumblebee; Northwest A & F University Press: Xianyang, China, 2011. [Google Scholar]
- Wu, G.A.; Yin, Y.Y.; Chen, H.; Zhou, H.; Yu, Y.; Zheng, L.; Zhai, Y.F. Comparative study on pollination by Bombus terrestris and Apis mellifera for blueberry in greenhouse. Apic. China 2019, 70, 68–70. [Google Scholar]
- Huang, X.B.; Li, H.; Dai, X.Y.; Wu, G.A.; Zhou, H.; Chen, H.; Zheng, L.; Zhai, Y.F. Research progress of bumblebee behavior and pollination application. Shandong Agric. Sci. 2021, 53, 130–137. [Google Scholar]
- Li, L.; Li, M.; Ni, X.; Wei, M. Effects of different pollination methods on fruit setting and seed yield of tomato. J. Anhui Agric. Sci. 2022, 50, 42–44. [Google Scholar]
- Dyer, A.G. Bumblebee search time without ultraviolet light. J. Exp. Biol. 2004, 207, 1683–1688. [Google Scholar] [CrossRef]
- Yin, Y.; Zhai, Y.; Sun, M.; Wu, G.; Zheng, L.; Chen, H. Application efficiency of insect natural enemies combined with Bombus terrestris pollination on cucumber in greenbouse. North Hortic. 2018, 17, 64–68. [Google Scholar]
- Whitney, H.M.; Dyer, A.; Chittka, L.; Rands, S.A.; Glover, B.J. The interaction of temperature and sucrose concentration on foraging preferences in bumblebees. Naturwissenschaften 2008, 95, 845–850. [Google Scholar] [CrossRef]
- Worden, B.D.; Papaj, D.R. Flower choice copying in bumblebees. Biol. Lett. 2005, 4, 504–507. [Google Scholar] [CrossRef]
- Alem, S.; Perry, C.J.; Zhu, X.; Loukola, O.J.; Ingraham, T.; Søvik, E.; Chittka, L. Associative mechanisms allow for social learning and cultural transmission of string pulling in an insect. PLoS Biol. 2016, 14, e1002564. [Google Scholar] [CrossRef]
- Röseler, P.F. A technique for year-round rearing of Bombus terrestris (Apidae, Bombini) colonies in captivity. Apidologie 1985, 16, 165–170. [Google Scholar] [CrossRef]
- Wang, G. Analysis of planting benefit of facility tomato industry in China. North Hortic. 2021, 16, 155–161. [Google Scholar]
- Sun, Y.; Xu, G.; Gao, W.; Li, D.; Shi, L.; Han, B. Effects of different pollination methods on yield, quality and benefit of tomato in solar greenhouse. China Veg. 2017, 6, 38–41. [Google Scholar]
- Huang, X.; Li, H.; Chen, G.; Dai, X.; Zhai, Y.; Zheng, L.; Chen, H.; Zhu, P.; Ding, J. Pollination behavior of Bombus terristris in winter greenhouse for tomato production in northern China. Shandong Agric. Sci. 2022, 54, 111–116. [Google Scholar]
- Bukowska, B. Toxicity of 2,4-Dichlorophenoxyacetic acid-molecular mechanisms. Pol. J. Environ. Stud. 2006, 15, 365–374. [Google Scholar]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2018, 158, 80–88. [Google Scholar] [CrossRef]
- Dyer, A.G.; Whitney, H.M.; Arnold, S.E.J.; Glover, B.J.; Glover, L. Behavioural ecology: Bees associate warmth with floral colour. Nature 2006, 442, 525. [Google Scholar] [CrossRef] [PubMed]
- Ayasse, M.; Jarau, S. Chemical ecology of bumble bees. Annu. Rev. Entomol. 2014, 59, 299–319. [Google Scholar] [CrossRef]
- Brown, M.; Brown, M.J.F. Nectar preferences in male bumblebees. Insectes Sociaux 2020, 67, 221–228. [Google Scholar] [CrossRef]
- Goulson, D. Bumblebees: Behavior, Ecology, and Conservation; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Ruedenauer, F.A.; Spaethe, J.; Leonhardt, S.D. How to know which food is good for you: Bumblebees use taste to discriminate between different concentrations of food differing in nutrient content. J. Exp. Biol. 2015, 218, 2233–2240. [Google Scholar] [CrossRef]
- Morgan, T.; Whitehorn, P.; Lye, G.C.; Vallejo-Marín, M. Floral sonication is an innate behaviour in bumblebees that can be fine-tuned with experience in manipulating flowers. J. Insect. Behav. 2016, 29, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. Food Exploitation by Social Insects: Ecological, Behavioral, and Theoretical Approaches; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Witjes, S.; Eltz, T. Influence of scent deposits on flower choice: Experiments in an artificial flower array with bumblebees. Apidologie 2007, 38, 12–18. [Google Scholar] [CrossRef]
- Baracchi, D.; Brown, M.J.F.; Chittka, L. Behavioural evidence for self-medication in bumblebees? F1000 Res. 2015, 4, 73. [Google Scholar] [CrossRef]
- Tanaka, C.S.; Ikemoto, M.; Nikkeshi, A.; Kanbe, Y.; Mitsuhata, M.; Yokoi, T. Ovarian development related to pollen feeding in workers of the bumblebee Bombus ignitus (Hymenoptera: Apidae). Appl. Entomol. Zool. 2019, 54, 85–89. [Google Scholar] [CrossRef]
- Goulson, D.; Stout, J.C.; Langley, J.; Hughes, W.O.H. Identity and function of scent marks deposited by foraging bumblebees. J. Chem. Ecol. 2000, 26, 2897–2911. [Google Scholar] [CrossRef]
- Molet, M.; Chittka, L.; Raine, N.E. How floral odours are learned inside the bumblebee (Bombus terrestris) nest. Naturwissenschaften 2008, 96, 213–219. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Chen, D.; Wu, W. Effects of different pollination methods on yield and quality of greenhouse tomato. J. Changjiang Veg. 2020, 24, 56–58. [Google Scholar]
- Goulson, D.; Lye, C.G.; Darvill, B. Decline and conservation of bumblebees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Carvell, C.; Bourke, A.F.G.; Dreier, S.; Freeman, S.N.; Hulmes, S.; Jordan, W.C.; Heard, M.S. Bumblebee family lineage survival is enhanced in high-quality landscapes. Nature 2017, 543, 547–549. [Google Scholar] [CrossRef] [PubMed]
- Rundlöf, M.; Nilsson, H.; Smith, H.G. Interacting effects of farming practice and landscape context on bumble bees. Biol. Conserv. 2008, 141, 417–426. [Google Scholar] [CrossRef]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Mass flowering crops enhance pollinator densities at a landscape scale. Ecol. Lett. 2010, 6, 961–965. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Blacquiere, T.; Field, L.M.; Hails, R.S.; Petrokofsky, G.; Potts, S.G.; Raine, N.E.; Vanbergen, A.J.; McLean, A.R. A restatement of the natural science evidence base concerning neonicotinoid insecticides and insect pollinators. Proc. R. Soc. B 2014, 281, 20140558. [Google Scholar] [CrossRef]
- Smith, D.B.; Arce, A.N.; Rodrigues, A.R.; Bischoff, P.H.; Gill, R.J. Insecticide exposure during brood or early-adult development reduces brain growth and impairs adult learning in bumblebees. Proc. R. Soc. B 2020, 287, 20192442. [Google Scholar] [CrossRef]
- Stanley, D.A.; Russell, A.L.; Morrison, S.J.; Rogers, C.; Raine, N.E. Investigating the impacts of field-realistic exposure to a neonicotinoid pesticide on bumblebee foraging, homing ability and colony growth. J. Appl. Ecol. 2016, 53, 1440–1449. [Google Scholar] [CrossRef]
- Muth, F.; Leonard, A.S. A neonicotinoid pesticide impairs foraging, but not learning, in free-flying bumblebees. Sci. Rep. 2019, 9, 4764. [Google Scholar] [CrossRef]
- Bryden, J.; Gill, R.J.; Mitton, R.A.A.; Raine, N.E.; Jansen, V.A.A.; Hodgson, D. Chronic sublethal stress causes bee colony failure. Ecol. Lett. 2013, 16, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Zhou, T.; Sun, M.; Feng, M.; Guo, W.; Nie, L.; Song, Y.; Men, X.; Li, L.; Yu, Y. Exploratory comparison of flower visiting behavior and pollination ability of mason bees, bumblebees, and honey bees. J. Econ. Entomol. 2023, 116, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Orr, M.C.; Ren, Z.X.; Ge, J.; Tian, L.; An, J.; Huang, J.; Zhu, C.D.; Williams, P.H. The rising threat of the invasive bumblebee Bombus terrestris highlights the need for sales restrictions and domestication of unique local biodiversity in Asia. Entomol. Gen. 2022, 42, 655–658. [Google Scholar] [CrossRef]
- Rendoll-Carcamo, J.A.; Contador, T.A.; Saavedra, L.; Montalva, J. First record of the invasive bumblebee Bombus terrestris (Hymenoptera: Apidae) on Navarino Island, southern Chile (55 S). J. Melittol. 2017, 71, 1–5. [Google Scholar] [CrossRef]
- Otterstatter, M.C.; Thomson, J.D. Does pathogen spillover from commercially reared bumble bees threaten wild pollinators? PLoS ONE 2008, 3, e2771. [Google Scholar] [CrossRef] [PubMed]
- Fürst, M.A.; McMahon, D.P.; Osborne, J.L.; Paxton, R.J.; Brown, M.J.F. Disease associations between honeybees and bumblebees as a threat to wild pollinators. Nature 2014, 506, 364–366. [Google Scholar] [CrossRef]
- Arismendi, N.; Riveros, G.; Zapata, N.; Smagghe, G.; Gonzalez, C.; Vargas, M. Occurrence of bee viruses and pathogens associated with emerging infectious diseases in native and non-native bumble bees in southern Chile. Biol. Invasions 2021, 23, 1175–1189. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flowers | Number of Visited Flowers per min | Single-Flower Residence Time (s) |
|---|---|---|
| Cherry tomato | 8.6 ± 1.2 | 5.6 ± 0.7 |
| Pear | 6.4 ± 0.9 | 7.1 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Zhang, Q.; Sheikh, U.A.A.; Wang, Y.; Zheng, L. Bumblebee Foraging Dynamics and Pollination Outcomes for Cherry Tomato and Pear Varieties in Northern China. Insects 2024, 15, 216. https://doi.org/10.3390/insects15040216
Huang X, Zhang Q, Sheikh UAA, Wang Y, Zheng L. Bumblebee Foraging Dynamics and Pollination Outcomes for Cherry Tomato and Pear Varieties in Northern China. Insects. 2024; 15(4):216. https://doi.org/10.3390/insects15040216
Chicago/Turabian StyleHuang, Xunbing, Qianwen Zhang, Umer Ayyaz Aslam Sheikh, Yueyue Wang, and Li Zheng. 2024. "Bumblebee Foraging Dynamics and Pollination Outcomes for Cherry Tomato and Pear Varieties in Northern China" Insects 15, no. 4: 216. https://doi.org/10.3390/insects15040216
APA StyleHuang, X., Zhang, Q., Sheikh, U. A. A., Wang, Y., & Zheng, L. (2024). Bumblebee Foraging Dynamics and Pollination Outcomes for Cherry Tomato and Pear Varieties in Northern China. Insects, 15(4), 216. https://doi.org/10.3390/insects15040216