


Copulation Duration and Sperm Precedence with Reference to Larval Diapause Induction in Monochamus alternatus Hope (Coleoptera: Cerambycidae)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Pine Bolt, Branch and Twig Sections
2.3. Larval Inoculation
2.4. Identification of Diapause and Larval Instars
2.5. Relationship between the Duration of Copulation and Egg Fertilization (Experiment 1)
2.6. Second-Male Sperm Precedence (Experiments 2 and 3)
2.7. Estimation of P2 Values (Second-Male Sperm Precedence)
2.8. Statistical Analysis
3. Results
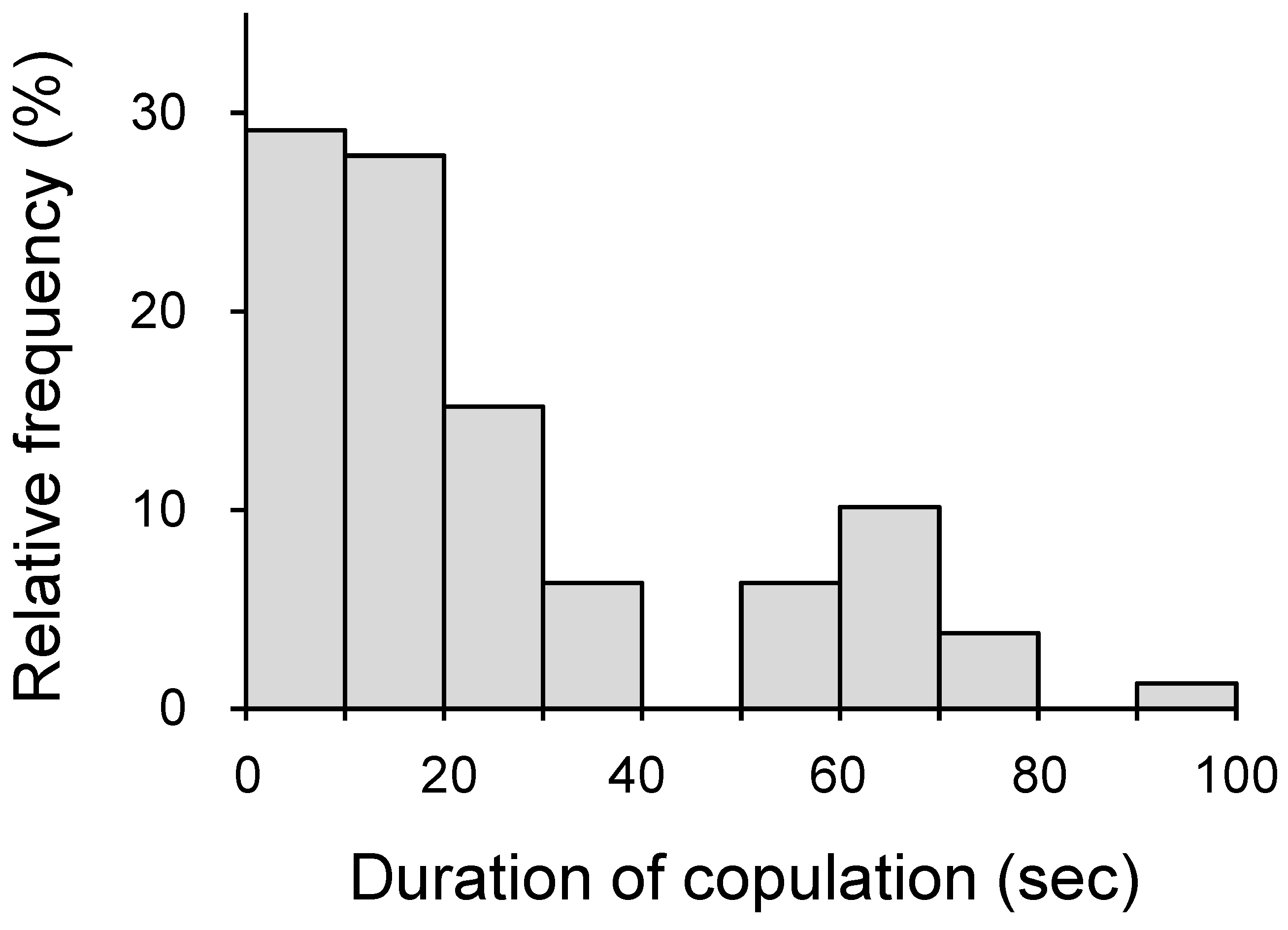
3.1. Relationship between Copulation Duration and Egg Fertilization (Experiment 1)
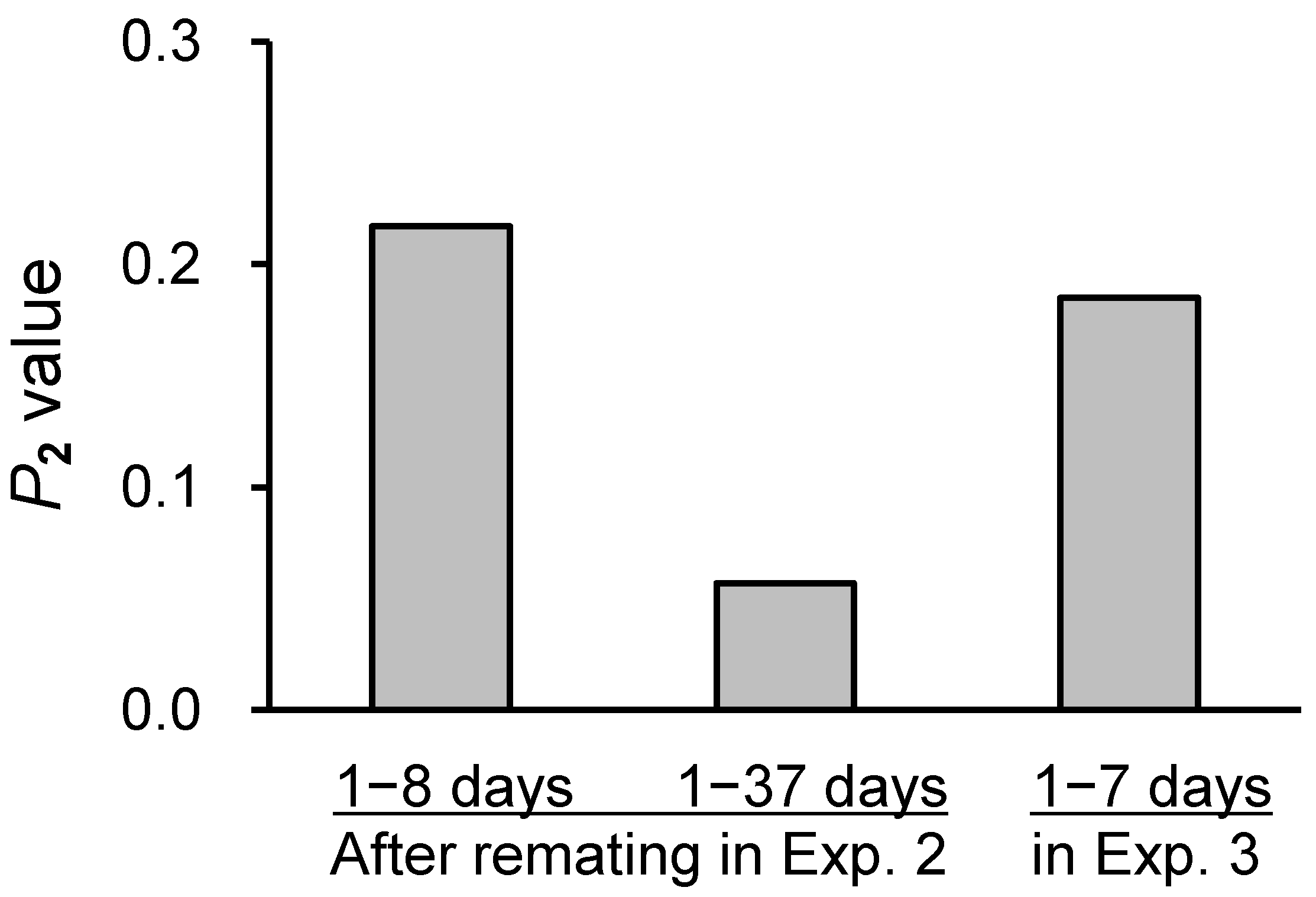
3.2. Second-Male Sperm Precedence (Experiments 2 and 3)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, M.L. Evolution Through Genetic Exchange; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Ellstrand, N.C.; Schierenbeck, K.A. Hybridization as a stimulus for the evolution of invasiveness in plants? Proc. Natl. Acad. Sci. USA 2000, 97, 7043–7050. [Google Scholar] [CrossRef] [PubMed]
- Hasselman, D.J.; Argo, E.E.; McBride, M.C.; Bentzen, P.; Schultz, T.F.; Perez-Umphrey, A.A.; Palkovacs, E.P. Human disturbance causes the formation of a hybrid swarm between two naturally sympatric fish species. Mol. Ecol. 2014, 23, 1137–1152. [Google Scholar] [CrossRef] [PubMed]
- Hovick, S.M.; Whitney, K.D. Hybridisation is associated with increased fecundity and size in invasive taxa: Meta-analytic support for the hybridisation-invasion hypothesis. Ecol. Lett. 2014, 17, 1464–1477. [Google Scholar] [CrossRef] [PubMed]
- Schierenbeck, K.A.; Ellstrand, N.C. Hybridization and the evolution of invasiveness in plants and other organisms. Biol. Invasions 2009, 11, 1093–1105. [Google Scholar] [CrossRef]
- Prentis, P.J.; Wilson, J.R.U.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. Adaptive evolution in invasive species. Trends Plant Sci. 2008, 13, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Rieseberg, L.H.; Raymond, O.; Rosenthal, D.M.; Lai, Z.; Livingstone, K.; Nakazato, T.; Durphy, J.L.; Schwarzbach, A.E.; Donovan, L.A.; Lexer, C. Major ecological transitions in wild sunflowers facilitated by hybridization. Science 2003, 301, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Culley, T.M.; Hardiman, N.A. The role of intraspecific hybridization in the evolution of invasiveness: A case study of the ornamental pear tree Pyrus calleryana. Biol. Invasions 2009, 11, 1107–1119. [Google Scholar] [CrossRef]
- Chapman, R.F. The Insects—Structure and Function; American Elsevier: New York, NY, USA, 1971. [Google Scholar]
- Fedina, T.Y.; Lewis, S.M. An integrative view of sexual selection in Tribolium flour beetles. Biol. Rev. 2008, 83, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R. Sexual selection in the black-tipped hangingfly. Sci. Am. 1980, 242, 162–172. [Google Scholar] [CrossRef]
- Filippi, L.; Nomakuchi, S.; Hironaka, M.; Tojo, S. Insemination success discrepancy between long-term and short-term copulations in the provisioning shield bug, Parastrachia japonensis (Hemiptera: Cydnidae). J. Ethol. 2000, 18, 29–36. [Google Scholar] [CrossRef]
- Kuriwada, T.; Kumano, N.; Shiromoto, K.; Haraguchi, D. Female walking during copulation reduces the likelihood of sperm transfer from males in the sweet potato weevil, Cylas formicarius. Entomol. Exp. Appl. 2013, 147, 25–230. [Google Scholar] [CrossRef]
- Parker, G.A.; Simmons, L.W. A model of constant random sperm displacement during mating: Evidence from Scatophaga. Proc. R. Sci. Lond. B 1991, 246, 107–115. [Google Scholar] [CrossRef]
- Yokoi, N. Observation on the mating behavior of the yellow spotted longicorn beetle, Psacothea hilaris Pascoe (Coleoptera: Cerambycidae). Jpn. J. Appl. Entomol. Zool. 1989, 33, 175–179. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Pizzari, T. Postcopulatory sexual selection. Nat. Rev. Genet. 2002, 3, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Connolly, S.R.; Hisano, M.; Dornelas, M.; Magurran, A.E. Fitness consequences of female multiple mating: A direct test of indirect benefits. BMC Evol. Biol. 2012, 12, 185. [Google Scholar] [CrossRef] [PubMed]
- Jennions, M.D.; Petrie, M. Why do females mate multiply? A review of the genetic benefits. Biol. Rev. 2000, 75, 21–64. [Google Scholar] [CrossRef] [PubMed]
- Kvarnemo, C.; Simmons, L.W. Polyandry as a mediator of sexual selection before and after mating. Phil. Trans. R. Soc. B 2013, 368, 20120042. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R.; Alcock, J. The Evolution of Insect Mating System; Harvard University Press: Cambridge, UK, 1983. [Google Scholar]
- Parker, G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–568. [Google Scholar] [CrossRef]
- Waage, J.K. Dual function of the damselfly penis: Sperm removal and transfer. Science 1979, 203, 916–918. [Google Scholar] [CrossRef]
- Yokoi, N. The sperm removal behavior of the yellow spotted longicorn beetle Psacothea hilaris (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1990, 25, 383–388. [Google Scholar] [CrossRef]
- Parker, G.A. Conceptual developments in sperm competition: A very brief synopsis. Phil. Trans. R. Soc. B 2020, 375, 20200061. [Google Scholar] [CrossRef] [PubMed]
- Córdoba-Aguilar, A.; Uhía, E.; Cordero Rivera, A. Sperm competition in Odonata (Insecta): The evolution of female sperm storage and rivals’ sperm displacement. J. Zool. Lond. 2003, 261, 381–398. [Google Scholar] [CrossRef]
- Pischedda, A.; Rice, W.R. Partitioning sexual selection into its mating success and fertilization success components. Proc. Natl. Acad. Sci. USA 2012, 109, 2049–2053. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, W.G. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Princeton, NJ, USA, 1996. [Google Scholar]
- Simmons, L.W. Sperm Competition and Its Evolutionary Consequences in the Insects; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Morimoto, J.; McDonald, G.C.; Wigby, S. Social group composition modulates the role of last male sperm precedence in post-copulatory sexual selection. J. Evol. Biol. 2023, 36, 1102–1115. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, S.; Togashi, K.; Linit, M.J. Cerambycids as Plant Disease Vectors with Special Reference to Pine Wilt. In Cerambycidae of the World—Biology and Pest Management; Wang, Q., Ed.; CRC Press: New York, NY, USA, 2017; pp. 209–252. [Google Scholar]
- Makihara, H. Two new species and a new subspecies of Japanese Cerambycidae (Coleoptera). Bull. For. For. Prod. Res. Inst. 2004, 3, 15–24. [Google Scholar]
- Lee, S.; Oh, D.-J.; Lee, S.; Chung, S.; Kim, D. Subspecific synonym of Monochamus alternatus (Coleoptera: Cerambycidae): Population genetics and morphological reassessment. J. Econ. Entomol. 2022, 115, 1987–1994. [Google Scholar] [CrossRef]
- Nishimura, M. Daily observation on behaviours of Japanese pine sawyer adult, Monochamus alternatus Hope. J. Jpn. For. Soc. 1973, 55, 100–104, (In Japanese with English Summary). [Google Scholar]
- Fauziah, B.A.; Hidaka, T.; Tabata, K. The reproductive behavior of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1987, 22, 272–285. [Google Scholar] [CrossRef]
- Anbutsu, H.; Togashi, K. Deterred oviposition response of Monochamus alternatus (Coleoptera: Cerambycidae) to oviposition scars occupied by eggs. Agric. For. Entomol. 2000, 2, 217–223. [Google Scholar] [CrossRef]
- Togashi, K. Development of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in relation to oviposition time. Jpn. J. Appl. Entomol. Zool. 1989, 33, 1–8, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Togashi, K. Life table for Monochamus alternatus (Coleoptera: Cerambycidae) within dead trees of Pinus thunbergii. Jpn. J. Entomol. 1990, 58, 217–230. [Google Scholar] [CrossRef]
- Enda, N.; Kitajima, H. Rearing of adults and larvae of the Taiwanese pine sawyer (Monochamus alternatus Hope, Coleoptera, Cerambycidae) on artificial diets. Trans. Annu. Meet. Jpn. For. Soc. 1990, 101, 503–504. (In Japanese) [Google Scholar]
- Togashi, K. Effects of photoperiod and chilling on diapause induction, intensity and termination in Monochamus alternatus endai (Coleoptera: Cerambycidae). J. For. Res. 2019, 24, 243–249. [Google Scholar] [CrossRef]
- Enda, N.; Makihara, H. Biology of the genus Monochamus, especially Japanese pine sawyer M. alternatus (Coleoptera, Cerambycidae) (2). For. Pests 2006, 55, 11–21. (In Japanese) [Google Scholar]
- Togashi, K. Effects of larval food shortage on diapause induction and adult traits in Taiwanese Monochamus alternatus alternatus. Entomol. Exp. Appl. 2014, 151, 34–42. [Google Scholar] [CrossRef]
- Togashi, K. Effects of crowding on larval diapause and adult body size in Monochamus alternatus alternatus (Coleoptera: Cerambycidae). Can. Entomol. 2017, 149, 159–173. [Google Scholar] [CrossRef]
- Togashi, K.; Toki, W. Effects of inter-subspecies hybridization between non-native Monochamus alternatus alternatus and native M. a. endai (Coleoptera: Cerambycidae) on the induction of larval diapause and adult body size. Appl. Entomol. Zool. 2018, 53, 29–40. [Google Scholar] [CrossRef]
- Togashi, K. Different developments of overwintered larvae of Monochamus alternatus (Coleoptera: Cerambycidae) under a constant temperature. Jpn. J. Entomol. 1991, 59, 149–154. [Google Scholar]
- Kozima, K.; Katagiri, K. On the larval instar and changes of its composition of Monochamus alternatus Hope. J. Jpn. For. Soc. 1964, 46, 307–310, (In Japanese with English Summary). [Google Scholar]
- Morimoto, K.; Iwasaki, A. Studies on the pine sawyer. X. Number of larval instar. Trans. Annu. Meet. Jpn. For. Soc. 1974, 85, 503–504. (In Japanese) [Google Scholar]
- Ochi, K. Ecological studies on the Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae) (II). Size of the first instar larva. Trans. Annu. Meet. Jpn. For. Soc. 1975, 86, 323–324. (In Japanese) [Google Scholar]
- Togashi, K. Effects of maternal body mass, age and crowding on offspring diapause induction and life-history traits in Monochamus alternatus alternatus (Coleoptera; Cerambycidae). J. For. Res. 2023; submitted. [Google Scholar]
- Freeman, J.B.; Dale, R. Assessing bimodality to detect the presence of a dual cognitive process. Behav. Res. 2013, 45, 83–97. [Google Scholar] [CrossRef]
- Pfister, R.; Schwarz, K.A.; Janczyk, M.; Dale, R.; Freeman, J.B. Good things peak in pairs: A note on the bimodality coefficient. Front. Psychol. 2013, 4, 700. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L. Reproductive behavior and sexual dimorphism in the white-spotted sawyer Monochamus scutellatus (Say). Coleopt. Bull. 1979, 33, 45–47. [Google Scholar]
- Kobayashi, H.; Yamane, A.; Iwata, R. Mating behavior of the pine sawyer, Monochamus saltuarius (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2003, 38, 141–148. [Google Scholar] [CrossRef]
- Yamamura, N. An evolutionarily stable strategy (ESS) model of postcopulatory guarding in insects. Theor. Popul. Biol. 1986, 29, 438–455. [Google Scholar] [CrossRef]
- Harts, A.M.F.; Kokko, H. Understanding promiscuity: When is seeking additional mates better than guarding an already found one? Evolution 2013, 67, 2838–2848. [Google Scholar] [CrossRef] [PubMed]
- McLain, D.K. Female choice and the adaptive significance of prolonged copulation in Nezara viridula (Hemiptera: Pentatomaidae). Psyche 1980, 87, 325–336. [Google Scholar] [CrossRef]
- Rutowski, R.L.; Alcock, J. Temporal variation in male copulatory behavior in the solitary bee Nomadopsis puellae (Hymenoptera: Andrenidae). Behaviour 1980, 73, 175–188. [Google Scholar] [CrossRef]
- Parker, G.A.; Simmons, L.W. Evolution of phenotypic optima and copula duration in dungflies. Nature 1994, 370, 53–56. [Google Scholar] [CrossRef]
- Kelly, C.D. Male-biased sex ratios and plasticity in post-insemination behaviour in the New Zealand stick insect Micrarchus hystriculeus. Behaviour 2015, 152, 653–666. [Google Scholar] [CrossRef]
- Hughes, A.L.; Hughes, M.K. Male size, mating success, and breeding habitat partitioning in the whitespotted sawyer Monochamus scutellatus (Say) (Coleoptera: Cerambycidae). Oecologia 1982, 55, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Linit, M.J. Comparison of oviposition and longevity of Monochamus alternatus and M. carolinensis (Coleoptera: Cerambycidae) under laboratory conditions. Environ. Entomol. 1998, 27, 855–891. [Google Scholar] [CrossRef]
- Arnqvist, G.; Nilsson, T. The evolution of polyandry: Multiple mating and female fitness in insects. Anim. Behav. 2000, 60, 145–160. [Google Scholar] [CrossRef]
- Katsuyama, N.; Sakurai, H.; Tabata, K.; Takeda, S. Effect of age of post-feeding twig on the ovarian development of Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Res. Bull. Fac. Agric. Gifu. Univ. 1989, 54, 81–89. (In Japanese) [Google Scholar]
- Ikeda, T.; Enda, N.; Yamane, A.; Oda, K.; Toyoda, T. Attractants for the Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1980, 15, 358–361. [Google Scholar] [CrossRef]
- Ikeda, T.; Oda, K. The occurrence of attractiveness for Monochamus alternatus Hope (Coleoptera: Cerambycidae) in nematode-infested pine trees. J. Jpn. For. Soc. 1980, 62, 432–434. [Google Scholar]
- Ikeda, T.; Yamane, A.; Enda, N.; Oda, K.; Makihara, H.; Ito, K.; Ôkochi, I. Attractiveness of volatile components of felled pine trees for Monochamus alternatus Hope (Coleoptera: Cerambycidae). J. Jpn. For. Soc. 1986, 68, 15–19, (In Japanese with English Summary). [Google Scholar]
- Shibata, E. Dispersal movement of the adult Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae) in a young pine forest. Appl. Entomol. Zool. 1986, 21, 184–186. [Google Scholar] [CrossRef]
- Togashi, K. Studies on population dynamics of Monochamus alternatus Hope (Coleoptera: Cerambycidae) and spread of pine wilt disease caused by Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae). Bull. Ishikawa For. Exp. Sta. 1989, 20, 1–142, (In Japanese with English Summary). [Google Scholar]
- Allison, J.D.; Borden, J.H.; McIntosh, R.L.; de Groot, P.; Gries, R. Kairomonal response by four Monochamus species (Coleoptera: Cerambycidae) to bark beetle pheromones. J. Chem. Ecol. 2001, 27, 633–646. [Google Scholar] [CrossRef]
- Fierke, M.K.; Skabeikis, D.D.; Millar, J.G.; Teale, S.A.; McElfresh, J.S.; Hanks, L.M. Identification of a male-produced aggregation pheromone for Monochamus scutellatus scutellatus and an attractant for the congener Monochamus notatus (Coleoptera: Cerambycidae). J. Econ. Entomol. 2012, 105, 2029–2034. [Google Scholar] [CrossRef]
- Lee, H.-R.; Lee, S.-C.; Lee, D.H.; Choi, W.-S.; Jung, C.-S.; Jeon, J.-H.; Kim, J.-E.; Park, I.-K. Identification of the aggregation-sex pheromone produced by male Monochamus saltuarius, a major insect vector of the pine wood nematode. J. Chem. Ecol. 2017, 43, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-R.; Lee, S.-C.; Lee, D.H.; Jung, M.; Kwon, J.-H.; Huh, M.-J.; Kim, D.-S.; Lee, J.-E.; Park, I.-K. Identification of aggregation-sex pheromone of the Korean Monochamus alternatus (Coleoptera: Cerambycidae) population, the main vector of pine wood nematode. J. Econ. Entomol. 2018, 111, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Pajares, J.A.; Álvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef]
- Teale, S.A.; Wickham, J.D.; Zhang, F.; Su, J.; Chen, Y.; Xiao, W.; Hanks, L.M.; Millar, J.G. A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J. Econ. Entomol. 2011, 104, 1592–1598. [Google Scholar] [CrossRef]
- Parker, G.A. Sexual conflict over mating and fertilization: An overview. Phil. Trans. R. Soc. B 2006, 361, 235–259. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Number of Copulations Allowed | Number of Trials | Female | Male | ||
|---|---|---|---|---|---|
| Body Mass (mg) 1 | Elytron Length (mm) 1 | Body Mass (mg) 1 | Elytron Length (mm) 1 | ||
| 1 | 9 | 394 ± 29 | 16.1 ± 0.5 | 348 ± 33 | 14.7 ± 0.5 |
| 10 | 8 | 359 ± 31 | 16.0 ± 0.5 | 347 ± 30 | 14.7 ± 0.3 |
| >10 (3 h) | 1 | 342 | 16.3 | 281 | 12.9 |
| Type of Mating (Female × Male) | Characteristics | Experiment 2 | Experiment 3 |
|---|---|---|---|
| T females that mated with a T male (mating T × T) | Number of trials | 4 | 10 |
| Age of females when paired (days) | 37 or 38 | 17–38 | |
| Age of males when paired (days) | 20–43 | 13–47 | |
| Number of larvae inoculated | 40 | 31 | |
| Time period from the start of pairing to egg collection (days) (Time period A) | 1–14 | – | |
| Time period from the start of pairing to larval inoculation (days) (Time period B) | 8–21 | 8–30 | |
| T females that remated with a J male after first mating with a T male (mating T × J after mating T × T) | Number of trials | 4 | 10 |
| Age of females when paired (days) | 45–51 | 34–56 | |
| Age of males when paired (days) | 13–23 | 79–97 | |
| Number of larvae inoculated immediately after being paired | 40 | 31 | |
| Time period from the end of pairing to egg collection (days) (Time period C) | 1–8 | 1–7 | |
| Time period from the end of pairing to larval inoculation (days) (Time period D) | 7–15 | 7–13 | |
| Number of larvae inoculated 29–43 days after being paired 1 | 37 | – | |
| Time period C | 20–37 | – | |
| Time period D | 29–43 | – | |
| T females that mated with a J male (mating T × J) 2 | Number of trials | 4 | 11 |
| Age of females when paired (days) | 33–44 | 8–32 | |
| Age of males when paired (days) | 4–17 | 65–92 | |
| Number of larvae inoculated | 36 | 31 | |
| Time period A | 1–33 | – | |
| Time period B | 8–53 | 7–45 | |
| J females that mated with a J male (mating J × J) 3 | Number of trials | 4 | 10 |
| Age of females when paired (days) | 22–26 | 61–75 | |
| Age of males when paired (days) | 15–22 | 61–73 | |
| Number of larvae inoculated | 36 | 31 | |
| Time period A | 2–20 | – | |
| Time period B | 8–27 | 7–19 |
| Pair of Adults (Female Code × Male Code) | Duration of Copulation (s) | Female Age When the First Egg Was Laid (Days) | Time from Copulation to First Egg Laying (Days) | Period of Egg Collection after 1st Pairing (Days) | Number of Eggs Laid | % of Eggs Fertilized after 1st Pairing 1 | % of Eggs Fertilized after 2nd Pairing 2 |
|---|---|---|---|---|---|---|---|
| F1 × M3 | 12.10 | 15 | 1 | 10 | 9 | 0.0 (0/9) | 100.0 (8/8) |
| F6 × M9 | 5.72 | 15 | 1 | 10 | 11 | 0.0 (0/11) | 100.0 (27/27) |
| F7 × M12 | 5.36 | 15 | 1 | 10 | 11 | 0.0 (0/11) | 66.7 (4/6) |
| F9 × M15 | 6.38 | 17 | 3 | 12 | 8 | 0.0 (0/8) | 90.0 (18/20) |
| F13 × M12 | 1.99 | 23 | 9 | 18 | 8 | 0.0 (0/8) | 94.7 (18/19) |
| F15 × M23 | 6.93 | 15 | 1 | 10 | 6 | 0.0 (0/6) | 28.6 (2/7) |
| F18 × M21 | 5.38 | 25 | 11 | 21 | 8 | 0.0 (0/7) | 100.0 (3/3) |
| F8 × M10 | 63.96 | 15 | 1 | 87 3 | 106 | 97.1 (100/103) | ― 4 |
| F16 × M24 | 108.89 | 33 | 19 | 38 3 | 12 | 100.0 (10/10) | ― 4 |
| Pair of Adults (Female Code × Male Code) | Duration of First 10 Copulations (s) 1 | Number of Long-Term Copula-tions 2 | Female Age when the First Egg Was Laid (Days) | Time from Copulation to First Egg Laying (Days) | Period of Egg Collection after 1st Pairing (Days) | Number of Eggs Laid | % of Eggs Fertilized after 1st Pairing 3 | % of Eggs Fertilized after 2nd Pairing 4 |
|---|---|---|---|---|---|---|---|---|
| F2 × M4 | 18.54 ± 3.60 (5.66–35.35) | 0 | 15 | 1 | 10 | 6 | 0.0 (0/6) | 100.0 (9/9) |
| F4 × M3 | 13.55 ± 2.11 (2.91–26.85) | 0 | 22 | 8 | 17 | 12 | 0.0 (0/11) | 96.2 (25/26) |
| F5 × M4 | 7.28 ± 0.97 (1.70–11.10) | 0 | 20 | 6 | 15 | 4 | 0.0 (0/4) | ― |
| F19 × M17 | 19.22 ± 2.90 (7.32–36.93) | 0 | 20 | 6 | 17 | 10 | 0.0 (0/10) | 100.0 (4/4) |
| F10 × M20 | 37.76 ± 8.16 (5.88–73.97) | 3 | 15 | 1 | 31 5 | 28 | 88.0 (22/28) | ― 6 |
| F12 × M9 | 49.41 ± 9.56 (6.46–90.32) | 3 | 15 | 1 | 65 5 | 111 | 86.1 (93/108) | ― 6 |
| F14 × M22 | 29.48 ± 6.81 (3.70–73.80) | 1 | 17 | 3 | 83 5 | 110 | 95.0 (96/101) | ― 6 |
| F17 × M22 | 33.37 ± 8.10 (2.29–69.57) | 1 | 24 | 10 | 72 5 | 98 | 96.9 (95/98) | ― 6 |
| F3 × M5 | Paired for 3 h | ― | 16 | 2 | 34 5 | 49 | 100.0 (46/46) | ― 6 |
| Female Characteristics | Number of Females Used | Number of Offspring | Incidence of Diapause (B/(A + B)) | P2 Value Estimated | |||||
|---|---|---|---|---|---|---|---|---|---|
| Forgoing Diapause (A) | Undergoing Diapause (B) | In Pre-Diapause State | Dead in Larval Stage | Dead without Feeding 1 | Total | ||||
| T females that mated with a T male | 4 | 23 | 4 | 1 | 7 | 5 | 40 | 0.148 b 2 | |
| T females 1–8 days after remating with a J male 3 | 4 | 20 (2) 4 | 10 | 0 | 7 | 3 | 40 | 0.333 b 2 | 0.217 |
| T females 20–37 days after remating with a J male 3 | 3 | 29 | 2 | 1 | 0 | 5 | 37 | 0.065 b 2 | |
| T females that mated with a J male | 4 | 0 | 33 | 1 | 1 | 1 | 36 | 1.000 a 2 | |
| J females that mated with a J male | 4 | 0 | 28 | 1 | 5 | 2 | 36 | 1.000 a 2 | |
| Female Characteristics | Number of Females Used | Number of Offspring | Incidence of Diapause (B/(A + B)) | P2 Value Estimated | ||||
|---|---|---|---|---|---|---|---|---|
| Forgoing Diapause (A) | Undergoing Diapause (B) | Dead in Larval Stage | Dead without Feeding 1 | Total | ||||
| T females that mated with a T male | 10 | 22 (1) 2 | 4 | 5 | 0 | 31 | 0.154 b 3 | |
| T females 1–7 days after remating with a J male 4 | 10 | 17 (1) 2 | 7 | 7 | 0 | 31 | 0.292 b 3 | 0.185 |
| T females that mated with a J male | 10 | 3 (1) 2 | 27 | 1 | 0 | 31 | 0.900 a 3 | |
| J females that mated with a J male | 10 | 0 | 28 | 2 | 1 | 31 | 1.000 a 3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Togashi, K.; Sugimoto, H. Copulation Duration and Sperm Precedence with Reference to Larval Diapause Induction in Monochamus alternatus Hope (Coleoptera: Cerambycidae). Insects 2024, 15, 255. https://doi.org/10.3390/insects15040255
Togashi K, Sugimoto H. Copulation Duration and Sperm Precedence with Reference to Larval Diapause Induction in Monochamus alternatus Hope (Coleoptera: Cerambycidae). Insects. 2024; 15(4):255. https://doi.org/10.3390/insects15040255
Chicago/Turabian StyleTogashi, Katsumi, and Hiroyuki Sugimoto. 2024. "Copulation Duration and Sperm Precedence with Reference to Larval Diapause Induction in Monochamus alternatus Hope (Coleoptera: Cerambycidae)" Insects 15, no. 4: 255. https://doi.org/10.3390/insects15040255
APA StyleTogashi, K., & Sugimoto, H. (2024). Copulation Duration and Sperm Precedence with Reference to Larval Diapause Induction in Monochamus alternatus Hope (Coleoptera: Cerambycidae). Insects, 15(4), 255. https://doi.org/10.3390/insects15040255