


Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.1.1. Testing the Effects of FVOCs on the Attraction of Subcortical Beetles
2.1.2. Testing the Combined Effects of FVOCs and MPB Pheromones on Subcortical Beetle Attraction
2.1.3. Testing the Effects of SVCs on the Attraction of Subcortical Beetles
2.2. Statistical Analysis
3. Results
3.1. The Effects of FVOCs, SVCs and MPB Lure on the Attraction of Subcortical Beetles
3.1.1. Catches of Subcortical Beetles in FVOC Treatments
3.1.2. Catches of Subcortical Insects in FVOCs with MPB Lure
3.1.3. Catches of Subcortical Insects in SVC Treatments
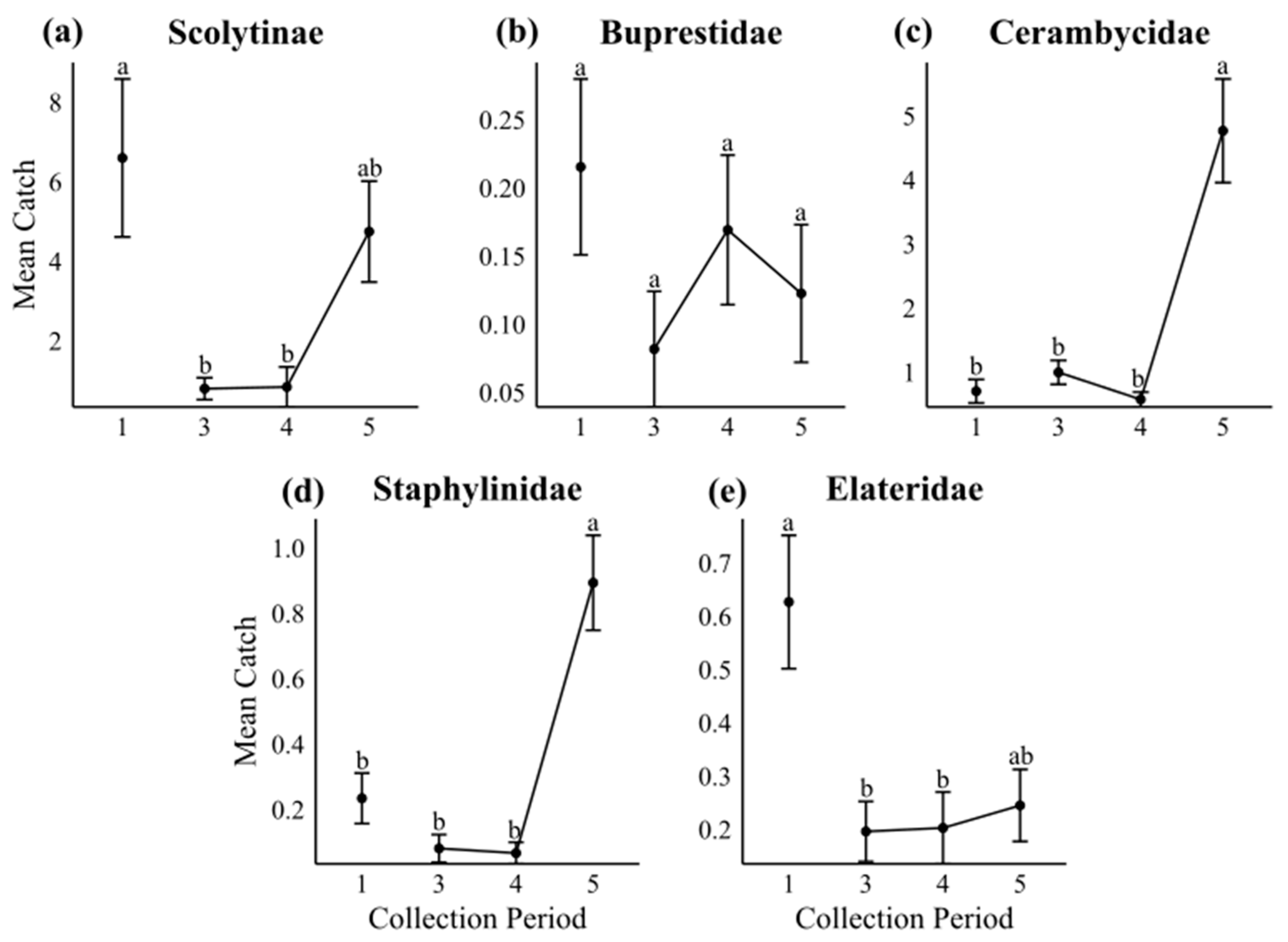
3.2. Temporal Variation in Subcortical Beetle Catches
3.3. Differences in Subcortical Beetle Catches Between Volatile Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Byers, J.A. Chemical ecology of bark beetles in a complex olfactory landscape. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.C., Evans, H.F., Eds.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar] [CrossRef]
- Erbilgin, N.; Christiansen, E.; Krokene, P. A host monoterpene influences Ips typographus responses (Coleoptera: Curculionidae, Scolytinae) to its aggregation pheromone: Implications for host colonization of bark beetles. Agric. For. Entomol. 2007, 9, 135–140. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Schlyter, F.; Byers, J.A.; Löfqvist, J. Attraction to pheromone sources of different quantity, quality, and spacing: Density-regulation mechanisms in bark beetle Ips typographus. J. Chem. Ecol. 1987, 13, 1503–1523. [Google Scholar] [CrossRef] [PubMed]
- Wallin, K.F.; Raffa, K.F. Influences of host chemicals and internal physiology on the multiple steps of postlanding host acceptance behavior of Ips pini (Coleoptera: Scolytidae). Environ. Entomol. 2000, 29, 442–453. [Google Scholar] [CrossRef]
- Seybold, S.J.; Bentz, B.J.; Fettig, C.J.; Lundquist, J.E.; Progar, R.A.; Gillette, N.E. Management of western North American bark beetles with semiochemicals. Annu. Rev. Entomol. 2018, 63, 407–432. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Gilmore, D.W.; Haack, R.A.; Katovich, S.A.; Krauth, S.J.; Mattson, W.J.; Zasada, J.C.; Seybold, S.J. Application of semiochemicals to assess the biodiversity of subcortical insects following an ecosystem disturbance in a sub-boreal forest. J. Chem. Ecol. 2009, 35, 1384–1410. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.; Nahrung, H.F.; Lawson, S.A.; Hayes, R.A. How Effective Are push–pull semiochemicals as deterrents for bark beetles? A global meta-analysis of thirty years of research. Insects 2023, 14, 812. [Google Scholar] [CrossRef]
- Singh, V.V.; Naseer, A.; Mogilicherla, K.; Trubin, A.; Zabihi, K.; Roy, A.; Jakuš, R.; Erbilgin, N. Understanding bark beetle outbreaks: Exploring the impact of changing temperature regimes, droughts, forest structure, and prospects for future forest pest management. Rev. Environ. Sci. Bio/Technol. 2024, 23, 257–290. [Google Scholar] [CrossRef]
- Erbilgin, N.; Powell, J.S.; Raffa, K.F. Effect of varying monoterpene concentrations on the response of Ips pini (Coleoptera: Scolytidae) to its aggregation pheromone: Implications for pest management and ecology of bark beetles. Agric. For. Entomol. 2003, 5, 269–274. [Google Scholar] [CrossRef]
- Borden, J.H. Semiochemicals and bark beetle populations: Exploitation of natural phenomena by pest management strategists. Ecography 1989, 12, 501–510. [Google Scholar] [CrossRef]
- Erbilgin, N.; Raffa, K.F. Kairomonal range of generalist predators in specialized habitats: Responses to multiple phloeophagous species emitting pheromones vs. host odors. Entomol. Exp. Appl. 2001, 99, 205–210. [Google Scholar] [CrossRef]
- Boone, C.K.; Six, D.L.; Raffa, K.F. The enemy of my enemy is still my enemy: Competitors add to predator load of a tree-killing bark beetle. Agric. For. Entomol. 2008, 10, 411–421. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kännaste, A.; Copolovici, L. Quantitative patterns between plant volatile emissions induced by biotic stresses and the degree of damage. Front. Plant Sci. 2013, 4, 262. [Google Scholar] [CrossRef] [PubMed]
- Faiola, C.; Taipale, D. Impact of insect herbivory on plant stress volatile emissions from trees: A synthesis of quantitative measurements and recommendations for future research. Atmos. Environ. X 2020, 5, 100060. [Google Scholar] [CrossRef]
- Zaman, R.; Antonioli, F.; Shah, A.; Ullah, A.; May, C.; Klutsch, J.G.; Erbilgin, N. A pine in distress: How infection by different pathogenic fungi affect lodgepole pine chemical defenses. Microb. Ecol. 2023, 86, 2666–2673. [Google Scholar] [CrossRef]
- Byers, J.A. Attraction of bark beetles, Tomicus piniperda, Hylurgops palliatus, and Trypodendron domesticum and other insects to short-chain alcohols and monoterpenes. J. Chem. Ecol. 1992, 18, 2385–2402. [Google Scholar] [CrossRef]
- Erbilgin, N.; Raffa, K.F. Opposing effects of host monoterpenes on responses by two sympatric species of bark beetles to their aggregation pheromones. J. Chem. Ecol. 2000, 26, 2527–2548. [Google Scholar] [CrossRef]
- Kandasamy, D.; Gershenzon, J.; Hammerbacher, A. Volatile organic compounds emitted by fungal associates of conifer bark beetles and their potential in bark beetle control. J. Chem. Ecol. 2016, 42, 952–969. [Google Scholar] [CrossRef]
- Kandasamy, D.; Gershenzon, J.; Andersson, M.N.; Hammerbacher, A. Volatile organic compounds influence the interaction of the Eurasian spruce bark beetle (Ips typographus) with its fungal symbionts. ISME J. 2019, 13, 1788–1800. [Google Scholar] [CrossRef]
- Jirošová, A.; Modlinger, R.; Hradecký, J.; Ramakrishnan, R.; Beránková, K.; Kandasamy, D. Ophiostomatoid fungi synergize attraction of the Eurasian spruce bark beetle, Ips typographus to its aggregation pheromone in field traps. Front. Microbiol. 2022, 13, 980251. [Google Scholar] [CrossRef]
- Zaman, R.; May, C.; Ullah, A.; Erbilgin, N. Bark beetles utilize ophiostomatoid fungi to circumvent host tree defenses. Metabolites 2023, 13, 239. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Asaro, C.; Crowe, C.M.; Duerr, D.A. Bark beetle pheromones and pine volatiles: Attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the southeastern United States. J. Econ. Entomol. 2011, 104, 1245–1257. [Google Scholar] [CrossRef]
- Grant, G.G.; Poland, T.M.; Ciaramitaro, T.; Lyons, D.B.; Jones, G.C. Comparison of male and female emerald ash borer (Coleoptera: Buprestidae) responses to phoebe oil and (Z)-3-Hexenol lures in light green prism traps. J. Econ. Entomol. 2011, 104, 173–179. [Google Scholar] [CrossRef]
- Domingue, M.J.; Baker, T.C.; Blanco, J.J.; Fernandes, A.T. A multi-disciplinary approach for developing tools to monitor invasive buprestid beetle species. In Invasive Species: Threats, Ecological Impact and Control; Nova Science Pub Inc: Hauppauge, NY, USA, 2012; Volume 3, pp. 77–100. [Google Scholar]
- Allison, J.D.; McKenney, J.L.; Miller, D.R.; Gimmel, M.L. Kairomonal responses of natural enemies and associates of the southern Ips (Coleoptera: Curculionidae: Scolytinae) to ipsdienol, ipsenol and cis-Verbenol. J. Insect Behav. 2012, 26, 321–335. [Google Scholar] [CrossRef]
- Miller, D.R. Coleopteran predators of bark and woodboring beetles attracted to traps baited with ethanol and α-pinene in pine (Pinaceae) forests of the southern United States of America. Can. Entomol. 2023, 155, e5. [Google Scholar] [CrossRef]
- Wood, D.L. The role of pheromones, kairomones, and allomones in the host selection and colonization behavior of bark beetles. Annu. Rev. Entomol. 1982, 27, 411–446. [Google Scholar] [CrossRef]
- Erbilgin, N.; Raffa, K.F. Modulation of predator attraction to pheromones of two prey species by stereochemistry of plant volatiles. Oecologia 2001, 127, 444–453. [Google Scholar] [CrossRef]
- Gitau, C.W.; Bashford, R.; Carnegie, A.J.; Gurr, G.M. A review of semiochemicals associated with bark beetle (Coleoptera: Curculionidae: Scolytinae) pests of coniferous trees: A focus on beetle interactions with other pests and their associates. For. Ecol. Manag. 2013, 297, 1–14. [Google Scholar] [CrossRef]
- Tóth, M. Pheromones and attractants of click beetles: An overview. J. Pest Sci. 2012, 86, 3–17. [Google Scholar] [CrossRef]
- Payne, T.L.; Coster, J.E.; Richerson, J.V.; Edson, L.J.; Hart, E.R. Field response of the southern pine beetle to behavioral chemicals. Environ. Entomol. 1978, 7, 578–582. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Cale, J.A.; Whitehouse, C.; Kanekar, S.S.; Erbilgin, N. Trap trees: An effective method for monitoring mountain pine beetle activities in novel habitats. Can. J. For. Res. 2017, 47, 1432–1437. [Google Scholar] [CrossRef]
- Borden, J.H.; Hunt, D.W.A.; Miller, D.R.; Slessor, K.N. Orientation in Forest Coleoptera: An Uncertain Outcome to Responses by Individual Beetles to Variable Stimuli; Clarendon Press: Oxford, UK, 1986. [Google Scholar]
- Payne, T.L.; Andryszak, N.A.; Wieser, H.; Dixon, E.A.; Ibrahim, N.; Coers, J. Antennnal olfactory and behavioral response of southern pine beetle, Dendroctonus frontalis, to analogs of its aggregation pheromone frontalin. J. Chem. Ecol. 1988, 14, 1217–1225. [Google Scholar] [CrossRef]
- Sullivan, B.T.; Shepherd, W.P.; Pureswaran, D.S.; Tashiro, T.; Mori, K. Evidence that (+)-endo-brevicomin is a male-produced component of the southern pine beetle aggregation pheromone. J. Chem. Ecol. 2007, 33, 1510–1527. [Google Scholar] [CrossRef]
- Francke, W.; Bartels, J.; Meyer, H.; Schröder, F.; Kohnle, U.; Baader, E.; Vité, J.P. Semiochemicals from bark beetles: New results, remarks, and reflections. J. Chem. Ecol. 1995, 21, 1043–1063. [Google Scholar] [CrossRef]
- Coster, J.E.; Vité, J.P. Effects of feeding and mating on pheromone release in the southern pine beetle. Ann. Entomol. Soc. Am. 1972, 65, 263–266. [Google Scholar] [CrossRef]
- McCarty, F.A.; Billings, P.; Richerson, J.V.; Payne, T.L.; Edson, L.J. Response of the southern pine beetle to behavioral chemicals in the laboratory. J. Entomol. Sci. 1980, 15, 307–317. [Google Scholar]
- Vité, J.P.; Bakke, A.; Renwick, J.A.A. Pheromones in Ips (Coleoptera: Scolytidae): Occurrence and production. Can. Entomol. 1972, 104, 1967–1975. [Google Scholar] [CrossRef]
- Byers, J.A.; Birgersson, G. Pheromone production in a bark beetle independent of myrcene precursor in host pine species. Sci. Nat. 1990, 77, 385–387. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Chen, Z.; Gaylord, M.L.; McMillin, J.D.; Wagner, M.R. Synergistic effects of α-pinene and exo-brevicomin on pine bark beetles and associated insects in Arizona. J. Appl. Entomol. 2008, 132, 387–397. [Google Scholar] [CrossRef]
- Miller, D.R. Ethanol and (−)-α-pinene: Attractant kairomones for some large wood-boring beetles in southeastern USA. J. Chem. Ecol. 2006, 32, 779–794. [Google Scholar] [CrossRef] [PubMed]
- Hanks, L.M.; Millar, J.G.; Moreira, J.A.; Barbour, J.D.; Lacey, E.S.; McElfresh, J.S.; Reuter, F.R.; Ray, A.M. Using generic pheromone lures to expedite identification of aggregation pheromones for the cerambycid beetles Xylotrechus nauticus, Phymatodes lecontei, and Neoclytus modestus modestus. J. Chem. Ecol. 2007, 33, 889–907. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef]
- Molander, M.A.; Winde, I.B.; Burman, J.; Nyabuga, F.N.; Lindblom, T.U.T.; Hanks, L.M.; Millar, J.G.; Larsson, M.C. Common cerambycid pheromone components as attractants for longhorn beetles (Cerambycidae) breeding in ephemeral oak substrates in northern Europe. J. Chem. Ecol. 2019, 45, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Pajares, J.A.; Álvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef]
- Teale, S.A.; Wickham, J.D.; Zhang, F.; Su, J.; Chen, Y.; Xiao, W.; Hanks, L.M.; Millar, J.G. A male-produced aggregation pheromone of Monochamus alternatus (Coleoptera: Cerambycidae), a major vector of pine wood nematode. J. Econ. Entomol. 2011, 104, 1592–1598. [Google Scholar] [CrossRef]
- Miller, D.R.; Allison, J.D.; Crowe, C.M.; Dickinson, D.M.; Eglitis, A.; Hofstetter, R.W.; Munson, A.S.; Poland, T.M.; Reid, L.S.; Steed, B.E.; et al. Pine sawyers (Coleoptera: Cerambycidae) attracted to α-pinene, monochamol, and ipsenol in North America. J. Econ. Entomol. 2016, 109, 1205–1214. [Google Scholar] [CrossRef]
- Schröder, F.; Fettköther, R.; Noldt, U.; Dettner, K.; König, W.A.; Francke, W. Synthesis of (3R)-3-hydroxy-2-hexanone, (2R,3R)-2,3-hexanediol and (2S,3R)-2,3-hexanediol, the male sex pheromone of Hylotrupes bajulus and Pyrrhidium sanguineum (Cerambycidae). Eur. J. Org. Chem. 1994, 1994, 1211–1218. [Google Scholar] [CrossRef]
- Miller, D.R.; Asaro, C. Predators attracted to combination of bark beetle pheromones and host kairomones in pine forests of southeastern United States. Environ. Entomol. 2023, 52, 787–794. [Google Scholar] [CrossRef]
- Peschke, K.; Friedrich, P.; Kaiser, U.; Franke, S.; Francke, W. Isopropyl (Z9) -hexadecenoate as a male attractant pheromone from the sternal gland of the rove beetle Aleochara curtala (Coleoptera: Staphylinidae). Chemoecology 1999, 9, 47–54. [Google Scholar] [CrossRef]
- Avşar, İ.; Turan, Y. An overview of the pheromones of Staphylinidae (Coleoptera). Trans. Am. Entomol. Soc. 2024, 150, 205–215. [Google Scholar] [CrossRef]
- Tóth, M.; Furlan, L.; Yatsynin, V.G.; Ujváry, I.; Szarukán, I.; Imrei, Z.; Subchev, M.; Tolasch, T.; Francke, W. Identification of sex pheromone composition of click beetle Agriotes brevis candeze. J. Chem. Ecol. 2002, 28, 1641–1652. [Google Scholar] [CrossRef]
- Tóth, M.; Furlan, L.; Yatsynin, V.G.; Ujváry, I.; Szarukán, I.; Imrei, Z.; Tolasch, T.; Francke, W.; Jossi, W. Identification of pheromones and optimization of bait composition for click beetle pests (Coleoptera: Elateridae) in Central and Western Europe. Pest Manag. Sci. 2003, 59, 417–425. [Google Scholar] [CrossRef]
- Amman, G.D. The role of the mountain pine beetle in lodgepole pine ecosystems: Impact on succession. In The Role of Arthropods in Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 1977; pp. 3–18. [Google Scholar]
- Safranyik, L.; Carroll, A.L. The biology and epidemiology of the mountain pine beetle in lodgepole pine forests. In The Mountain Pine Beetle: A Synthesis of its Biology, Management and Impacts on Lodgepole Pine; Natural Resources Canada: Victoria, BC, Canada, 2006; pp. 3–66. [Google Scholar]
- Cudmore, T.J.; Björklund, N.; Carroll, A.L.; Lindgren, B.S. Climate change and range expansion of an aggressive bark beetle: Evidence of higher beetle reproduction in naïve host tree populations. J. Appl. Ecol. 2010, 47, 1036–1043. [Google Scholar] [CrossRef]
- Sambaraju, K.R.; Goodsman, D.W. Mountain pine beetle: An example of a climate-driven eruptive insect impacting conifer forest ecosystems. CAB Rev. Perspect. Agric. Veter-Sci. Nutr. Nat. Resour. 2021, 16, 1–18. [Google Scholar] [CrossRef]
- Amman, G.D. Prey consumption and variations in larval biology of Enoclerus spegeus (Coleoptera: Cleridae). Can. Entomol. 1970, 102, 1374–1379. [Google Scholar] [CrossRef]
- Krause, A.M.; Townsend, P.A.; Lee, Y.; Raffa, K.F. Predators and competitors of the mountain pine beetle Dendroctonus ponderosae (Coleoptera: Curculionidae) in stands of changing forest composition associated with elevation. Agric. For. Entomol. 2018, 20, 402–413. [Google Scholar] [CrossRef]
- Carroll, A.L.; Aukema, B.H.; Raffa, K.F.; Linton, D.A.; Smith, G.D.; Lindgren, B.S. Mountain pine beetle outbreak development: The endemic—Incipient epidemic transition. In Mountain Pine Beetle Initiative Project; Canadian Forest Service: Marie, ON, Canada, 2006; Volume 1, p. 22. [Google Scholar]
- Bleiker, K.P.; Six, D.L. Dietary benefits of fungal associates to an eruptive herbivore: Potential implications of multiple associates on host population dynamics. Environ. Entomol. 2007, 36, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Goodsman, D.W.; Erbilgin, N.; Lieffers, V.J. The impact of phloem nutrients on overwintering mountain pine beetles and their fungal symbionts. Environ. Entomol. 2012, 41, 478–486. [Google Scholar] [CrossRef]
- Therrien, J.; Mason, C.J.; Cale, J.A.; Adams, A.; Aukema, B.H.; Currie, C.R.; Raffa, K.F.; Erbilgin, N. Bacteria influence mountain pine beetle brood development through interactions with symbiotic and antagonistic fungi: Implications for climate-driven host range expansion. Oecologia 2015, 179, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Cale, J.A.; Muskens, M.; Najar, A.; Ishangulyyeva, G.; Hussain, A.; Kanekar, S.S.; Klutsch, J.G.; Taft, S.; Erbilgin, N.; Schnitzler, J.-P. Rapid monoterpene induction promotes the susceptibility of a novel host pine to mountain pine beetle colonization but not to beetle-vectored fungi. Tree Physiol. 2017, 37, 1597–1610. [Google Scholar] [CrossRef]
- Guevara-Rozo, S.; Hussain, A.; Cale, J.A.; Klutsch, J.G.; Rajabzadeh, R.; Erbilgin, N. Nitrogen and ergosterol concentration varied in live jack pine phloem following inoculations with fungal associates of mountain pine beetle. Front. Microbiol. 2020, 11, 1703. [Google Scholar] [CrossRef] [PubMed]
- Cale, J.A.; Collignon, R.M.; Klutsch, J.G.; Kanekar, S.S.; Hussain, A.; Erbilgin, N. Fungal volatiles can act as carbon sources and semiochemicals to mediate interspecific interactions among bark beetle-associated fungal symbionts. PLoS ONE 2016, 11, e0162197. [Google Scholar] [CrossRef]
- Chiu, C.C.; Bohlmann, J. Mountain pine beetle epidemic: An interplay of terpenoids in host defense and insect pheromones. Annu. Rev. Plant Biol. 2022, 73, 475–494. [Google Scholar] [CrossRef]
- Moore, M.L.; Six, D.L. Effects of temperature on growth, sporulation, and competition of mountain pine beetle fungal symbionts. Microb. Ecol. 2015, 70, 336–347. [Google Scholar] [CrossRef]
- Aukema, B.H.; Dahlsten, D.L.; Raffa, K.F. Improved population monitoring of bark beetles and predators by incorporating disparate behavioral responses to semiochemicals. Environ. Entomol. 2000, 29, 618–629. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Gaylord, M.L.; Martinson, S.; Wagner, M.R. Attraction to monoterpenes and beetle-produced compounds by syntopic Ips and Dendroctonus bark beetles and their predators. Agric. For. Entomol. 2012, 14, 207–215. [Google Scholar] [CrossRef]
- Amman, G.D. Mountain pine beetle brood production in relation to thickness of lodgepole pine phloem. J. Econ. Entomol. 1972, 65, 138–140. [Google Scholar] [CrossRef]
- Erbilgin, N.; Ma, C.; Whitehouse, C.; Shan, B.; Najar, A.; Evenden, M. Chemical similarity between historical and novel host plants promotes range and host expansion of the mountain pine beetle in a naïve host ecosystem. New Phytol. 2013, 201, 940–950. [Google Scholar] [CrossRef]
- Wijerathna, A.; Evenden, M. Effect of environmental conditions on flight capacity in mountain pine beetle (Coleoptera: Curculionidae: Scolytinae). J. Insect Behav. 2020, 33, 201–215. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subcortical Beetle Community | Known Attractants | Source | Species Attracted | References |
|---|---|---|---|---|
| Curculionidae, Scolytinae | trans-Verbenol | Female pheromone | Dendroctonus ponderosae | [32,33] |
| exo-Brevicomin | Male pheromone | D. ponderosae, D. terebrans, D. brevicomis | [34] | |
| (-)-endo-Brevicomin | Male pheromone | D. frontalis | [35,36] | |
| (+)-Sulcatol | Female pheromone | Gnathotrichus sulcatus; Ips sexdentatus | [37] | |
| Frontalin | Female pheromone | D. rufipennis, D. brevicomis, D. pseudotsugae | [35,38,39] | |
| Ipsdienol | Male pheromone | Ips spp. | [26,40,41] | |
| α-Pinene | Host tree volatile | D. frontalis, D. brevicomis, Ips spp., Hylastes porculus | [18,32,42,43] | |
| Cerambycidae | 2-Methyl-1-butanol | Pheromone component | Neoclytus, Xylotrechus spp., Monochamus spp. | [44,45,46] |
| Ethanol | Similar compound to male pheromone | Monochamus spp., Xylotrechus spp. | [43,47,48] | |
| Monochamol | Male aggregation pheromone | Monochamus spp. | [49] | |
| 3-Hydroxy-2-hexanone | Male aggregation pheromone | Various Cerambycinae species | [46,50] | |
| Buprestidae | α-Pinene | Monoterpene | Buprestis lineata | [23,43,51] |
| Staphylinidae | Isopropyl (Z9)-hexadecenoate | Female aggregation pheromone | Aleochara curtula | [52,53] |
| Elateridae | Ethanol | Released by stressed trees | Alaus myops | [43] |
| Geranyl butanoate | Female aggregation pheromone | Agriotes spp. | [54,55] |
| Treatments/Chemical | Chemical Purity (%) | Concentrations (µL mL−1) | Release Rate * (mg day−1) |
|---|---|---|---|
| Acetoin | ≥96 | 61.87 | 1.48 |
| 3-Methyl-1-butanol | 98 | 39.87 | 0.98 |
| 2-Methyl-1-butanol | ≥99 | 20.92 | 0.56 |
| Isobutanol | ≥99 | 30.33 | 0.62 |
| 2-Methyl-2-butanol | 99 | 33.27 | 0.84 |
| FVOC mixture | 48.39 | 0.96 | |
| Mineral oil |
| Compounds | Enantiomeric Ratios | Chemical Purity (%) | Source |
|---|---|---|---|
| Geranyl acetate | ≥97 | Sigma-Aldrich | |
| α-Pinene | (−) | 98 | Sigma-Aldrich |
| Camphene | (−) | 90 | SAFC (Kent Town, Australia) |
| β-Pinene | (+) | ≥94 | TCI Chemicals (Tokyo, Japan) |
| 3-Carene | 90 | Sigma-Aldrich | |
| β-Myrcene | 90 | Sigma-Aldrich | |
| Limonene | (S)-(−) | 96 | Fluka Analytical (Buchs, Switzerland) |
| Terpinolene | ≥90 | SAFC | |
| Bornyl acetate | ≥99 | SAFC | |
| γ-Terpinene | 97 | Fluka Analytical | |
| Camphor | ≥95 | Fluka Analytical | |
| Borneol | (−) | 97 | Sigma-Aldrich |
| Group | Treatments | Scolytinae | Buprestidae | Cerambycidae | Staphylinidae | Elateridae |
|---|---|---|---|---|---|---|
| FVOC | 2-Methyl-1-butanol | 0 | 4 | 36 | 1 | 3 |
| 2-Methyl-2-butanol | 0 | 1 | 27 | 3 | 3 | |
| 3-Methyl-1-butanol | 0 | 2 | 31 | 1 | 1 | |
| Acetoin | 0 | 1 | 17 | 3 | 3 | |
| Isobutanol | 0 | 1 | 20 | 4 | 2 | |
| Control- Mineral oil | 0 | 2 | 19 | 2 | 3 | |
| Mixture | 0 | 0 | 21 | 4 | 3 | |
| Total | 0 | 11 | 171 | 18 | 18 | |
| FVOC with pheromones | 2-Methyl-1-butanol | 148 | 2 | 15 | 4 | 3 |
| 2-Methyl-2-butanol | 118 | 2 | 8 | 1 | 7 | |
| 3-Methyl-1-butanol | 83 | 2 | 6 | 2 | 5 | |
| Acetoin | 95 | 2 | 45 | 3 | 2 | |
| Isobutanol | 85 | 4 | 15 | 1 | 1 | |
| Control- Pheromone alone | 74 | 2 | 18 | 1 | 5 | |
| Mixture | 91 | 3 | 35 | 0 | 5 | |
| Total | 694 | 17 | 142 | 12 | 28 | |
| SVC | AP | 5 | 1 | 17 | 9 | 7 |
| Control- Mineral oil | 6 | 2 | 14 | 9 | 6 | |
| GC | 4 | 0 | 16 | 15 | 2 | |
| Healthy | 3 | 0 | 16 | 13 | 15 | |
| LL | 5 | 2 | 21 | 5 | 0 | |
| OM | 4 | 1 | 13 | 6 | 7 | |
| EH | 3 | 0 | 26 | 5 | 13 | |
| Total | 30 | 6 | 123 | 62 | 50 | |
| Total Catches | 724 | 34 | 436 | 92 | 96 |
| Subcortical Beetle Family | Known Attractants | Key FVOCs | Key FVOCs When Pheromones Are Added | Key SVC Profiles |
|---|---|---|---|---|
| Curculionidae Sub-family: Scolytinae | trans-Verbenol, exo-Brevicomin, Terpinolene, Myrcene | Low catch | 2-Methyl-1-butanol, 2-Methyl-2-butanol | Low catch |
| Cerambycidae | 2-Methyl-1-butanol, ⍺-Pinene | 2-Methyl-1-butanol | Acetoin, blend | Endocronartium harknessii |
| Buprestidae | Tree stress volatiles primarily | 2-Methyl-1-butanol | Isobutanol | Leptographium longiclavatum |
| Staphylinidae | Bark beetle pheromones, some monoterpenes | Isobutanol, blend | 2-Methyl-1-butanol, Acetoin | Grosmannia clavigera, Healthy tree |
| Elateridae | Bark beetle pheromones, some monoterpenes | Low catch | 2-Methyl-2-butanol | Healthy tree, Endocronartium harknessii |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crandall, L.; Zaman, R.; Duthie-Holt, M.; Jarvis, W.; Erbilgin, N. Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles. Insects 2025, 16, 57. https://doi.org/10.3390/insects16010057
Crandall L, Zaman R, Duthie-Holt M, Jarvis W, Erbilgin N. Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles. Insects. 2025; 16(1):57. https://doi.org/10.3390/insects16010057
Chicago/Turabian StyleCrandall, Leah, Rashaduz Zaman, Marnie Duthie-Holt, Wade Jarvis, and Nadir Erbilgin. 2025. "Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles" Insects 16, no. 1: 57. https://doi.org/10.3390/insects16010057
APA StyleCrandall, L., Zaman, R., Duthie-Holt, M., Jarvis, W., & Erbilgin, N. (2025). Navigating the Semiochemical Landscape: Attraction of Subcortical Beetle Communities to Bark Beetle Pheromones, Fungal and Host Tree Volatiles. Insects, 16(1), 57. https://doi.org/10.3390/insects16010057