
Body Size of Female Strepsipteran Parasites (Strepsiptera, Xenidae, Xenos) Depends on Several Key Factors in a Vespine Wasp (Hymenoptera, Vespidae, Vespa)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
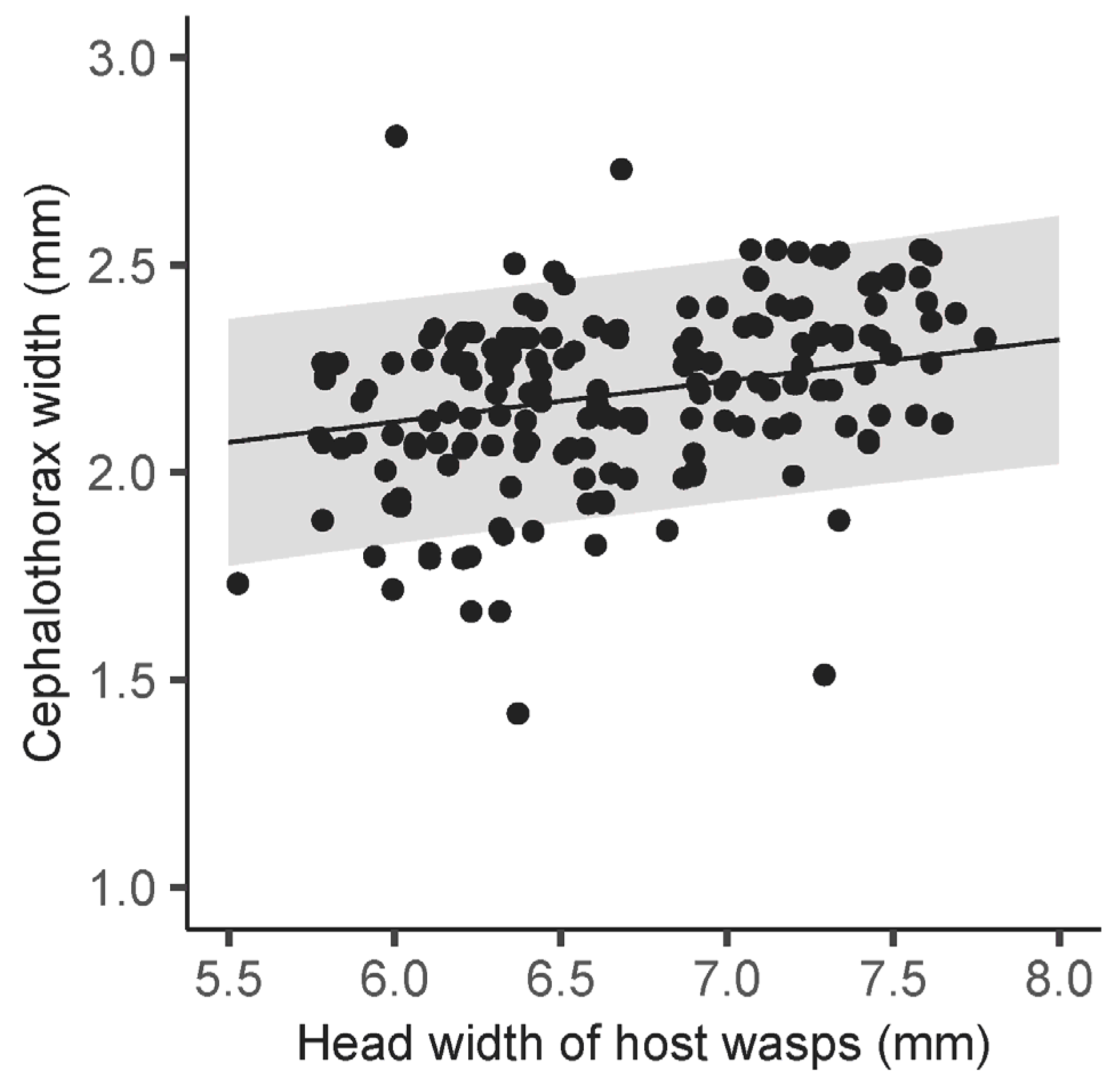
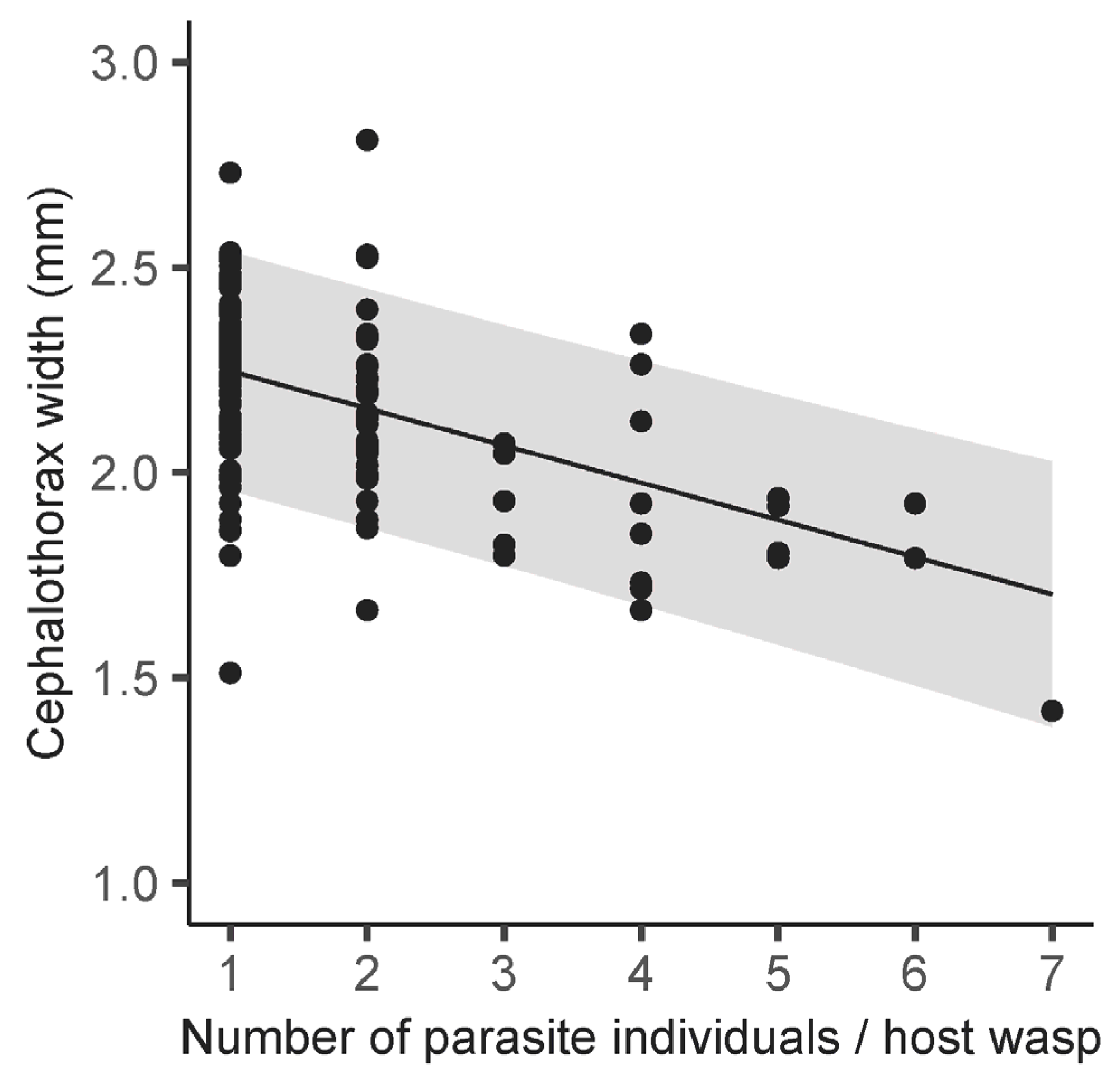
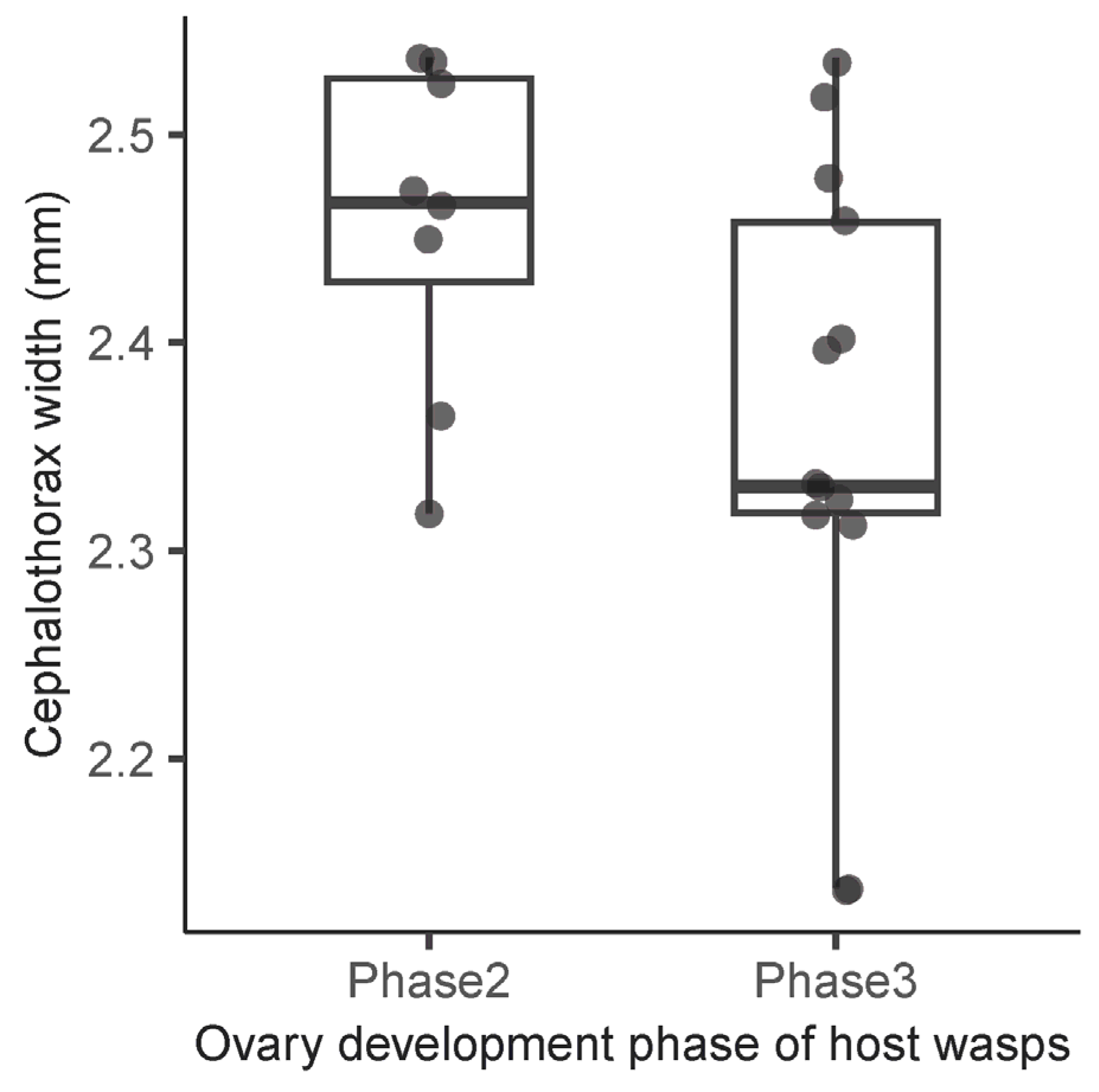
3. Results
4. Discussion
4.1. Body Size of Host Wasps and Seasonality
4.2. Number of Female Parasites per Host Wasp
4.3. Reproductive Conditions of Host Wasps
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmid-Hempel, P. Parasites in Social Insects; Princeton University Press: Princeton, NJ, USA, 1998; p. 409. [Google Scholar]
- Shykoff, J.A.; Schmid-Hempel, P. Parasite delay worker reproductionin the bumblebee: Consequences for Eusociality. Behav. Ecol. 1992, 2, 242–248. [Google Scholar] [CrossRef]
- Boots, M.; Bowers, R.G. Three mechanisms of host resistance to microparasites-Avoidance, recovery and tolerance show different evolutionary dynamics. J. Theor. Biol. 1999, 201, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Schmid-Hempel, P.; Ebert, D. On the evolutionary ecology of specific immune defence. Trends Ecol. Evol. 2003, 18, 27–32. [Google Scholar] [CrossRef]
- Kathirithamby, K. Review of the order Strepsiptera. Syst. Entomol. 1989, 14, 41–92. [Google Scholar] [CrossRef]
- Matsuura, M.; Yamane, S. Biology of the Vespine Wasps; Springer: Berlin/Heidelberg, Germany, 1990; p. 323. [Google Scholar]
- Matsuura, M. Notes on the number of eggs laid by Vespaxenos. Insectarium 1975, 12, 18. (In Japanese) [Google Scholar]
- Kinzelbach, R.K. Morphologische Befunde an Fächerflünglern und Ihrephylogenetische Bedeutung (Insecta: Strepsiptera); Science Publishers: Berlin, Germany, 1971; p. 256. [Google Scholar]
- Kifune, T. Detailed location of the Japanese strepsipterans. Trans. Essa Entomol. Soc. 1992, 74, 55–71. (In Japanese) [Google Scholar]
- Kifune, T.; Makino, S. The first record of Xenos moutoni from Kyush, Japan. Pulex 1996, 85, 459. (In Japanese) [Google Scholar]
- Nakase, Y.; Kato, M. Cryptic diversity and host specificity in giant Xenos Strepsipterans parasitic in large Vespa hornets. Zool. Sci. 2013, 30, 331–336. [Google Scholar] [CrossRef]
- Makino, S.; Yamashita, Y. Levels of parasitism by Xenos moutoni de Buysson (Strepsiptera, Stylopidae) and their seasonal changes in hornets (Hymenoptera: Vespidae, Vespa) caught with bait traps. J. Entomol. Sci. 1998, 1, 537–543. [Google Scholar]
- Makino, S. Seasonal changes in levels of parasitism and sex ratio of Xenos moutoni du Buysson (Strepsiptera, Stylopicae) in the Japanese hornet, Vespa analis insularis Dalla Torre (Hymenoptera, Vespidae). Tijdschr. Entomol. 2001, 144, 217–222. [Google Scholar] [CrossRef]
- Tatsuta, H.; Makino, S. Rate of strepsipteran parasitization among overwintered females of the hornet Vespa analis (Hymenoptera: Vespidae). Environ. Entomol. 2003, 32, 175–179. [Google Scholar] [CrossRef]
- Oyaizu, W.; Kudô, K. Seasonal changes in the number of vespine wasps and levels of parasitism by Xenos moutoni (Strepsiptera, Stylopicae) collected with attractant traps in Matunoyama forest, Tokamachi city, Japan. Bull. Fac. Edu. (Nat. Sci.) Niigata Univ. 2013, 6, 49–57, (In Japanese with English Summary). [Google Scholar]
- Kanzaki, N.; Makino, S.; Kosaka, H.; Sayama, K.; Hamaguchi, K.; Narayama, S. Nematode and strepsipteran parasitism in bait-trapped and hand-collected hornets (Hymenoptera, Vespidae, Vespa). Insects 2023, 14, 398. [Google Scholar] [CrossRef]
- Kudô, K.; Oyaizu, W.; Kusama, R.; Yamagishi, K.; Yamaguchi, Y.; Koji, S. Mating and post-hibernation ovarian development in stylopized and non-stylopized queens of the hornet Vespa analis (Hymenoptera Vespidae). Ethol. Ecol. Evol. 2024, 36, 86–95. [Google Scholar] [CrossRef]
- Makino, S.; Yamaura, Y.; Yamauchi, H. Smaller nests of the hornet Vespa analis (Hymenoptera, Vespidae) are more severely affected by the strepsipteran parasite Xenos moutoni (Strepsiptera, Stylopidae) than are larger nests. Insectes Soc. 2010, 57, 83–90. [Google Scholar] [CrossRef]
- Kudô, K.; Yamane, S.; Mateus, S.; Tsuchida, K.; Itô, Y.; Miyano, S.; Zucchi, R. Parasitism affects worker size in the Neotropical swarm-founding social wasp, Polybia paulista. Insectes Soc. 2004, 51, 221–225. [Google Scholar] [CrossRef]
- Kudô, K.; Ohka, H.; Zucchi, R. Modification of morphological characteristics by endoparasites in workers of the swarm-founding wasp Polybia occidentalis. In Social Insects Structure, Function, and Behavior; Nova Science Publishers, Inc.: New York, NY, USA, 2011; pp. 83–96. [Google Scholar]
- Beani, L.; Dallai, R.; Cappa, F.; Manfredini, F.; Zaccaroni, M.; Lorenzi, M.C.; Mercati, D. A strepsipteran parasite extends the lifespan of workers in a social wasp. Sci. Rep. 2021, 11, 7235. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023. [Google Scholar]
- Maeta, Y.; Kifune, T. Biology of Strepsiptera (2)–(5). Insectarium 1990, 27, 198–205, 240–247, 278–286. (In Japanese) [Google Scholar]
- Chole, H.; Woodard, S.H.; Bloch, G. Body size variation in bees: Regulation, mechanism, and relationship to social organization. Curr. Opin. Insect Sci. 2019, 35, 77–87. [Google Scholar] [CrossRef]
- Maeta, Y. Some biological notes on Pseudoxenos iwatai Esaki (I) Its Biology and Life History. Kontyû 1963, 31, 3–15, (In Japanese with English Summary). [Google Scholar]
- Maeta, Y.; Takahashi, K.; Shimada, N. Hosts body size as a factor for determining the egg complement of Strepsiptera, an insect parasite. Int. J. Insect Morphol. Embryol. 1998, 22, 27–37. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Estimate | SE | t |
|---|---|---|---|
| (Intercept) | 1.68 | 0.18 | 9.06 |
| HW of host wasps | 0.10 | 0.03 | 3.75 |
| Number of parasites per host | −0.09 | 0.01 | −6.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudô, K.; Oyaizu, W.; Kusama, R.; Yamaguchi, Y.; Koji, S. Body Size of Female Strepsipteran Parasites (Strepsiptera, Xenidae, Xenos) Depends on Several Key Factors in a Vespine Wasp (Hymenoptera, Vespidae, Vespa). Insects 2024, 15, 266. https://doi.org/10.3390/insects15040266
Kudô K, Oyaizu W, Kusama R, Yamaguchi Y, Koji S. Body Size of Female Strepsipteran Parasites (Strepsiptera, Xenidae, Xenos) Depends on Several Key Factors in a Vespine Wasp (Hymenoptera, Vespidae, Vespa). Insects. 2024; 15(4):266. https://doi.org/10.3390/insects15040266
Chicago/Turabian StyleKudô, Kazuyuki, Wataru Oyaizu, Rikako Kusama, Yuki Yamaguchi, and Shinsaku Koji. 2024. "Body Size of Female Strepsipteran Parasites (Strepsiptera, Xenidae, Xenos) Depends on Several Key Factors in a Vespine Wasp (Hymenoptera, Vespidae, Vespa)" Insects 15, no. 4: 266. https://doi.org/10.3390/insects15040266
APA StyleKudô, K., Oyaizu, W., Kusama, R., Yamaguchi, Y., & Koji, S. (2024). Body Size of Female Strepsipteran Parasites (Strepsiptera, Xenidae, Xenos) Depends on Several Key Factors in a Vespine Wasp (Hymenoptera, Vespidae, Vespa). Insects, 15(4), 266. https://doi.org/10.3390/insects15040266