Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
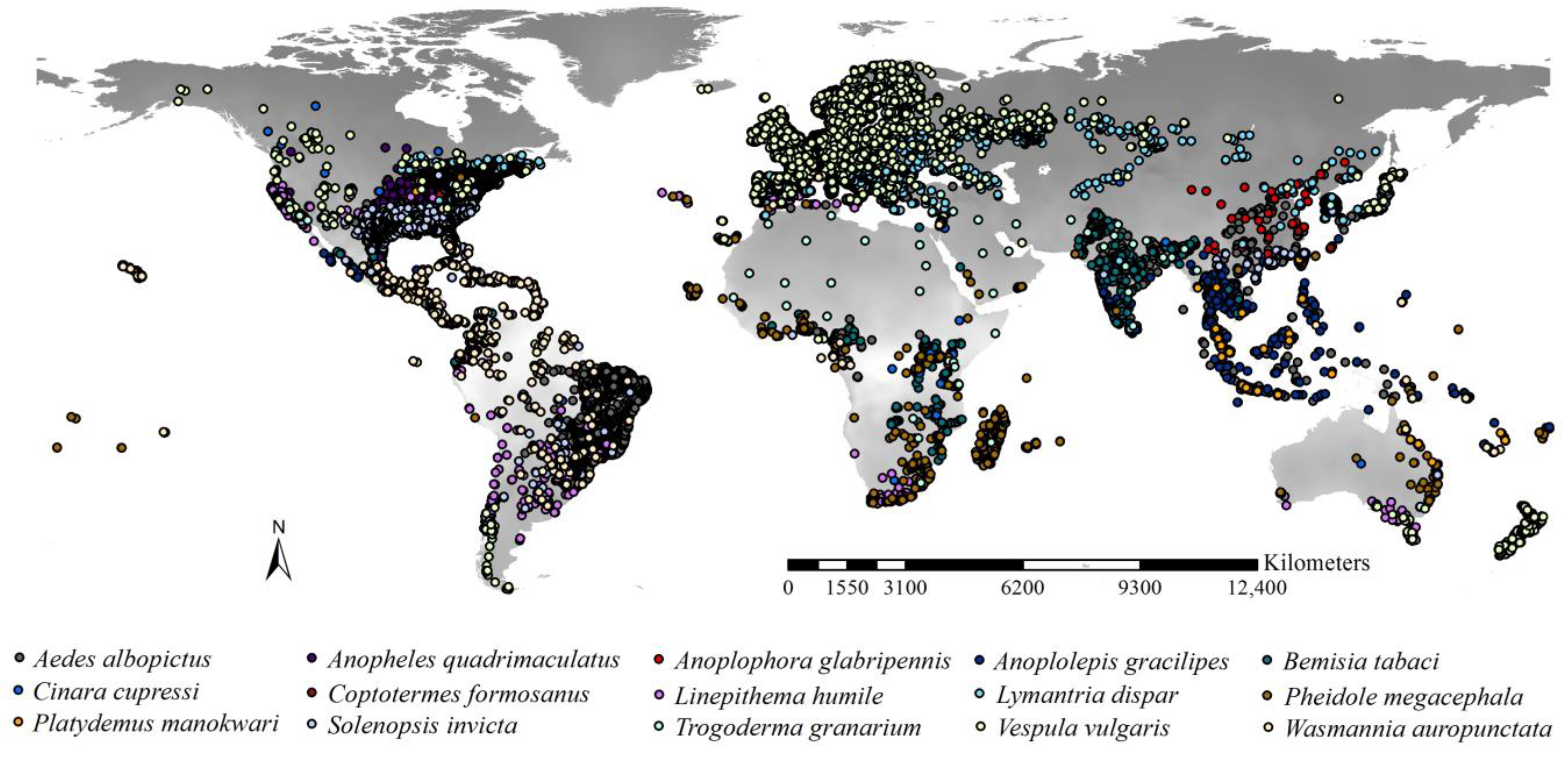
2.1. Occurrences of the Worst IIPs
2.2. Predictors in the Range Dynamic Models
2.3. Predictor Selection
2.4. Estimating Habitat Suitability and Potential Ranges
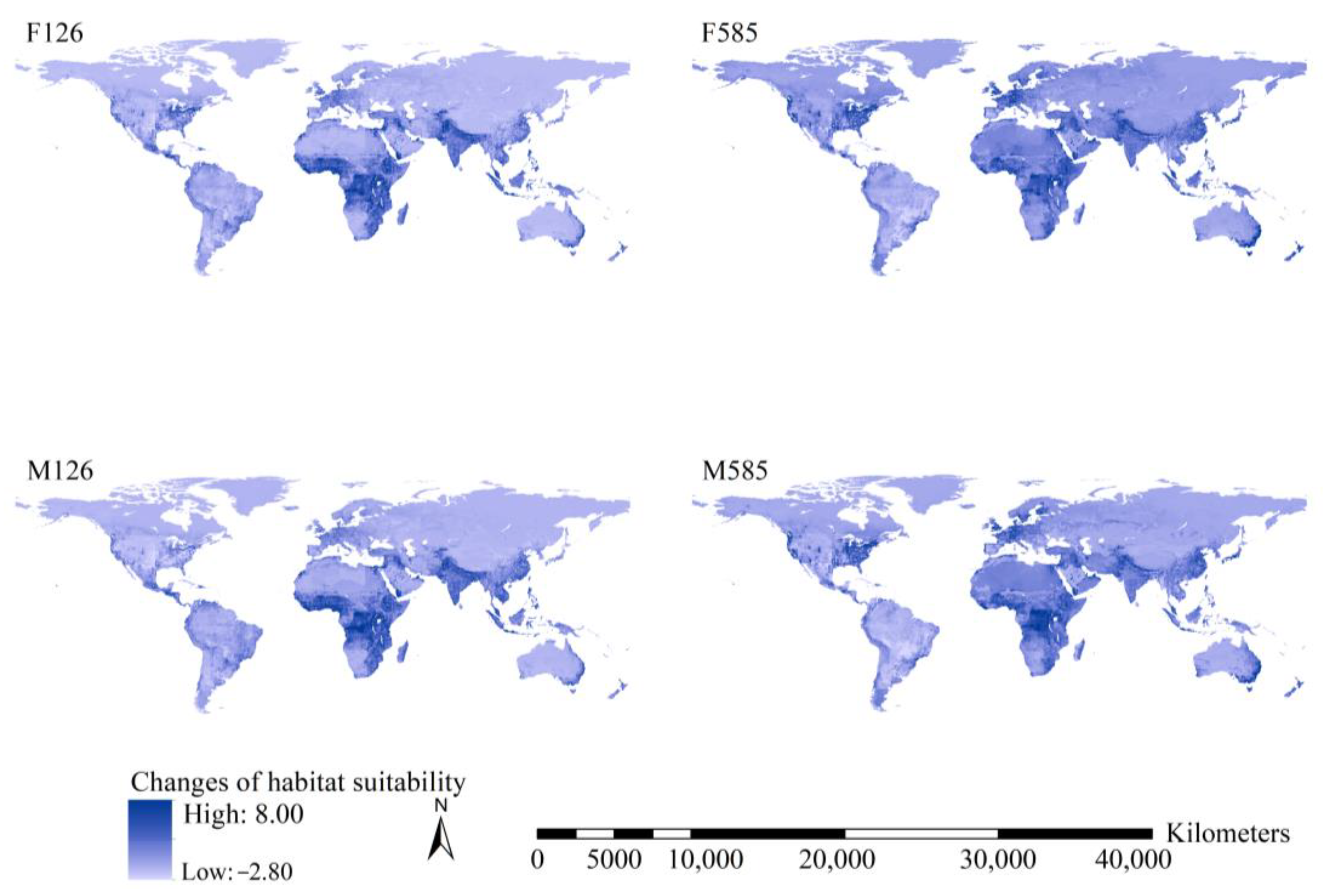
2.5. Dynamics in Habitat Suitability
2.6. Estimating Range Dynamics
3. Results
3.1. Model Performance
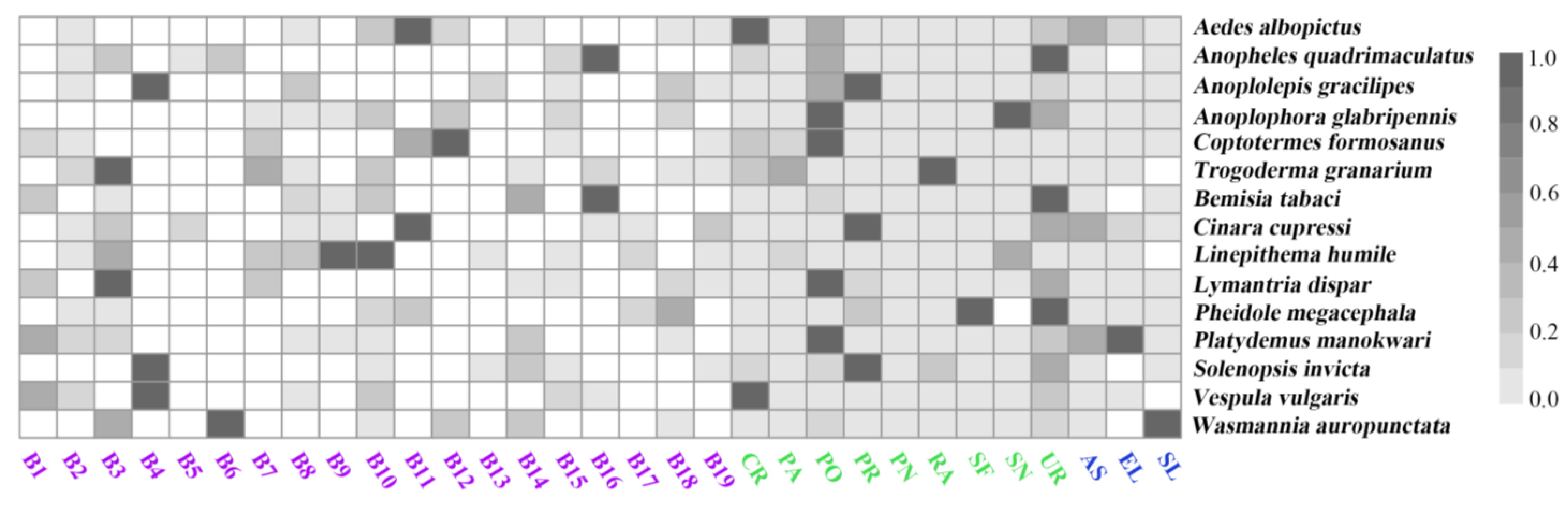
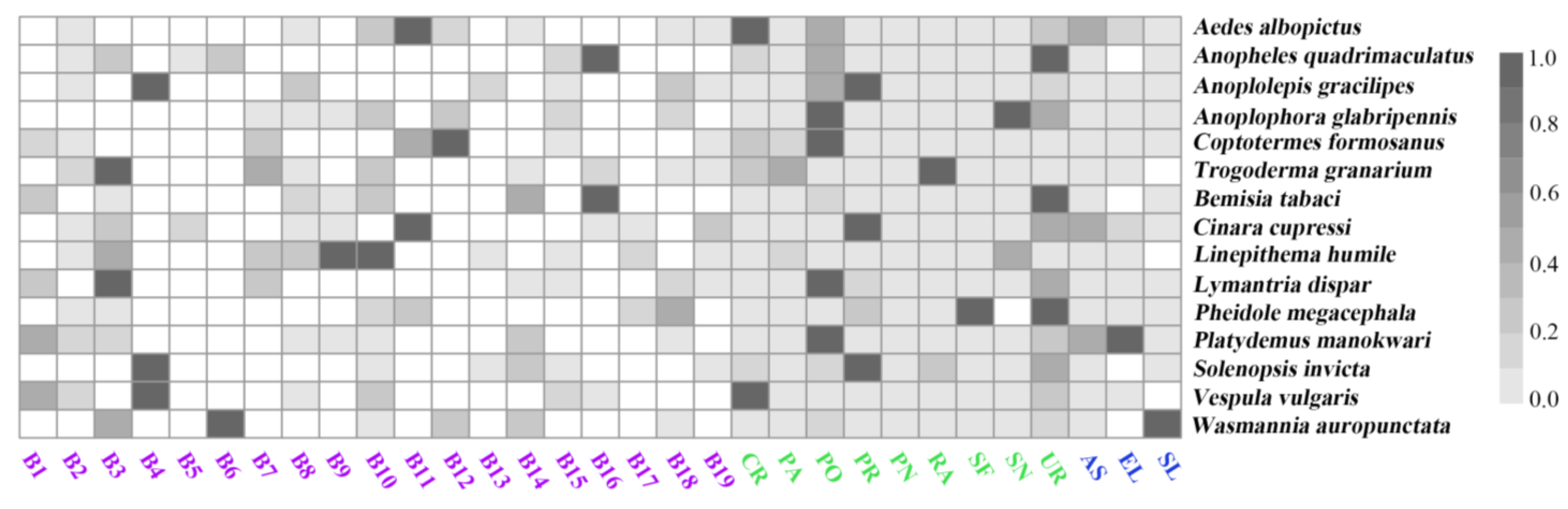
3.2. Importances of the Predictors
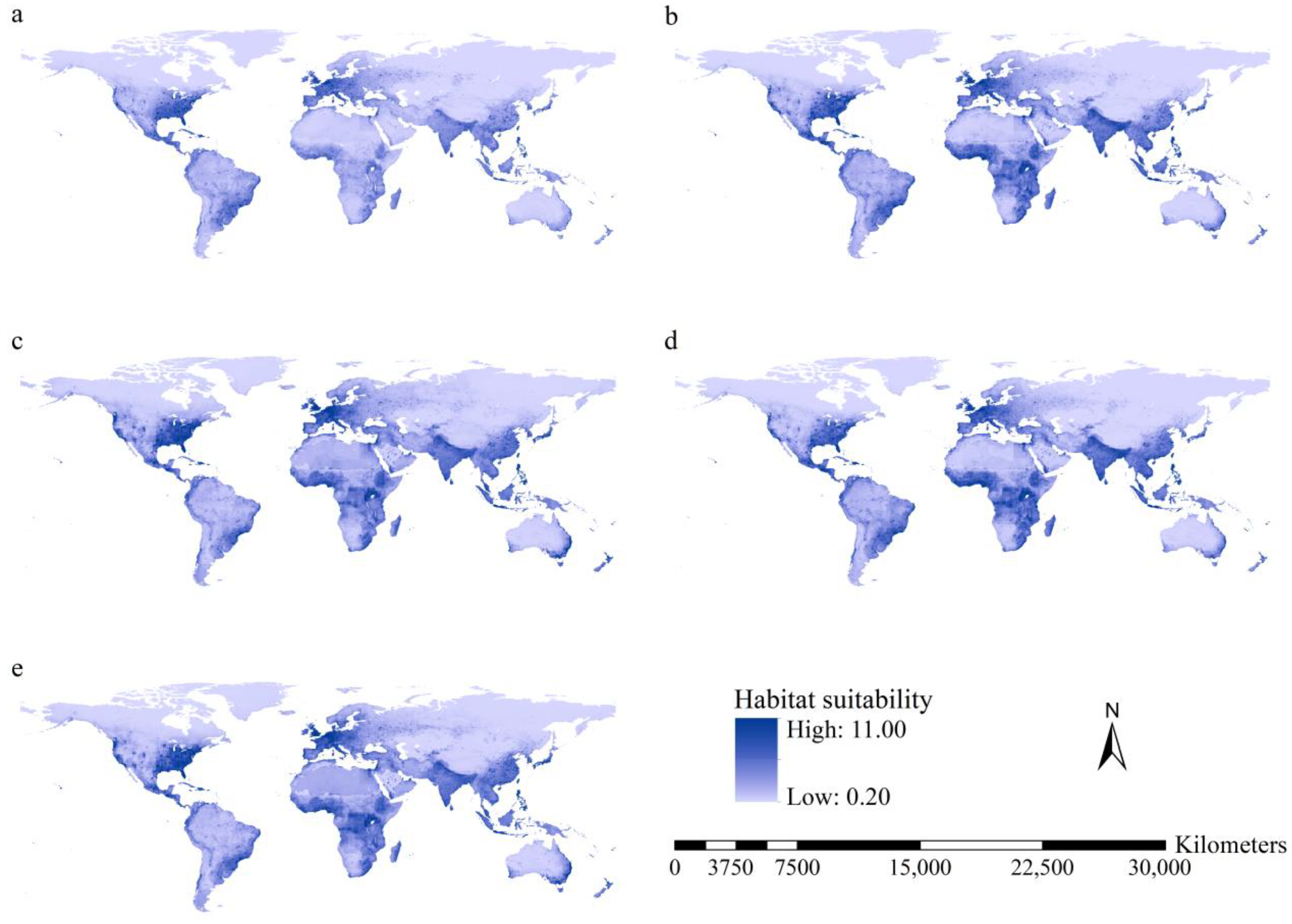
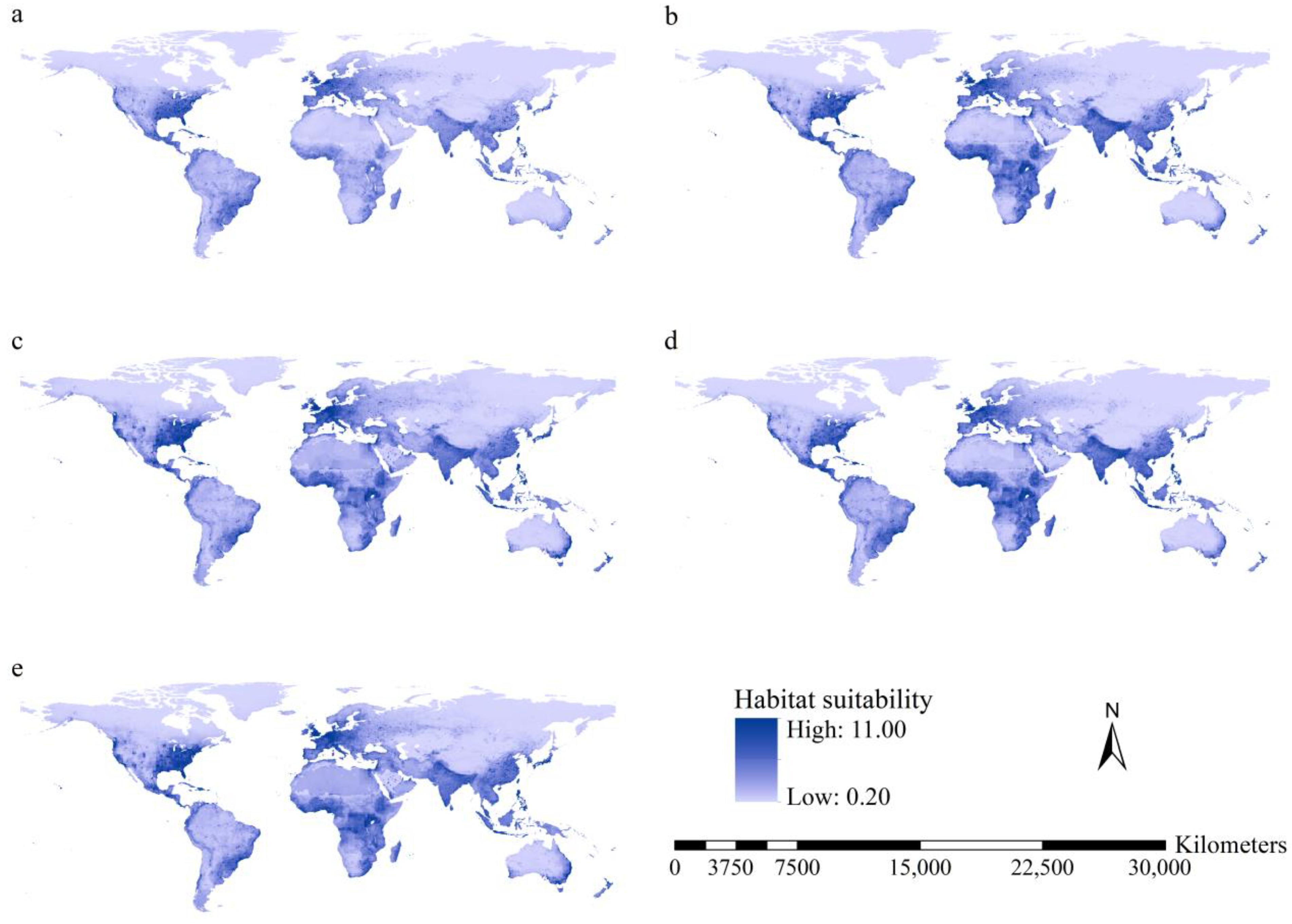
3.3. Habitat Suitability of the Worst IIPs
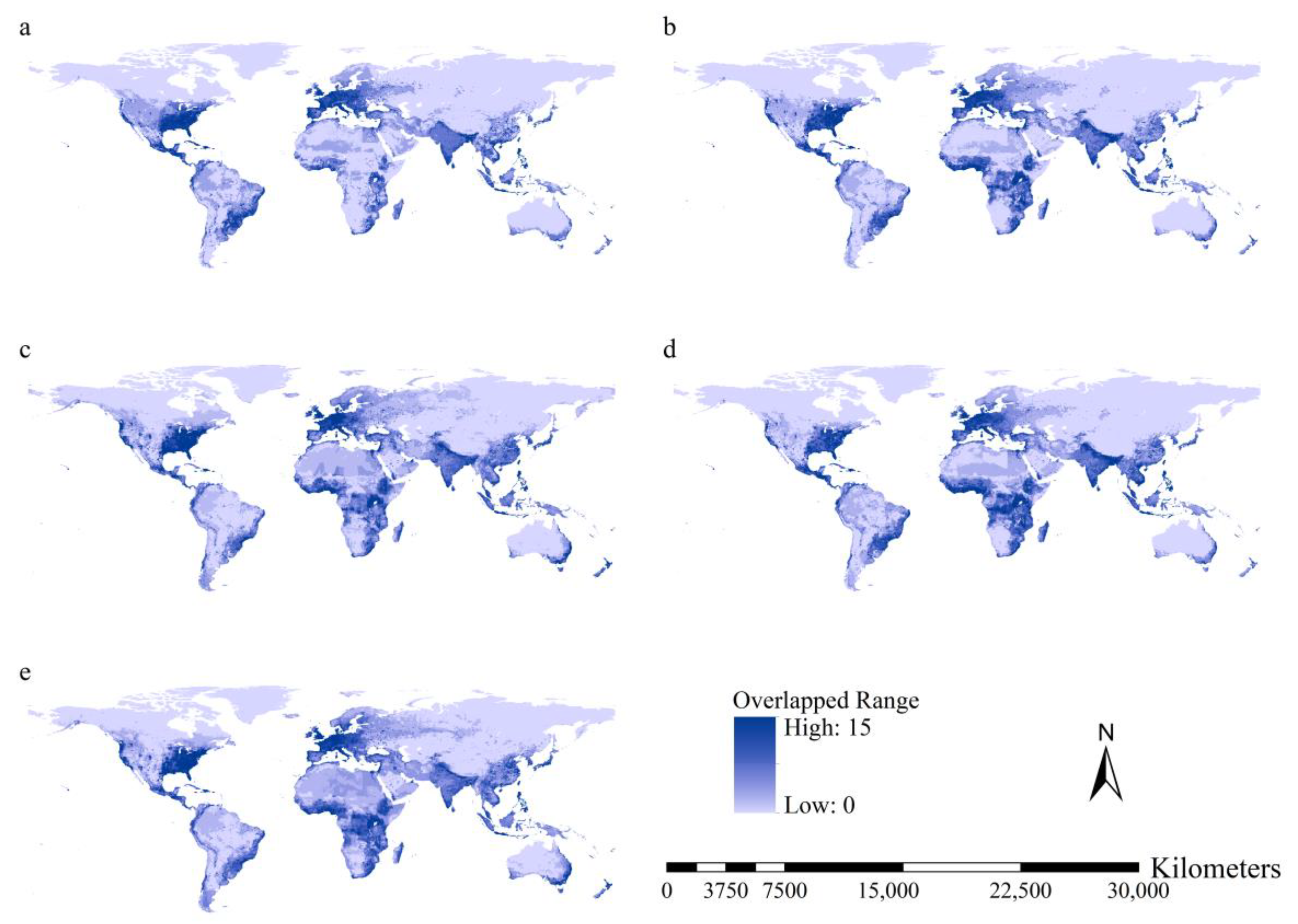
3.4. Potential Ranges of the Worst IIPs
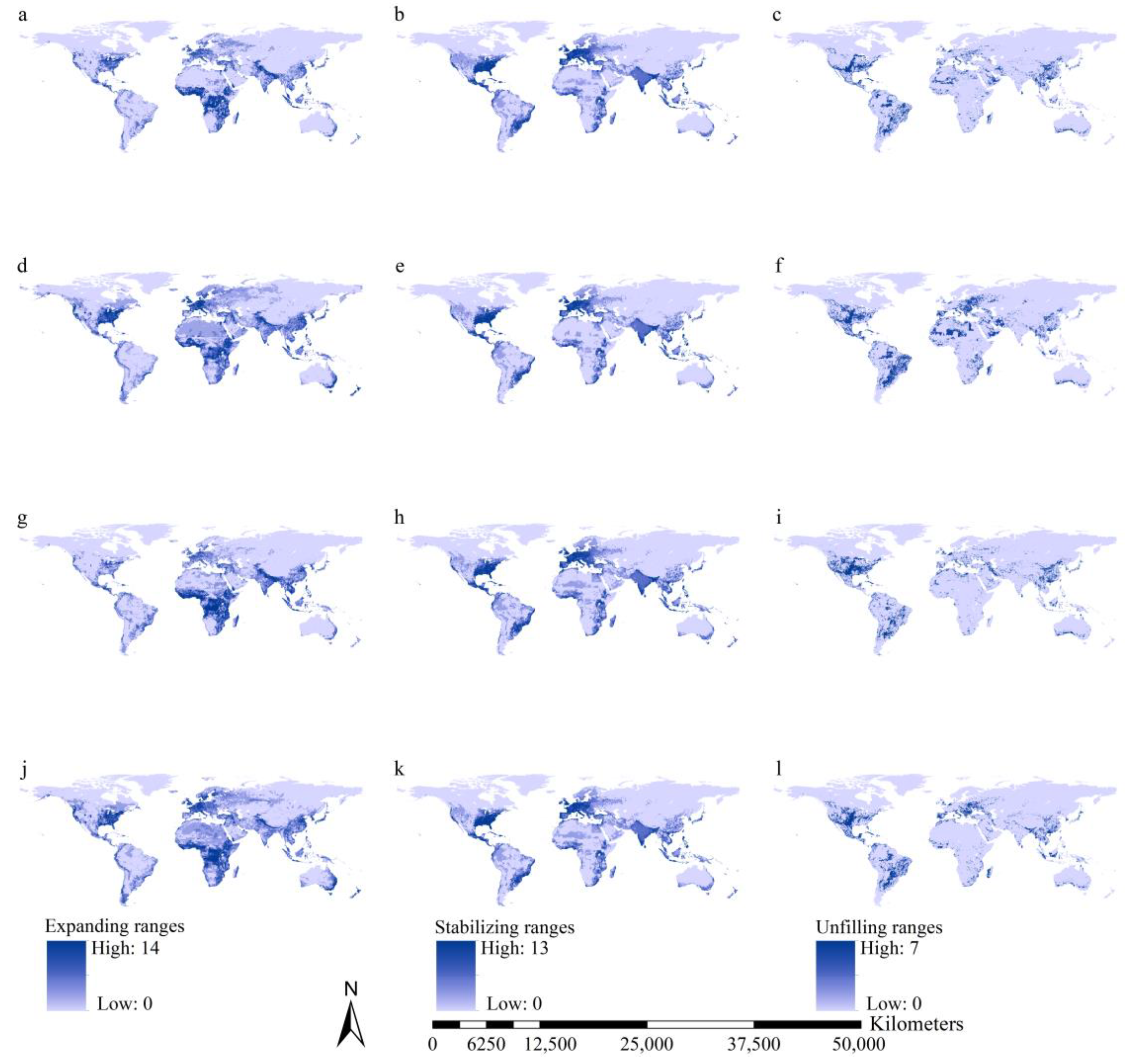
3.5. Range Dynamics of the Worst IIPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sileshi, G.W.; Gebeyehu, S.; Mafongoya, P.L. The threat of alien invasive insect and mite species to food security in Africa and the need for a continent-wide response. Food Secur. 2019, 11, 763–775. [Google Scholar] [CrossRef]
- Koop, J.A.H.; Causton, C.E.; Bulgarella, M.; Cooper, E.; Heimpel, G.E. Population structure of a nest parasite of Darwin’s finches within its native and invasive ranges. Conserv. Genet. 2020, 22, 11–22. [Google Scholar] [CrossRef]
- Venette, R.C.; Hutchison, W.D. Invasive insect species: Global challenges, strategies & opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar]
- Jankovic, M.; Petrovskii, S. Gypsy moth invasion in North America: A simulation study of the spatial pattern and the rate of spread. Ecol. Complex. 2013, 14, 132–144. [Google Scholar] [CrossRef]
- Srivastava, V.; Griess, V.C.; Keena, M.A. Assessing the Potential Distribution of Asian Gypsy Moth in Canada: A Comparison of Two Methodological Approaches. Sci. Rep. 2020, 10, 22. [Google Scholar] [CrossRef]
- Song, J.W.; Jung, J.M.; Nam, Y.W.; Jung, J.K.; Jung, S.H.; Lee, W.H. Spatial ensemble modeling for predicting the poten-tial distribution of Lymantria dispar asiatica (Lepidoptera: Erebidae: Lymantriinae) in South Korea. Environ. Monit. Assess. 2022, 194, 889. [Google Scholar] [CrossRef]
- Bertelsmeier, C. Globalization and the anthropogenic spread of invasive social insects. Curr. Opin. Insect Sci. 2021, 46, 16–23. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L. Analysis of invasive insects: Links to climate change. In Invasive Species and Global Climate Change; CABI: Wallingford, UK, 2022; pp. 50–73. [Google Scholar]
- Ma, G.; Ma, C.S. Potential distribution of invasive crop pests under climate change: Incorporating mitigation responses of insects into prediction models. Curr. Opin. Insect Sci. 2022, 49, 15–21. [Google Scholar] [CrossRef]
- Tobin, P.C.; Robinet, C. Advances in understanding and predicting the spread of invading insect populations. Curr. Opin. Insect Sci. 2022, 54, 100985. [Google Scholar] [CrossRef]
- Hudgins, E.J.; Liebhold, A.M.; Leung, B. Predicting the spread of all invasive forest pests in the United States. Ecol. Lett. 2017, 20, 426–435. [Google Scholar] [CrossRef]
- Deng, J.; Li, J.; Zhang, X.; Zeng, L.; Guo, Y.; Wang, X.; Chen, Z.J.; Zhou, J.L.; Huang, X.L. Potential Global Invasion Risk of Scale Insects Based on a Self-Organizing Map. Insects 2023, 14, 572. [Google Scholar] [CrossRef]
- Pedrosa, M.C.; Borges, M.A.Z.; Eiras, Á.E.; Caldas, S.; Cecílio, A.B.; Brito, M.F.; Ribeiro, S.P. Invasion of tropical montane cities by Aedes aegypti and Aedes albopictus (Diptera: Culicidae) depends on continuous warm winters and suitable urban biotopes. J. Med. Entomol. 2021, 58, 333–342. [Google Scholar] [CrossRef]
- Justine, J.L.; Winsor, L.; Gey, D.; Gros, P.; Thévenot, J. The invasive New Guinea flatworm Platydemus manokwari in France, the first record for Europe: Time for action is now. PeerJ 2014, 2, e297. [Google Scholar] [CrossRef]
- Nunez-Mir, G.C.; Walter, J.A.; Grayson, K.L.; Johnson, D.M. Assessing drivers of localized invasive spread to inform large-scale management of a highly damaging insect pest. Ecol. Appl. 2022, 32, e2538. [Google Scholar] [CrossRef]
- Mogi, M.; Armbruster, P.A.; Tuno, N.; Aranda, C.; Yong, H.S. The climate range expansion of Aedes albopictus (Diptera: Culicidae) in Asia inferred from the distribution of Albopictus subgroup species of Aedes (Stegomyia). J. Med. Entomol. 2017, 54, 1615–1625. [Google Scholar] [CrossRef]
- Adler, C.; Athanassiou, C.; Carvalho, M.O.; Emekci, M.; Gvozdenac, S.; Hamel, D.; Riudavets, J.; Stejskal, V.; Trdan, S.; Trematerra, P. Changes in the distribution and pest risk of stored product insects in Europe due to global warming: Need for pan-European pest monitoring and improved food-safety. J. Stored Prod. Res. 2022, 97, 101977. [Google Scholar] [CrossRef]
- Netherer, S.; Schopf, A. Potential effects of climate change on insect herbivores in European forests—General aspects and the pine processionary moth as specific example. For. Ecol. Manag. 2010, 259, 831–838. [Google Scholar] [CrossRef]
- Battisti, A.; Larsson, S.; Roques, A. Processionary moths and associated urtication risk: Global change-driven effects. Annu. Rev. Entomol. 2017, 62, 323–342. [Google Scholar] [CrossRef]
- Olabimi, I.O.; Ileke, K.D.; Adu, B.W.; Arotolu, T.E. Potential distribution of the primary malaria vector Anopheles gambiae Giles [Diptera: Culicidae] in Southwest Nigeria under current and future climatic conditions. J. Basic Appl. Zool. 2021, 82, 63. [Google Scholar] [CrossRef]
- LeBrun, E.G.; Plowes, R.M.; Gilbert, L.E. Imported fire ants near the edge of their range: Disturbance and moisture determine prevalence and impact of an invasive social insect. J. Anim. Ecol. 2012, 81, 884–895. [Google Scholar] [CrossRef]
- Simões, M.V.; Nuñez-Penichet, C.; Warren, D.; Schmitt, T.; Krull, M. Regional anthropogenic disturbance and species-specific niche traits influence the invasiveness of European beetle species. Front. Ecol. Evol. 2023, 11, 1160598. [Google Scholar] [CrossRef]
- Munga, S.; Yakob, L.; Mushinzimana, E.; Zhou, G.; Ouna, T.; Minakawa, N.; Githeko, A.; Yan, G.Y. Land use and land cover changes and spatiotemporal dynamics of anopheline larval habitats during a four-year period in a highland community of Africa. Am. J. Trop. Med. Hyg. 2009, 81, 1079. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.Y.; Gao, X.; Ma, J.; Jiao, Z.H.; Xiao, J.H.; Hayat, M.A.; Wang, H.B. Modeling the present and future distribution of arbovirus vectors Aedes aegypti and Aedes albopictus under climate change scenarios in Mainland China. Sci. Total Environ. 2019, 664, 203–214. [Google Scholar] [CrossRef]
- Liu, T.M.; Wang, J.M.; Hu, X.K.; Feng, J.M. Land-use change drives present and future distributions of Fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Sci. Total Environ. 2020, 706, 135872. [Google Scholar] [CrossRef]
- Ageep, T.B.; Cox, J.; Hassan, M.O.M.; Knols, B.G.; Benedict, M.Q.; Malcolm, C.A. Spatial and temporal distribution of the malaria mosquito Anopheles arabiensis in northern Sudan: Influence of environmental factors and implications for vector control. Malaria J. 2009, 8, 123. [Google Scholar] [CrossRef]
- Nie, P.X.; Yang, R.J.; Cao, R.Y.; Hu, X.K.; Feng, J.M. Niche and Range Shifts of the Fall Webworm (Hyphantria cunea Dury) in Europe Imply Its Huge Invasion Potential in the Future. Insects 2023, 14, 316. [Google Scholar] [CrossRef] [PubMed]
- Dudov, S.V. Modeling of species distribution with the use of topography and remote sensing data on the example of vascular plants of the Tukuringra Ridge Low Mountain Belt (Zeya State Nature Reserve, Amur Oblast). Biol. Bull. Rev. 2017, 7, 246–257. [Google Scholar] [CrossRef]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.H.; Yang, Y.C.; Ohsawa, M.; Luu, H.T.; et al. Identifying long-term stable refugia for relict plant species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef]
- Virkkala, R.; Marmion, M.; Heikkinen, R.K.; Thuiller, W.; Luoto, M. Predicting range shifts of northern bird species: Influence of modelling technique and topography. Acta Oecol. 2010, 36, 269–281. [Google Scholar] [CrossRef]
- Azrag, A.G.; Pirk, C.W.; Yusuf, A.A.; Pinard, F.; Niassy, S.; Mosomtai, G.; Babin, R. Prediction of insect pest distribution as influenced by elevation: Combining field observations and temperature-dependent development models for the coffee stink bug, Antestiopsis thunbergii (Gmelin). PLoS ONE 2018, 13, e0199569. [Google Scholar] [CrossRef]
- Adeogun, A.; Babalola, A.S.; Okoko, O.O.; Oyeniyi, T.; Omotayo, A.; Izekor, R.T. Spatial distribution and ecological niche modeling of geographical spread of Anopheles gambiae complex in Nigeria using real time data. Sci. Rep. 2023, 13, 13679. [Google Scholar] [CrossRef] [PubMed]
- Macedo, F.L.; Ragonezi, C.; Reis, F.; de Freitas, J.G.; Lopes, D.H.; Aguiar, A.M.F.; Cravo, D.; de Carvalho, M.A.A.P. Prediction of the Potential Distribution of Drosophila suzukii on Madeira Island Using the Maximum Entropy Modeling. Agriculture 2023, 13, 1764. [Google Scholar] [CrossRef]
- de Souza, D.; Kelly-Hope, L.; Lawson, B.; Wilson, M.; Boakye, D. Environmental Factors Associated with the Distribution of Anopheles gambiae s.s in Ghana; an Important Vector of Lymphatic Filariasis and Malaria. PLoS ONE 2020, 5, e9927. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Fu, J.; Jiang, D.; Hao, M.; Lin, G. Maping the spatial distribution of Aedes aegypti and Aedes albopictus. Acta Trop. 2018, 178, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Echeverry-Cárdenas, E.; López-Castañeda, C.; Carvajal-Castro, J.D.; Aguirre-Obando, O.A. Potential geographic distribution of the tiger mosquito Aedes albopictus (Skuse, 1894) (Diptera: Culicidae) in current and future conditions for Colombia. PloS Neglect. Trop. D 2021, 15, e0008212. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Williams, R.; Chen, G. Modeled global invasive potential of Asian gypsy moths, Lymantria dispar. Entomol Exp. Appl. 2007, 125, 39–44. [Google Scholar] [CrossRef]
- Jung, J.M.; Kim, S.H.; Jung, S.; Lee, W.H. Spatial and climatic analyses for predicting potential distribution of an invasive ant, Linepithema humile (Hymenoptera: Formicidae). Entomol. Sci. 2022, 25, e12527.39. [Google Scholar] [CrossRef]
- Nie, P.; Feng, J. Niche and range shifts of Aedes aegypti and Ae. albopictus suggest that the latecomer shows a greater invasiveness. Insects 2023, 14, 810. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Wu, C.H.; Nie, P.X.; Feng, J.M.; Hu, X.K. Invasive Pest and Invasive Host: Where Might Spotted-Wing Drosophila (Drosophila suzukii) and American Black Cherry (Prunus serotina) Cross Paths in Europe? Forests 2024, 15, 206. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- O’Neill, B.C.; Tebaldi, C.; Van Vuuren, D.P.; Eyring, V.; Friedlingstein, P.; Hurtt, G.; Knutti, R.; Kriegler, E.; Lamarque, J.-F.; Lowe, J.; et al. The Scenario Model Intercomparison Project (ScenarioMIP) for CMIP6. Geosci. Model Dev. 2016, 9, 3461–3482. [Google Scholar] [CrossRef]
- Riahi, K.; Van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O.; et al. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Change 2017, 42, 153–168. [Google Scholar] [CrossRef]
- 4Zhang, M.Z.; Xu, Z.F.; Han, Y.; Guo, W.D. Evaluation of CMIP6 models toward dynamical downscaling over 14 CORDEX domains. Clim. Dyn. 2022, 58, 1–14. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araujo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G. White M, On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.J.; Yu, X.L.; Nie, P.X.; Cao, R.Y.; Feng, J.M.; Hu, X.K. Climatic niche and range shifts of grey squirrels (Sciurus carolinensis Gmelin) in Europe: An invasive pest displacing native squirrels. Pest Manag. Sci. 2023, 79, 3731–3739. [Google Scholar] [CrossRef] [PubMed]
- Cao, R.Y.; Feng, J.M. Future Range Shifts Suggest That the Six-Spined Spruce Bark Beetle Might Pose a Greater Threat to Norway Spruce in Europe than the Eight-Spined Spruce Bark Beetle. Forests 2023, 14, 2048. [Google Scholar] [CrossRef]
- Sirami, C.; Caplat, P.; Popy, S.; Clamens, A.; Arlettaz, R.; Jiguet, F. Impacts of global change on species distributions: Obstacles and solutions to integrate climate and land use. Glob. Ecol. Biogeogr. 2017, 26, 385–394. [Google Scholar] [CrossRef]
- Levine, R.S.; Peterson, A.T.; Benedict, M.Q. Distribution of members of Anopheles quadrimaculatus Say sl (Diptera: Culicidae) and implications for their roles in malaria transmission in the United States. J. Med. Entomol. 2004, 41, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Ward, P.S. Phylogeny, classification, and species-level taxonomy of ants (Hymenoptera: Formicidae). Zootaxa 2007, 1668, 549–563. [Google Scholar] [CrossRef]
- Zhao, Z.; Hui, C.; Peng, S.; Yi, S.; Li, Z.; Reddy, G.V.; van Kleunen, M. The world’s 100 worst invasive alien insect species differ in their characteristics from related non-invasive species. J. Appl. Ecol. 2023, 60, 1929–1938. [Google Scholar] [CrossRef]
- Fält-Nardmann, J.J.; Ruohomäki, K.; Tikkanen, O.P.; Neuvonen, S. Cold hardiness of Lymantria monacha and L. dispar (Lepidoptera: Erebidae) eggs to extreme winter temperatures: Implications for predicting climate change impacts. Ecol. Entomol. 2018, 43, 422–430. [Google Scholar] [CrossRef]
- Tanga, M.C.; Ngundu, W.I.; Judith, N.; Mbuh, J.; Tendongfor, N.; Simard, F.; Wanji, S. Climate change and altitudinal structuring of malaria vectors in south-western Cameroon: Their relation to malaria transmission. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Soleimani-Ahmadi, M.; Vatandoost, H.; Zare, M.; Turki, H.; Alizadeh, A. Topographical distribution of anopheline mosquitoes in an area under elimination programme in the south of Iran. Malaria J. 2015, 14, 262. [Google Scholar] [CrossRef] [PubMed]
- Mwakalinga, V.M.; Sartorius, B.K.; Limwagu, A.J.; Mlacha, Y.P.; Msellemu, D.F.; Chaki, P.P.; Govella, N.J.; Coetzee, M.; Dongus, S.; Killeen, G.F. Topographic maping of the interfaces between human and aquatic mosquito habitats to enable barrier targeting of interventions against malaria vectors. R. Soc. Open Sci. 2018, 5, 161055. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Xian, W.W.; Jeschke, J.M. Most invasive species largely conserve their climatic niche. Proc. Natl. Acad. Sci. USA 2020, 117, 23643–23651. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Names | Scientific Names | Family | Native Regions | Major Impacts |
|---|---|---|---|---|
| Argentine ant | Linepithema humile | Formicidae | South America | Biodiversity losses |
| Asian gypsy moth | Lymantria dispar | Erebidae | Asia and Europe | Forest losses |
| Asian longhorned beetle | Anoplophora glabripennis | Cerambycidae | Asia | Forest losses |
| Asian tiger mosquito | Aedes albopictus | Culicidae | Southeast Asia | Transmission of diseases |
| Big-headed ant | Pheidole megacephala | Formicidae | Southern Africa | Biodiversity and crop loss |
| Cocoa tree-ant | Wasmannia auropunctata | Formicidae | South America | Biodiversity losses |
| Common malaria mosquito | Anopheles quadrimaculatus | Culicidae | North America | Transmission of diseases |
| Common wasp | Vespula vulgaris | Vespidae | Europe | Human health and insect losses |
| Cotton whitefly | Bemisia tabaci | Aleyrodidae | India | Crop losses |
| Crazy ant | Anoplolepis gracilipes | Formicidae | Unclear | Biodiversity losses |
| Cypress aphid | Cinara cupressi | Aphididae | North America | Forest losses |
| Flatworm | Platydemus manokwari | Rhynchodemidae | New Guinea, Africa | Biodiversity losses |
| Formosa termite | Coptotermes formosanus | Rhinotermitidae | China | Economic losses |
| Khapra beetle | Trogoderma granarium | Dermestidae | Indian subcontinent | Food storage |
| Red imported fire ant | Solenopsis invicta | Formicidae | South America | Biodiversity losses |
| Potential Ranges (million km2) | Range Ratio Index | Range Similarity Index | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | CurPR | F126PR | F585PR | M126PR | M585PR | F126RRI | F585RRI | M126RRI | M585RRI | F126RSI | F585RSI | M126RSI | M585RSI |
| Linepithema humile | 7.16 | 8.76 | 10.92 | 8.87 | 10.41 | 1.19 | 1.47 | 1.20 | 1.40 | 0.79 | 0.63 | 0.78 | 0.66 |
| Anoplophora glabripennis | 4.23 | 9.74 | 16.11 | 11.20 | 12.00 | 2.32 | 3.83 | 2.66 | 2.85 | 0.49 | 0.36 | 0.43 | 0.47 |
| Aedes albopictus | 11.66 | 22.00 | 27.11 | 21.74 | 26.63 | 1.84 | 2.26 | 1.82 | 2.23 | 0.66 | 0.55 | 0.67 | 0.57 |
| Pheidole megacephala | 8.23 | 13.46 | 20.51 | 15.78 | 18.09 | 1.58 | 2.39 | 1.86 | 2.12 | 0.71 | 0.52 | 0.64 | 0.54 |
| Anopheles quadrimaculatus | 3.98 | 6.72 | 7.20 | 5.85 | 6.53 | 1.64 | 1.77 | 1.42 | 1.60 | 0.65 | 0.64 | 0.66 | 0.67 |
| Vespula vulgaris | 7.09 | 10.46 | 11.42 | 8.24 | 9.93 | 1.41 | 1.51 | 1.11 | 1.33 | 0.78 | 0.62 | 0.86 | 0.71 |
| Anoplolepis gracilipes | 5.84 | 8.12 | 9.57 | 9.01 | 9.14 | 1.41 | 1.67 | 1.57 | 1.59 | 0.75 | 0.69 | 0.73 | 0.69 |
| Cinara cupressi | 17.25 | 19.83 | 34.63 | 24.19 | 30.44 | 1.13 | 1.98 | 1.38 | 1.74 | 0.80 | 0.53 | 0.66 | 0.61 |
| Platydemus manokwari | 3.86 | 3.56 | 8.33 | 4.52 | 3.62 | 0.95 | 2.22 | 1.20 | 0.96 | 0.60 | 0.48 | 0.58 | 0.50 |
| Coptotermes formosanus | 1.19 | 2.14 | 6.47 | 1.45 | 3.59 | 1.72 | 5.25 | 1.17 | 2.92 | 0.56 | 0.29 | 0.58 | 0.39 |
| Lymantria dispar | 8.76 | 11.81 | 21.23 | 11.26 | 18.77 | 1.36 | 2.44 | 1.29 | 2.16 | 0.81 | 0.52 | 0.83 | 0.58 |
| Trogoderma granarium | 12.91 | 18.20 | 23.22 | 25.21 | 41.04 | 1.39 | 1.76 | 1.93 | 3.13 | 0.71 | 0.54 | 0.65 | 0.47 |
| Wasmannia auropunctata | 8.86 | 12.00 | 18.58 | 13.68 | 18.11 | 1.32 | 2.05 | 1.51 | 2.00 | 0.74 | 0.58 | 0.72 | 0.60 |
| Solenopsis invicta | 4.78 | 8.15 | 10.17 | 7.63 | 7.40 | 1.66 | 2.07 | 1.55 | 1.50 | 0.60 | 0.47 | 0.57 | 0.52 |
| Bemisia tabaci | 10.45 | 11.90 | 12.57 | 17.41 | 14.29 | 1.14 | 1.21 | 1.67 | 1.37 | 0.78 | 0.75 | 0.71 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, R.; Feng, J. Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests. Insects 2024, 15, 280. https://doi.org/10.3390/insects15040280
Cao R, Feng J. Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests. Insects. 2024; 15(4):280. https://doi.org/10.3390/insects15040280
Chicago/Turabian StyleCao, Runyao, and Jianmeng Feng. 2024. "Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests" Insects 15, no. 4: 280. https://doi.org/10.3390/insects15040280
APA StyleCao, R., & Feng, J. (2024). Future Climate Change and Anthropogenic Disturbance Promote the Invasions of the World’s Worst Invasive Insect Pests. Insects, 15(4), 280. https://doi.org/10.3390/insects15040280