
Bombus terrestris Prefer Mixed-Pollen Diets for a Better Colony Performance: A Laboratory Study

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pollens and Bumblebees
2.2. Different Pollen Diets Used to Feed Bumblebees
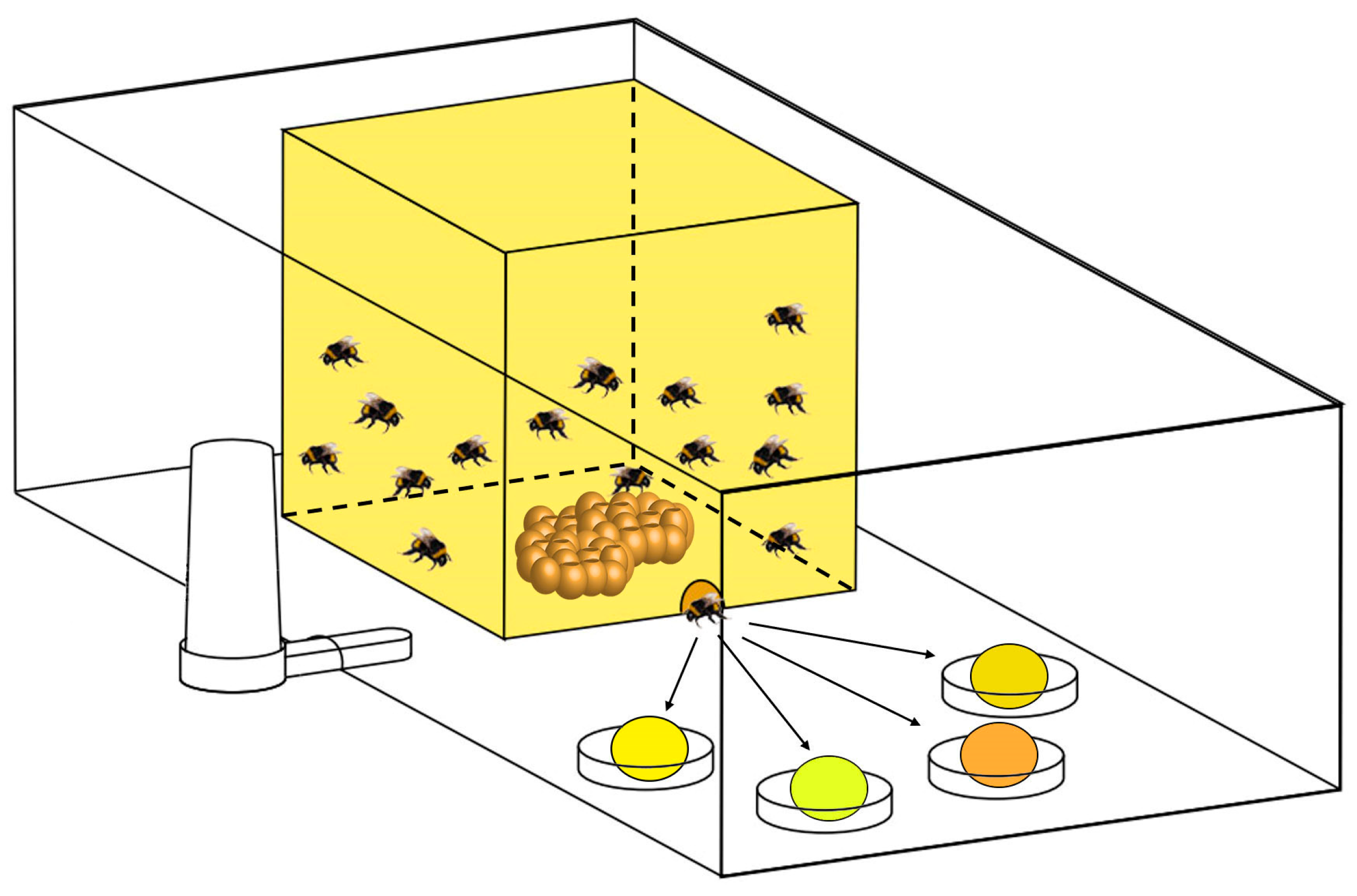
2.2.1. Testing Food Preference during Colony Development of Bumblebees with Four Different Pollen Varieties under Laboratory Conditions
2.2.2. Monitoring Bumblebee Colony Development When Feeding a Single Pollen Type under Laboratory Conditions
2.3. Determination of Pollen Proteins and Lipids
2.3.1. Determination of Protein Content of the Four Pollens
2.3.2. Determination of Lipid Content of the Four Pollens
2.3.3. Protein Consumption, Lipid Consumption, and P:L Ratio
2.4. Data Analysis
3. Results
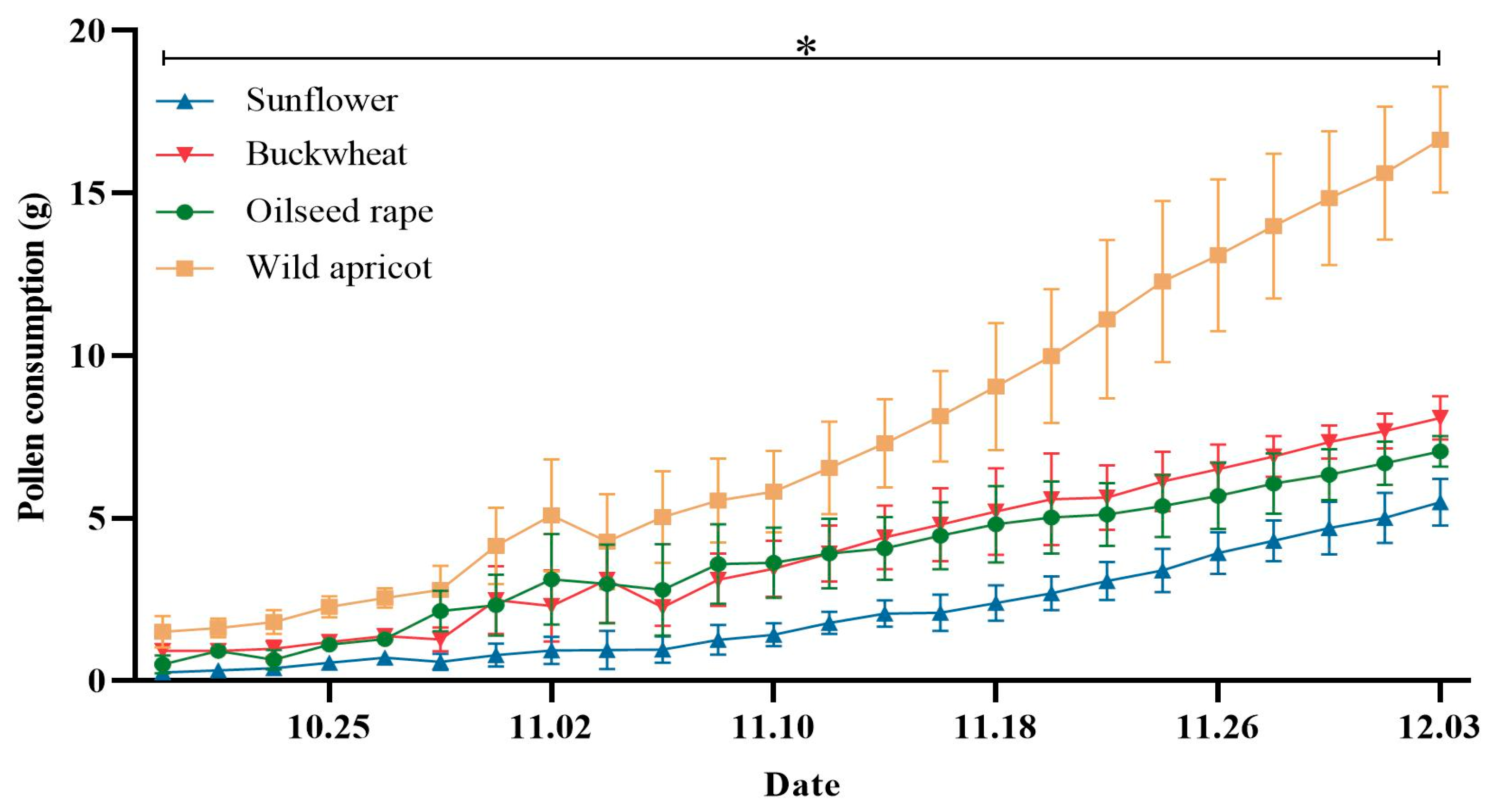
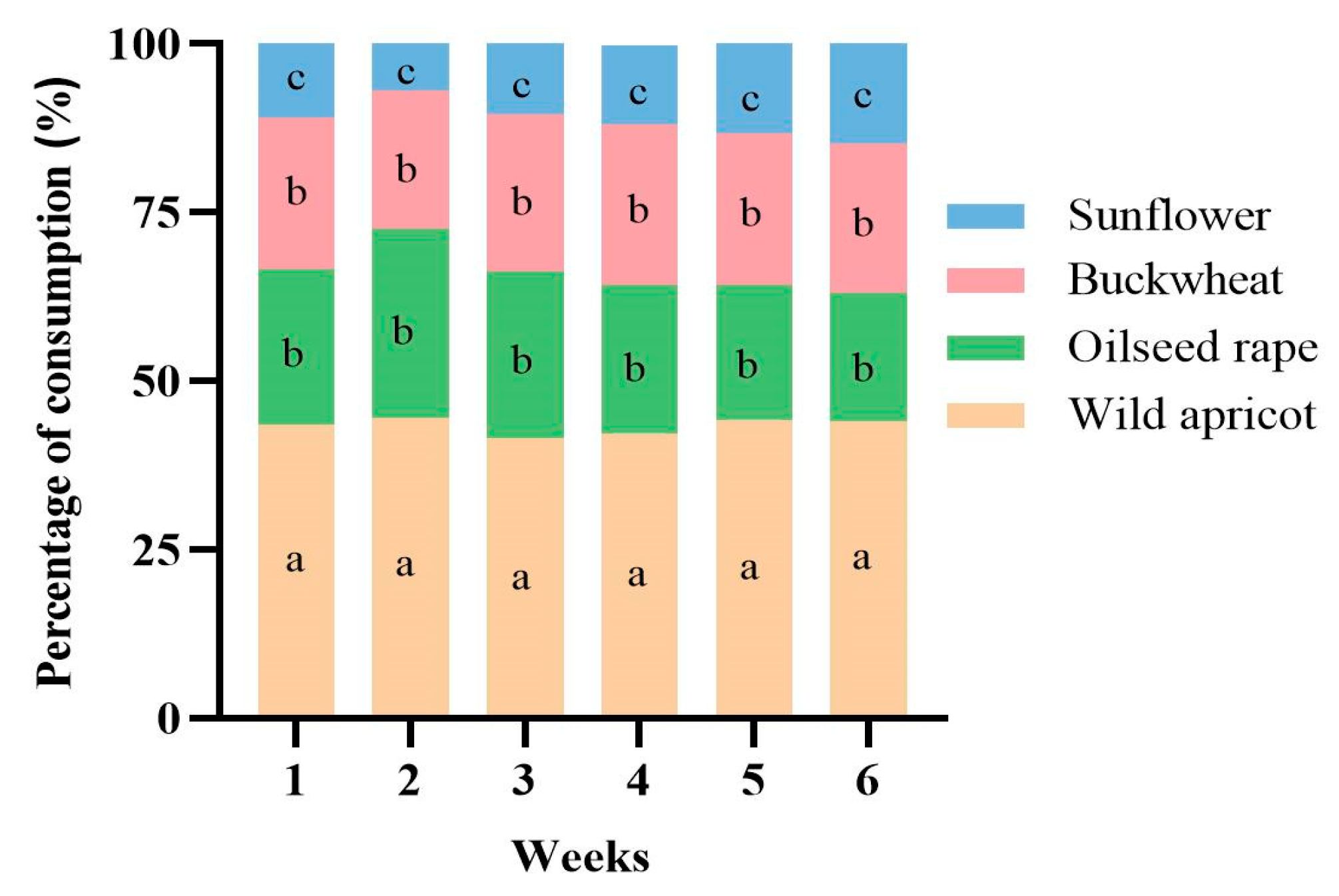
3.1. Bumblebees Prefer Wild Apricot Pollen during Colony Development
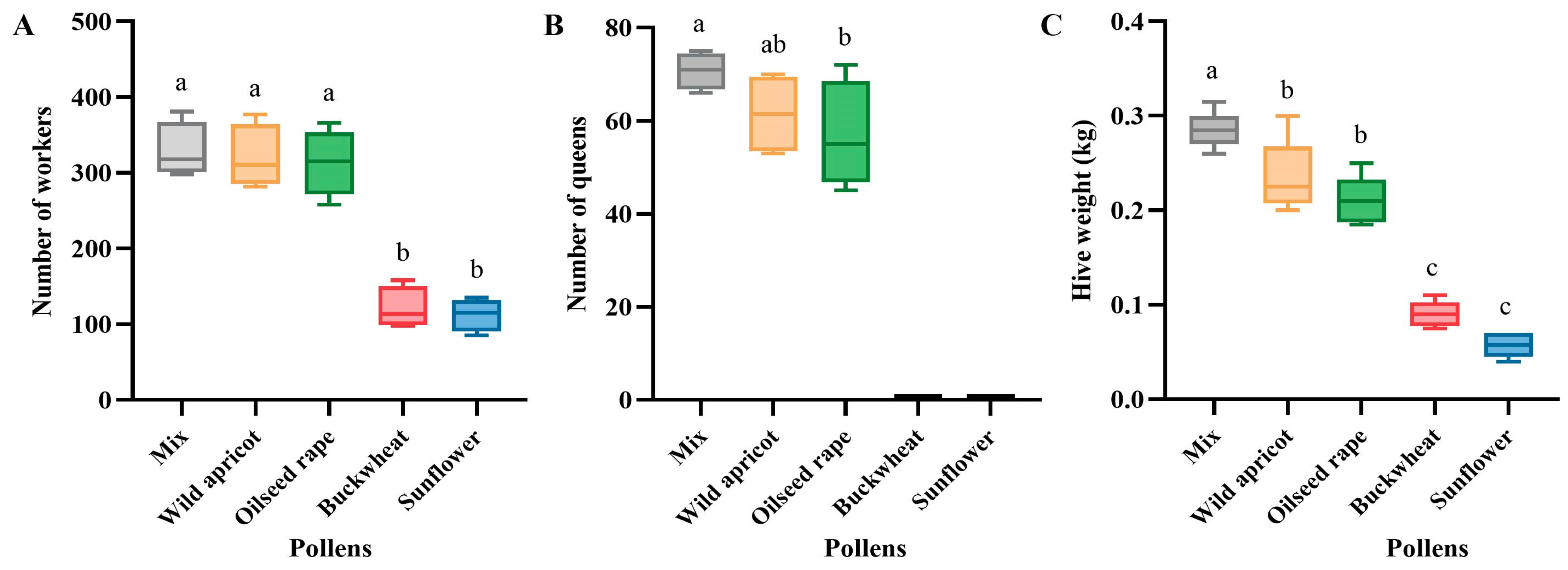
3.2. Effects of Different Pollen Diets on Bumblebee Colony Development
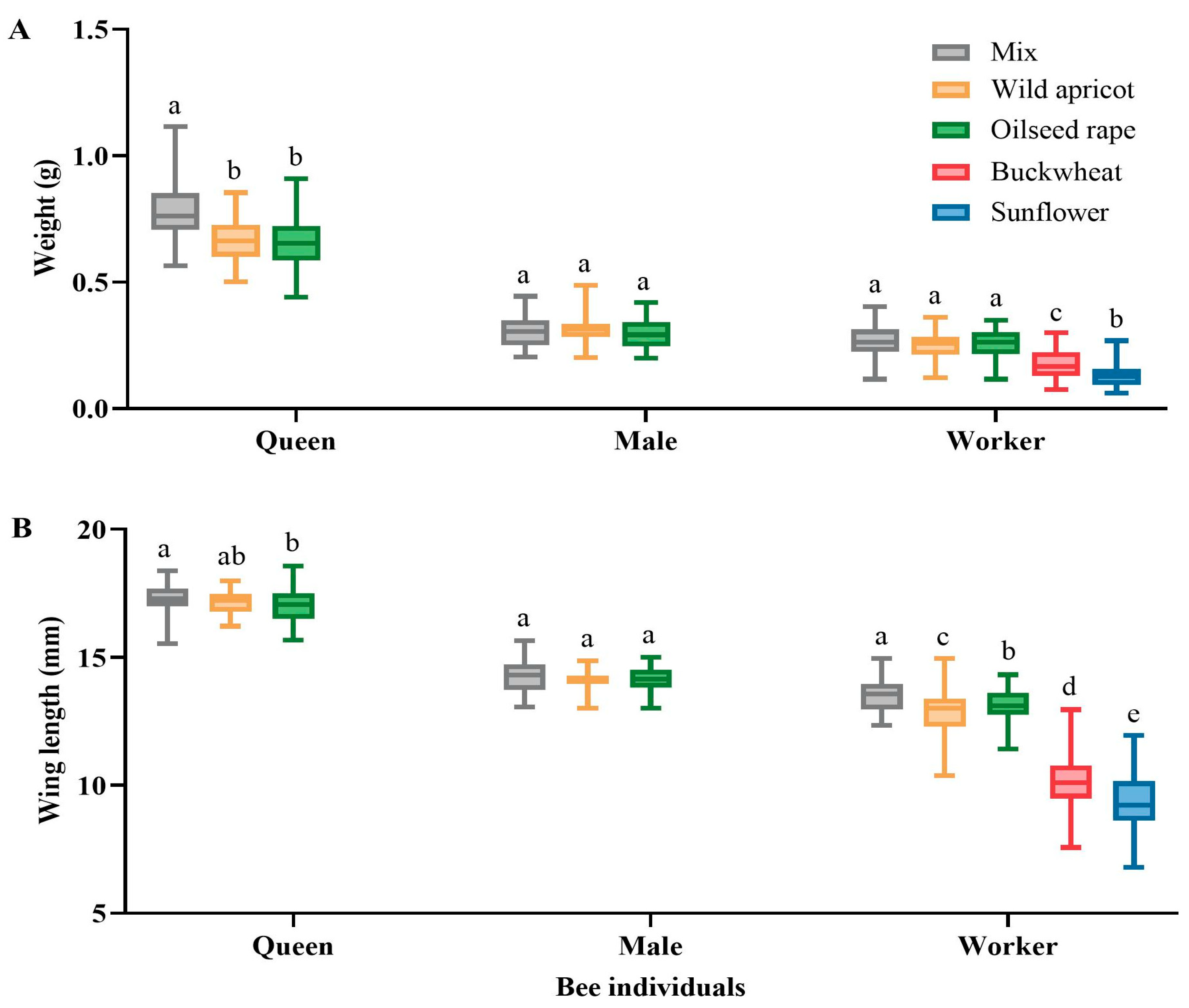
3.2.1. Effects on Bumblebee Individuals
3.2.2. Effects on Bumblebee Colonies
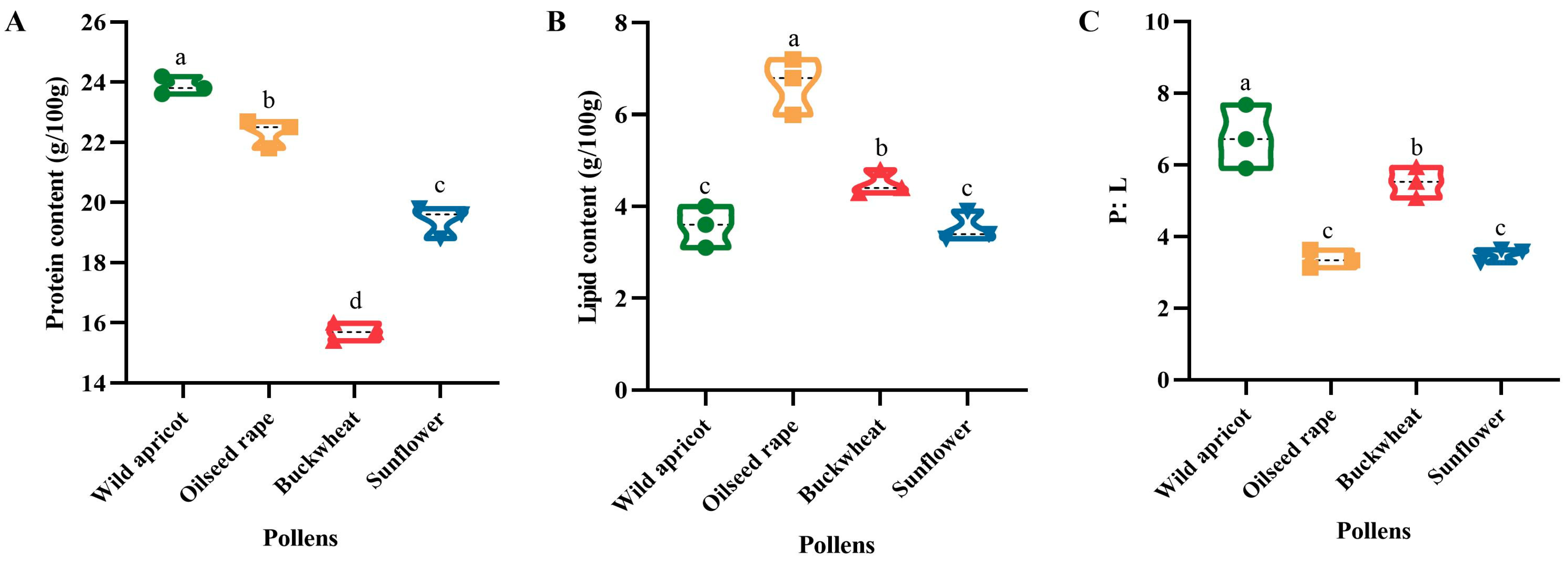
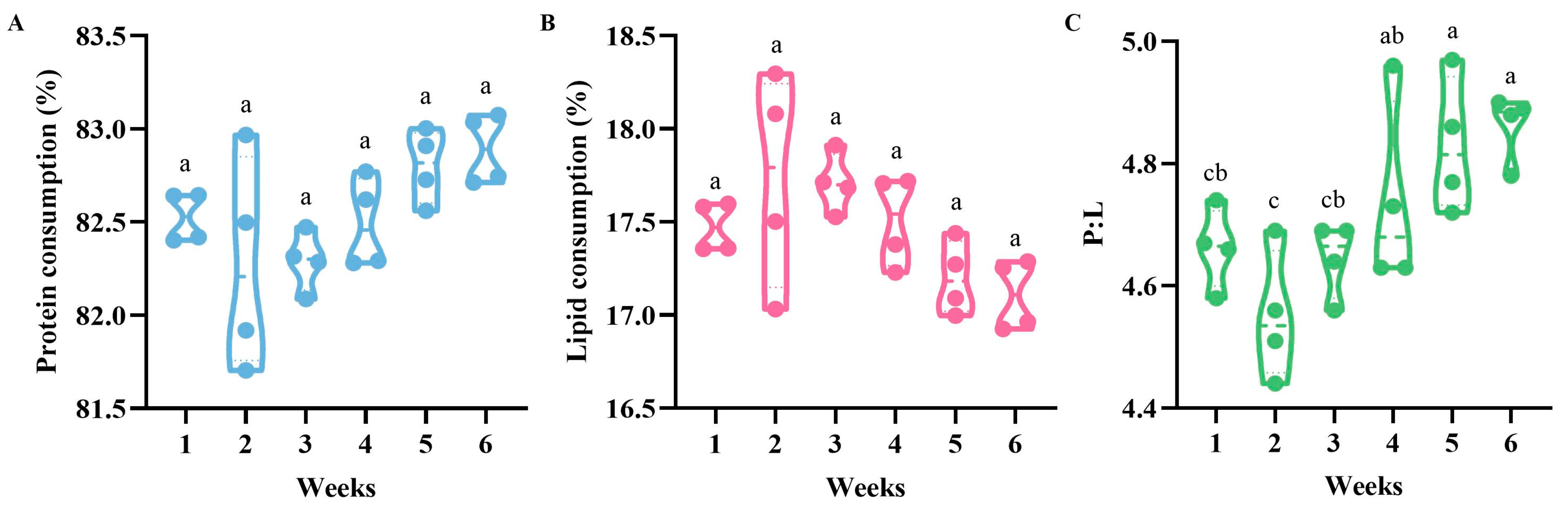
3.3. Protein and Lipid Consumption of Bumblebees during Colony Development
3.3.1. Protein and Lipid Contents and P:L Ratio of the Four Pollens
3.3.2. Consumption of Proteins and Lipids, and P:L Ratios of Bumblebees during Colony Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Burkle, L.A.; Martin, J.C.; Knight, T.M. Plant-pollinator interactions over 120 years: Loss of species, co-occurrence, and function. Science 2013, 339, 1611–1615. [Google Scholar] [CrossRef]
- Cameron, S.A.; Hines, H.M.; Williams, P.H. A comprehensive phylogeny of the bumble bees (Bombus). Biol. J. Linn. Soc. 2007, 91, 161–188. [Google Scholar] [CrossRef]
- Abrahamovich, A.H.; Díaz, N.B. Bumblebees of the neotropical region (Hymenoptera: Apidae). Biota Colomb. 2002, 3, 199–214. [Google Scholar]
- Huang, J.X.; An, J.D. Species diversity, pollination application and strategy for conservation of the bumblebees of China. Biodivers. Sci. 2018, 25, 486–497. [Google Scholar] [CrossRef]
- Iserbyt, S.; Rasmont, P. The effect of climatic variation on abundance and diversity of bumblebees: A ten years survey in a mountain hotspot. Ann. Socit Entomol. Fr. 2012, 48, 261–273. [Google Scholar] [CrossRef]
- Williams, P.H.; Cameron, S.A.; Hines, H.M.; Cederberg, B.; Rasmont, P. A simplified subgeneric classification of the bumblebees (genus Bombus). Apidologie 2008, 39, 46–74. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; van Doorn, A.V. A century of advances in bumblebee domestication and the economic and environ–mental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Zhang, H.; Han, C.; Breeze, T.D.; Li, M.D.; Mashilingi, S.K.; Hua, J.; Zhang, W.B.; Zhang, X.B.; Zhang, S.W.; An, J.D. Bumblebee pollination enhances yield and flavor of tomato in Gobi Desert greenhouses. Agriculture 2022, 12, 794. [Google Scholar] [CrossRef]
- Friend, W.G. Nutritional requirements of phytophagous insects. Annu. Rev. Entomol. 1958, 3, 57–74. [Google Scholar] [CrossRef]
- Qin, J.M.; Su, R.; Liang, C.; Liu, F.; Yang, S.; Zhao, H.M.; Zhao, Z.Y. Research progress of biology and population influencing factors in bumblebee. J. Environ. Entomol. 2020, 42, 1383–1393. [Google Scholar]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation, 1st ed.; Oxford University Press: Oxford, UK, 2010; p. 317. [Google Scholar]
- Carnell, J.D.; Hulse, R.A.; Hughes, W.O.H. Review of nutrition in bumblebees: The effect of caste, life–stage and life history traits. Adv. Insect Physiol. 2020, 59, 71–129. [Google Scholar]
- Génissel, A.; Aupinel, P.; Bressac, C.; Tasei, J.N.; Chevrier, C. Influence of pollen origin on performance of Bombus terrestris micro-colonies. Entomol. Exp. Appl. 2010, 104, 329–336. [Google Scholar] [CrossRef]
- Human, H.; Nicolson, S.W.; Strauss, K.; Pirk, C.W.W.; Dietemann, V. Influence of pollen quality on ovarian development in honeybee workers (Apis mellifera scutellata). J. Insect Physiol. 2007, 53, 649–655. [Google Scholar] [CrossRef]
- Tasei, J.N.; Aupinel, P. Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 2008, 39, 397–409. [Google Scholar] [CrossRef]
- Alaux, C.; Ducloz, F.; Crauser, D.; Conte, Y.L. Diet effects on honeybee immunocompetence. Biol. Lett. 2010, 6, 562–565. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Harris, G.K.; Grozinger, C.M. Effects of soil quality enhancement on pollinator-plant interactions. Psyche A J. Entomol. 2012, 2012, 581458. [Google Scholar] [CrossRef]
- Li, C.C.; Xu, B.H.; Wang, Y.X.; Feng, Q.Q.; Yang, W.R. Effects of dietary crude protein levels on development, antioxidant status, and total midgut protease activity of honey bee (Apis mellifera ligustica). Apidologie 2012, 43, 576–586. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Moerman, R.; Rasmont, P.; Lognay, G.; Wathelet, B.; Wattiez, R.; Michez, D. How does pollen chemistry impact development and feeding behaviour of polylectic bees? PLoS ONE 2014, 9, e0086209. [Google Scholar] [CrossRef]
- Canavoso, L.E.; Jouni, Z.E.; Karnas, K.J.; Pennington, J.E.; Wells, M.A. Fat metabolism in insects. Annu. Rev. Nutr. 2001, 21, 23–46. [Google Scholar] [CrossRef]
- Yael, A.; Arnon, D.; Sharoni, S. Omega-6:3 ratio more than absolute lipid level in diet affects associative learning in honey bees. Front. Psychol. 2018, 9, 1001. [Google Scholar]
- Herbert, E.W.; Svoboda, J.A.; Thompson, M.J.; Shimanuki, H. Sterol utilization in honeybees fed a synthetic diet: Effects on brood rearing. J. Insect Physiol. 1980, 26, 287–289. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Leonhardt, S.D.; Blüthgen, N. The same, but different: Pollen foraging in honeybee and bumblebee colonies. Apidologie 2012, 43, 449–464. [Google Scholar] [CrossRef]
- Kitaoka, T.K.; Nieh, J.C. Manuscript in preparation for behavioral ecology and sociobiology bumblebee pollen foraging regulation: Role of pollen quality, storage levels, and odor. Behav. Ecol. Sociobiol. 2009, 63, 501–510. [Google Scholar] [CrossRef]
- Robertson, A.W.; Mountjoy, C. Bumblebee selection of mimulus guttatus flowers: The effects of pollen quality and reward depletion. Ecology 1999, 80, 2594–2606. [Google Scholar] [CrossRef]
- Hanley, M.E.; Franco, M.; Pichon, S.; Darvill, B.; Goulson, D. Breeding system, pollinator choice and variation in pollen quality in british herbaceous plants. Funct. Ecol. 2008, 22, 592–598. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Stabler, D.; Patch, H.M.; Tooker, J.F.; Grozinger, C.M.; Wright, G.A. Bumblebees regulate their intake of essential protein and lipid pollen macronutrients. J. Exp. Biol. 2016, 219, 3962–3970. [Google Scholar] [PubMed]
- Vaudo, A.D.; Patch, H.M.; Mortensen, D.A.; Tooker, J.F.; Grozinger, C.M. Macronutrient ratios in pollen shape bumblebee (Bombus impatiens) foraging strategies and floral preferences. Proc. Natl. Acad. Sci. USA 2016, 113, E4035–E4042. [Google Scholar]
- Wang, H.; Xu, X.L. Research progress in bumblebee nutrition and feed. Acta Entomol. Sin. 2018, 61, 620–626. [Google Scholar]
- Zhao, H.Y.; Liu, Y.J.; Zhang, H.; Breeze, T.D.; An, J.D. Worker-born males are smaller but have similar reproduction ability to queen-born males in bumblebees. Insects 2021, 12, 1008. [Google Scholar] [CrossRef] [PubMed]
- He, C.L.; Xu, S.S.; Zhang, S.X.; Hou, X.G.; Zhu, C.D. Analysis of the protein and mineral elements content in pollen of nine Paeonia suffrutisosa cultivars. J. Nucl. Agric. Sci. 2015, 29, 2158–2164. [Google Scholar]
- Yang, Y.T. Fatty Acid Profiles in 20 Different Species of Bee Pollens. Master’s Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2014. [Google Scholar]
- Huang, J.X.; An, J.D.; Wu, J.; William, P.H. Extreme food-plant specialisation in megabombus bumblebees as a product of long tongues combined with short nesting seasons. PLoS ONE 2015, 10, e0132358. [Google Scholar] [CrossRef] [PubMed]
- Moerman, R.; Vanderplanck, M.; Fournier, D.; Jacquemart, A.L.; Michez, D. Pollen nutrients better explain bumblebee colony development than pollen diversity. Insect Conserv. Divers. 2017, 10, 171–179. [Google Scholar] [CrossRef]
- Moerman, R.; Roger, N.; Jonghe, R.D.; Michez, D.; Vanderplanck, M. Interspecific variation in bumblebee performance on pollen diet: New insights for mitigation strategies. PLoS ONE 2016, 11, e0168462. [Google Scholar] [CrossRef] [PubMed]
- Teulier, L.; Weber, J.M.; Crevier, J.; Darveau, C.A. Proline as a fuel for insect flight: Enhancing carbohydrate oxidation in hymenopterans. Proc. R. Soc. B-Biol. Sci. 2016, 283, 1834. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.; Shafir, S.; Yehonatan, L.; Palmer, R.G.; Thornburg, R. A novel role for proline in plant floral nectars. Naturwissenschaften 2006, 93, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Awmack, C.S.; Murray, D.A.; Williams, I.H. Are honeybees’ foraging preferences affected by pollen amino acid composition? Ecol. Entomol. 2003, 28, 622–627. [Google Scholar] [CrossRef]
- Somme, L.; Vanderplanck, M.; Michez, D.; Lombaerde, I.; Moerman, R.; Wathelet, B. Pollen and nectar quality drive the major and minor floral choices of bumblebees. Apidologie 2015, 46, 92–106. [Google Scholar] [CrossRef]
- Piskorski, R.; Kroder, S.; Dorn, S. Can pollen headspace volatiles and pollenkitt lipids serve as reliable chemical cues for bee pollinators? Chem. Biodivers. 2011, 8, 577–586. [Google Scholar] [CrossRef]
- Liu, F.L.; Zhang, X.W.; Chai, J.P.; Yang, D.R. Pollen phenolics and regulation of pollen foraging in honeybee colony. Behav. Ecol. Sociobiol. 2006, 59, 582–588. [Google Scholar] [CrossRef]
- Ghosh, S.; Jeon, H.; Jung, C.L. Foraging behaviour and preference of pollen sources by honey bee (Apis mellifera) relative to protein contents. J. Ecol. Environ. 2020, 44, 4. [Google Scholar] [CrossRef]
- Tasei, J.N.; Aupinel, P. Validation of a method using queenless Bombus terrestris micro-colonies for testing the nutritive value of commercial pollen mixes by comparison with queenright colonies. J. Econ. Entomol. 2008, 101, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, S.W.; Human, H. Chemical composition of the ‘low quality’ pollen of sunflower (Helianthus annuus, Asteraceae). Apidologie 2013, 44, 144–152. [Google Scholar] [CrossRef]
- Yang, K.; Wu, D.; Ye, X.Q.; Liu, D.H.; Chen, J.C.; Sun, P.L. Characterization of chemical composition of bee pollen in China. J. Agric. Food Chem. 2013, 61, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Palmer-Young, E.C.; Farrell, I.W.; Adler, L.S.; Milano, N.J.; Egan, P.A.; Junker, R.R.; Irwin, R.E.; Stevenson, P.C. Chemistry of floral rewards: Intra- and interspecific variability of nectar and pollen secondary metabolites across taxa. Ecol. Monogr. 2019, 89, e01335. [Google Scholar] [CrossRef]
- Blackmore, S.; Wortley, A.H.; Skvarla, J.J.; Rowley, J.R. Pollen wall development in flowering plants. New Phytol. 2007, 174, 483–498. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Spatial Arrangement (from Left to Right) | First | Second | Third | Forth |
|---|---|---|---|---|
| Colonies 1–3 | Oilseed rape | Wild apricot | Sunflower | Buckwheat |
| Colonies 4–6 | Buckwheat | Sunflower | Wild apricot | Oilseed rape |
| Colonies 7–8 | Wild apricot | Oilseed rape | Buckwheat | Sunflower |
| Colonies 9–10 | Sunflower | Buckwheat | Oilseed rape | Wild apricot |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Z.; Zhang, H.; Mashilingi, S.K.; Jie, C.; Guo, B.; Guo, Y.; Hu, X.; Iqbal, S.; Wei, B.; Liu, Y.; et al. Bombus terrestris Prefer Mixed-Pollen Diets for a Better Colony Performance: A Laboratory Study. Insects 2024, 15, 285. https://doi.org/10.3390/insects15040285
Zhou Z, Zhang H, Mashilingi SK, Jie C, Guo B, Guo Y, Hu X, Iqbal S, Wei B, Liu Y, et al. Bombus terrestris Prefer Mixed-Pollen Diets for a Better Colony Performance: A Laboratory Study. Insects. 2024; 15(4):285. https://doi.org/10.3390/insects15040285
Chicago/Turabian StyleZhou, Ziyu, Hong Zhang, Shibonage K. Mashilingi, Chunting Jie, Baodi Guo, Yi Guo, Xiao Hu, Shahid Iqbal, Bingshuai Wei, Yanjie Liu, and et al. 2024. "Bombus terrestris Prefer Mixed-Pollen Diets for a Better Colony Performance: A Laboratory Study" Insects 15, no. 4: 285. https://doi.org/10.3390/insects15040285
APA StyleZhou, Z., Zhang, H., Mashilingi, S. K., Jie, C., Guo, B., Guo, Y., Hu, X., Iqbal, S., Wei, B., Liu, Y., & An, J. (2024). Bombus terrestris Prefer Mixed-Pollen Diets for a Better Colony Performance: A Laboratory Study. Insects, 15(4), 285. https://doi.org/10.3390/insects15040285