
More Than 200 Years Later: Gluvia brunnea sp. nov. (Solifugae, Daesiidae), a Second Species of Camel Spider from the Iberian Peninsula


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
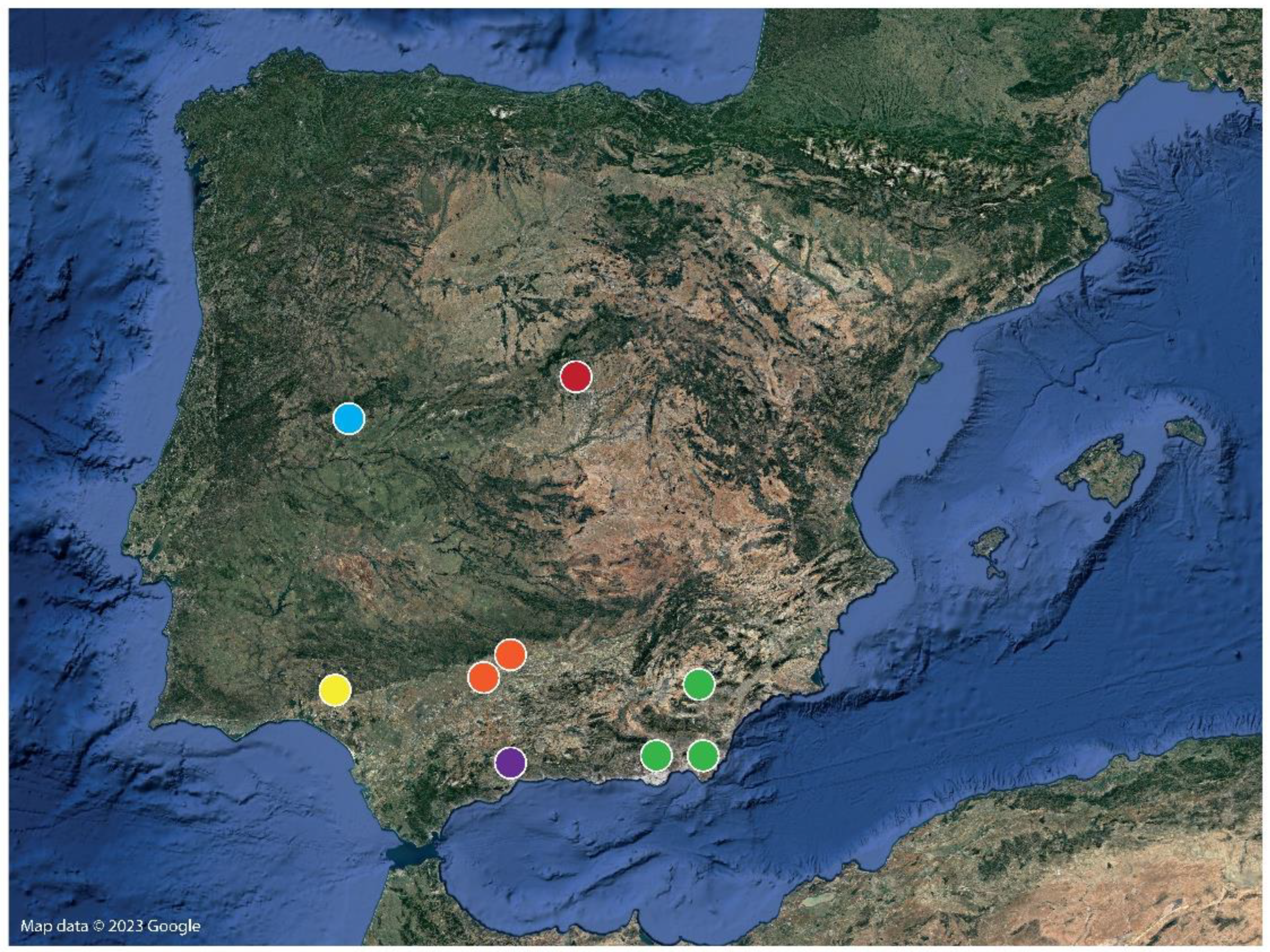
2.1. Sampling Methods
2.2. Scanning Electron Microscopy
2.3. Morphological Characterization
2.4. Statistical Analyses
2.5. DNA Analysis
3. Results
3.1. Taxonomy
3.2. Etymology
3.3. Diagnosis
3.4. Description Holotype (Male)
3.5. Description (Female)
3.6. Statistical Analyses
3.7. Molecular Analyses
4. Discussion
4.1. Intraspecific Variation in Morphology and Recent History of the Genus
4.2. Specimens of Solifugae Captured in Mesovoid Shallow Substratum Stations
4.3. Multiple Factor Analysis Followed by MANOVA, a Successful Procedure to Separate New Species
4.4. Molecular DNA Evidence
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bird, T.L. Cheliceral morphology of Solifugae (Arachnida): Primary Homology, Terminology, and Character Survey. Ph.D. Thesis, Colorado State University, Fort Collins, CO, USA, 2015. [Google Scholar]
- Cushing, P.E.; Graham, M.R.; Prendini, L.; Brookhart, J.O. A multilocus molecular phylogeny of the endemic North American camel spider family Eremobatidae (Arachnida: Solifugae). Mol. Phylogenet. Evol. 2015, 92, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Steiner, H.G.; Garcia, E.L.; Iuri, H.; Jones, R.R.; Ballesteros, J.A.; Gainett, G.; Graham, M.R.; Harms, D.; Lyle, R.; et al. Neglected no longer: Phylogenomic resolution of higher-level relationships in Solifugae. iScience 2023, 26, 107684. [Google Scholar] [CrossRef]
- Hebets, E.A.; Oviedo-Diego, M.; Cargnelutti, F.; Bollatti, F.; Calbacho-Rosa, L.; Mattoni, C.I.; Olivero, P.; Simian, C.; Abregú, D.; Vrech, D.E.; et al. A scientist’s guide to Solifugae: How solifuges could advance research in ecology, evolution, and behaviour. Zool. J. Linn. Soc. 2023, zlad174. [Google Scholar] [CrossRef]
- Roewer, C.F. Part IV. Solifuga, Palpigrada. In Klassen und Ordnungen des Tierreichs; Bronns, H.G., Ed.; Akadem. Verlagsgesell.: Leipzig, Germany, 1934; pp. 481–608. [Google Scholar]
- Wharton, R.A. Namibian (South Africa) Solifugae. Cim. Mem. 1981, 5, 3–87. [Google Scholar]
- Kulkarni, S.S.; Yamasaki, T.; Thi Hong Phung, L.; Karuaera, N.; Daniels, S.R.; GavishRegev, E.; Sharma, P.P. Phylogenomic data reveal three new families of poorly studied Solifugae (camel spiders). Mol. Phylogenetics Evol. 2023, 191, 107989. [Google Scholar] [CrossRef]
- World Solifugae Catalog. Available online: http://wac.nmbe.ch (accessed on 4 January 2024).
- Latreille, P.A. Galéode. In Nouveau Dictionnaire D’histoire Naturelle, Appliquée aux Arts, à L’agriculture, à L’économie Rurale et Domestique, à la Médecine, etc.; Par une société de naturalistes et d’agriculteurs: Paris, France, 1817; Volume 12, pp. 368–374. [Google Scholar]
- Muma, M.H. Basic behavior of North American Solpugida. Fla. Entomol. 1967, 50, 115–123. [Google Scholar] [CrossRef]
- Harvey, M.S. Nomenclatural notes on Solifugae, Amblypygi, Uropygi and Araneae (Arachnidda). Rec. West. Aust. Mus. 2002, 20, 449–459. [Google Scholar]
- Vogt, L.; Bartolomaeus, T.; Giribet, G. The linguistic problem of morphology: Structure versus homology and the standardization of morphological data. Cladistics 2010, 26, 301–325. [Google Scholar] [CrossRef] [PubMed]
- Maury, E.A. Nota sobre los géneros Namibesia y Syndaesia (Solifugae, Daesiidae). Aracnología 1985, 4, 1–6. [Google Scholar]
- Koch, L.C. Arachnoidea. In W.G. Rosenhauer, Die Thiere Andalusiens nach dem Resultate einer Reise Zusammengestellt, Nebst den Beschreibungen von 249 Neuen Oder bis Jetzt noch Unbeschreibenen Gattungen un Arten; Theodor Blaesing: Erlangen, Germany, 1856; pp. 406–413. [Google Scholar]
- Simon, E. Essai d’une classification des Galéodes, remarques synonymiques et description d’espèces nouvelles ou mal connues. Ann. Société Entomol. Fr. 1879, 9, 93–154. [Google Scholar]
- Pocock, R.I. Descriptions of four new Arachnida of the orders Pedipalpi, Solifugae, and Araneae. In Annals and Magazine of Natural History, Including Zoology, Botany and Geology, Being a Continuation of the “Magazine of Botany and Zoology”, and of Louden and Charlesworth’s “Magazine of Natural History”; Taylor and Francis, Ltd.: London, UK, 1903; Volume 11, pp. 220–226. [Google Scholar]
- Delle Cave, L.; Simonetta, A.M. A tentative revision of Daesiidae (Arachnida, Solifugae) from Ethiopia and Somalia. Monit. Zool. Ital. 1971, 4, 37–77. [Google Scholar]
- Maury, E.A. Solifugos de Colombia y Venezuela (Solifugae, Ammotrechidae). J. Arachnol. 1982, 10, 123–143. [Google Scholar]
- Rambla, M.; Barrientos, J.A. Nuevos datos sobre Gluvia dorsalis (Latreille, 1817) (Solifugae, Daesiidae). In Proceedings of the Ninth International Congress of Arachnology [9th International Congress of Arachnology, Panama City (Panama)], Panama City, Panama, 1–8 August 1983; Egerhard, W.G., Lubin, Y.D., Robinson, B.C., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1986; pp. 213–218. [Google Scholar]
- González-Moliné, A.L.; Melic, A.; Barrientos, J.A. Taxonomía, distribución geográfica e historia natural del endemismo iberico Gluvia dorsalis (Latreille, 1817) (Solifuguae: Daesiidae). Boletín Soc. Entomológica Aragonesa 2008, 42, 385–395. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; Waard, J.R. Biological identifications through DNA barcodes. Philos. Trans. R. Soc. 2003, B270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Mondragón, A.; Cortez, M.R. COI mtDNA barcoding and morphology for the description of a new species of ricinuleid of the genus Pseudocellus (Arachnida: Ricinulei: Ricinoididae) from El Triunfo Biosphere Reserve, Chiapas, Mexico. Eur. J. Taxon. 2021, 778, 1–25. [Google Scholar] [CrossRef]
- Young, M.R.; Moraza, M.L.; Ueckermann, E.; Heylen, D.; Baardsen, L.F.; Lima-Barbero, J.F.; Gal, S.; Gavish-Regev, E.; Gottlieb, Y.; Roy, L.; et al. Linking morphological and molecular taxonomy for the identification of poultry house, soil, and nest dwelling mites in the Western Palearctic. Sci. Rep. 2019, 9, 5784. [Google Scholar] [CrossRef] [PubMed]
- Muster, C.; Spelda, J.; Rulik, B.; Thormann, J.; von der Mark, L.; Astrin, J.J. The dark side of pseudoscorpion diversity: The German Barcode of Life campaign reveals high levels of undocumented diversity in European false scorpions. Ecol. Evol. Vol. 2021, 11, 13815–13829. [Google Scholar] [CrossRef] [PubMed]
- Friederich, S.; Lehman, T. Taito adrik, a new harvestman species from the Area de Conservacion Privada Panguana, Peruvian Amazonia (Opiliones: Laniatores: Cosmetidae). Zootaxa 2020, 4729, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.; Blagoev, G.; Tahir, H.M.; Khan, A.M.; Mukhtar, M.K.; Akhtar, S.; Butt, A.; Mansoor, S.; Hebert, P.D.N. Spiders. Assembling a DNA barcode reference library for the spiders (Arachnida: Araneae) of Pakistan. PLoS ONE 2019, 14, e0217086. [Google Scholar] [CrossRef]
- Valdez-Mondragón, A.; Navarro-Rodríguez, C.I.; Solís-Catalán, K.P.; Cortez-Roldán, M.R.; Juárez-Sánchez, A.R. Under an integrative taxonomic approach: The description of a new species of the genus Loxosceles (Araneae, Sicariidae) from Mexico City. ZooKeys 2019, 892, 93–133. [Google Scholar] [CrossRef]
- Trebicki, L.; Patoleta, B.; Dabert, M.; Zabka, M. Redescription of type species of the genus Cytaea Keyserling, 1882 (Araneae: Salticidae)—An integrative approach. Eur. Zool. J. 2021, 88, 933–947. [Google Scholar] [CrossRef]
- Ortiz, D.; Francke, O.F. Reconciling morphological and molecular systematics in tarantulas (Araneae: Theraphosidae): Revision of the Mexican endemic genus Bonnetina. Zool. J. Linn. Soc. 2017, 180, 819886. [Google Scholar] [CrossRef]
- Nolasco, S.; Valdez-Mondragón, A. To be or not to be… Integrative taxonomy and species delimitation in the daddy long-legs spiders of the genus Physocyclus (Araneae, Pholcidae) using DNA barcoding and morphology. Zookeys 2022, 1135, 93–118. [Google Scholar] [CrossRef] [PubMed]
- Więcek, M.; Szydło, W.; Dabert, J.; Proctor, H. Delimiting species of water mites of the genus Hydrodroma (Acari: Hydrachnidiae: Hydrodromidae) from North America and Europe: Integrative evidence of species status from COI sequences and morphology. Zool. Anz. 2020, 284, 16–29. [Google Scholar] [CrossRef]
- Barranco, P.; Gilgado, J.D.; Ortuño, V.M. A new mute species of the genus Nemobius Serville (Orthoptera, Gryllidae, Nemobiinae) discovered in colluvial, stony debris in the Iberian Peninsula: A biological, phenological and biometric study. Zootaxa 2013, 3691, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Baquero, E.; Ledesma, E.; Gilgado, J.D.; Ortuño, V.M.; Jordana, R. Distinctive Collembola communities in the mesovoid shallow substratum: First data for the sierra de Guadarrama national park (Central Spain) and a description of two new species of Orchesella (Entomobryidae). PLoS ONE 2017, 12, e0189205. [Google Scholar] [CrossRef] [PubMed]
- Gilgado, J.; Enghoff, H.; Ortuño, V.M. The hypogean Iberian genus Typhlopsychrosoma Mauriés 1982 (Diplopoda, Chordeumatida, Vandeleumatidae): Distribution map, key to species, first record in a Mesovoid Shallow Substratum (MSS) and detailed iconography of T. baeticaense (Mauriés 2013). Zootaxa 2015, 3937, 337–346. [Google Scholar] [CrossRef]
- Gilgado, J.; Enghoff, H.; Tinaut, A.; Mauriés, J.P.; Ortuño, V.M. Sierra Nevada (Granada, Spain): A high-altitude biogeographical crossroads for millipedes (Diplopoda), with first data on its MSS fauna and description of a new species of the genus Ceratosphys Ribaut, 1920 (Chordeumatida: Opisthocheiridae). Zootaxa 2015, 4044, 391–410. [Google Scholar] [CrossRef]
- Gilgado, J.; Enghoff, H.; Tinaut, A.; Ortuño, V.M. Hidden biodiversity in the Iberian mesovoid shallow substratum (MSS): New and poorly known species of the millipede genus Archipolydesmus Attems, 1898 (Diplopoda, Polydesmidae). Zool. Anz. 2015, 258, 13–38. [Google Scholar] [CrossRef]
- Gilgado, J.D.; Ledesma, E.; Enghoff, H.; Mauriés, J.P.; Ortuño, V.M. A new genus and species of Haplobainosomatidae (Diplopoda: Chordeumatida) from the MSS of the sierra de Guadarrama national park, central Spain. Zootaxa 2017, 4347, 492–510. [Google Scholar] [CrossRef]
- Peinado, M.; Alcaraz, F.; Martínez-Parras, J.M. Vegetation of Southeastern Spain. In Flora et Vegetatio Mundi; E.Schweizerbart’sche Verlagsbuchhandlung, Science Publishers: Stuttgart, Germany, 1992. [Google Scholar]
- Botero-Trujillo, R.; Martínez, L.; Iuri, H.A.; Ojanguren-Affilastro, A.A.; Carvalho, L.S. Revision of the genera Eutrecha and Xenotrecha (Solifugae: Ammotrechidae), taxonomic notes on Ammotrechinae, and description of a remarkable new Eutrecha from Colombia. Arthropod Syst. Phylogeny 2023, 81, 317–344. [Google Scholar] [CrossRef]
- Pagès, J. Analyse Factorielle Multiple Appliquée Aux Variables Qualitatives et Aux Données Mixtes. Rev. Stat. Appl. 2002, 4, 5–37. [Google Scholar]
- LeSebastien, J.J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kuhn, M. Building Predictive Models in R Using the caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 26 July 2023).
- Korkmaz, S.; Goksuluk, D.; Zararsiz, G. MVN: An R Package for Assessing Multivariate Normality. R J. 2014, 6, 151–162. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 14 January 2024).
- daSilva, A.R. Biotools: Tools for Biometry and Applied Statistics in Agricultural Science. R Package Version 4.2. 2021. Available online: https://cran.r-project.org/package=biotools (accessed on 14 January 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 15 March 2023).
- Shokralla, S.; Porter, T.M.; Gibson, J.F.; Dobosz, R.; Janzen, D.H.; Hallwachs, W.; Golding, G.B.; Hajibabaei, M. Massively parallel multiplex DNA sequencing for specimen identification using an Illumina MiSeq platform. Sci. Rep. 2015, 5, 9687. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Ji, Y.; Emerson, B.; Wang, X.; Ye, C.; Yang, C.; Ding, Z. Biodiversity soup: Metabarcoding of arthropods for rapid biodiversity assessment and biomonitoring. Methods Ecol. Evol. 2012, 3, 613–623. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.L. Systematische Uebersich über die familie der Galeoden. Arch. Naturgeschichte 1842, 8, 350–356. [Google Scholar]
- Pablos, F. Sobre la presencia de Gluvia dorsalis, var. conquensis en los puertos de Contreras (Cuenca). Publicaciones Inst. Biol. Apl. 1967, 43, 45–48. [Google Scholar]
- Legendre, P.; Oksanen, J.; ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Naseem, S.; Muhammad Tahir, H. Use of mitochondrial COI gene for the identification of family Salticidae and Lycosidae of spiders. Mitochondrial DNA Part A 2018, 29, 96–101. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Locality | Data | Code | GenBank Access Number |
|---|---|---|---|---|
| Gluvia brunnea sp.nov. | Cabo de Gata, Almería | 19 May 2022 | CDG-B05 | PP229911 |
| Gluvia brunnea sp.nov. | Cabo de Gata, Almería | 11 July 2022 | CDG-B06 | PP229912 |
| Gluvia brunnea sp.nov. | Sierra de Gádor, Almería | 10 July 2021 | EI2-D3 | PP229913 |
| Gluvia brunnea sp.nov. | Sierra de Gádor, Almería | 14 August 2023 | GAD-C11 | PP229914 |
| Gluvia dorsalis | Huelva | 01 June 2018 | CCPP-SOLIFUGO 5 | PP229915 |
| Gluvia dorsalis | Huelva | 18 June | CCPP-SOLIFUGO 8 | PP229916 |
| Gluvia dorsalis | Huelva | 18 June | CCPP-SOLIFUGO 1 | PP229917 |
| Gluvia dorsalis | Sierra de Gata, Cáceres | 30 June 2008 | EXT-B11 | PP229918 |
| Gluvia dorsalis | Alcolea, Córdoba | 28 July 2023 | CCPP-0495 | PP229919 |
| Gluvia dorsalis | Alcolea, Córdoba | 28 July 2023 | CCPP-0496 | PP229920 |
| Gluvia dorsalis | Alcolea, Córdoba | 28 July 2023 | CCPP-0497 | PP229921 |
| Gluvia dorsalis | El Patriarca, Córdoba | 29 April 2023 | Mmármol | PP229922 |
| Gluvia dorsalis | Málaga ciudad | 07 August 2001 | SOLI-2 | PP229923 |
| Gluvia dorsalis | Colmenar Viejo, Madrid | 22 July 2023 | CCPP-0498 | PP229924 |
| Gluvia dorsalis | Colmenar Viejo, Madrid | 22 July 2023 | CCPP-0499 | PP229925 |
| Gluvia dorsalis | - | - | - | MD038 |
| Gluviopsis nigrocinctus | - | - | - | SOL162 |
| Eremobates scopulatellus | - | - | - | DMNS:ZA19206 |
| Galumna dimorpha | Cabo de Gata, Almería | 05 February 2021 | - | PP228019 |
| Holotype | Males | Paratype Female | Females | ||
|---|---|---|---|---|---|
| Body length | 12.95 | 8.31–15.18 | 20.65 | 11.12–20.65 | |
| Chelicerae | Dorsal width | 1.24 | 0.89–1.32 | 2.55 | 1.36–2.55 |
| Lateral width | 1.26 | 0.96–1.48 | 2.83 | 1.47–2.83 | |
| Lateral length | 3.5 | 3.08–4.11 | 6.72 | 4.67–6.72 | |
| Flagellum length | 1.57 | 1.17–1.71 | - | - | |
| Flagellum distal lobe width | 0.54 | 0.42–0.65 | - | - | |
| Flagellum proximal lobe width | 0.33 | 0.29–0.37 | - | - | |
| Propeltidium | Area | 4.27 | 2.67–5.67 | 15.97 | 5.44–15.97 |
| Flatness ratio | 0.84 | 0.8–0.87 | 0.66 | 0.62–0.71 | |
| Width | 2.76 | 2.16–3.17 | 5.77 | 3.41–5.77 | |
| Length | 2.32 | 1.79–2.64 | 3.91 | 2.37–3.91 | |
| Ocular tubercle (area) | 0.31 | 0.19–0.42 | 0.51 | 0.27–0.51 | |
| Interocular | 0.24 | 0.15–0.29 | 0.38 | 0.17–0.38 | |
| Eye length | 0.3 | 0.21–0.33 | 0.31 | 0.24–0.36 | |
| Pedipalp | Telotarsus | 0.96 | 0.72–0.99 | 0.82 | 0.68–1.1 |
| Basitarsus | 2.79 | 2.29–3.28 | 3.54 | 2.63–3.54 | |
| Tibia | 3.62 | 2.88–4.29 | 4.35 | 2.86–4.35 | |
| Femur | 4.12 | 3.04–4.68 | 4.53 | 2.82–4.53 | |
| TOTAL | 11.49 | 8.95–13.23 | 13.24 | 9.01–13.24 | |
| Leg I | Telotarsus | 1.45 | 0.84–1.45 | 1.46 | 1.02–1.87 |
| Basitarsus | 1.52 | 1.13–2.18 | 2.38 | 1.16–2.38 | |
| Tibia | 2.81 | 2.36–3.67 | 3.91 | 2.17–3.91 | |
| Femur | 2.73 | 1.61–2.78 | 2.74 | 1.5–2.86 | |
| TOTAL | 8.51 | 6.45–10.05 | 10.49 | 6.33–10.49 | |
| Leg II | Claw | 0.69 | 0.45–0.81 | 0.75 | 0.57–0.8 |
| Telotarsus | 0.71 | 0.57–0.9 | 1.16 | 0.6–1.16 | |
| Basitarsus | 1.15 | 1.02–1.57 | 1.84 | 1.13–1.84 | |
| Tibia | 1.55 | 1.43–1.96 | 2.4 | 1.53–2.4 | |
| Femur | 1.86 | 1.21–2.26 | 1.89 | 1.27–1.91 | |
| TOTAL | 5.96 | 4.68–7.5 | 8.04 | 5.14–8.04 | |
| Leg III | Claw | 0.86 | 0.55–0.93 | 0.72 | 0.69–0.96 |
| Telotarsus | 0.9 | 0.65–0.99 | 1.33 | 0.71–1.33 | |
| Basitarsus | 1.69 | 1.26–1.96 | 2.48 | 1.38–2.48 | |
| Tibia | 2.19 | 1.68–3.07 | 3.18 | 1.78–3.18 | |
| Femur | 2.78 | 1.82–3.18 | 3.02 | 1.93–3.02 | |
| TOTAL | 8.42 | 6.6–10.03 | 10.73 | 6.75–10.73 | |
| Leg IV | Claw | 1.42 | 0.99–1.38 | 1.26 | 0.91–1.31 |
| Telotarsus | 1.74 | 1.47–2.09 | 2.23 | 1.17–2.23 | |
| Basitarsus | 3.71 | 2.56–4.24 | 4.02 | 2.54–4.02 | |
| Tibia | 5.02 | 3.46–5.71 | 5.68 | 3.31–5.68 | |
| Femur | 4.63 | 3.34–5.5 | 5.51 | 2.77–5.51 | |
| TOTAL | 16.2 | 10.94–18.92 | 18.7 | 10.8–18.7 | |
| Females | ||
| Group | Dim.1 | Dim.2 |
| qualitative | 10.00 | 23.94 |
| propeltidium | 16.59 | 28.53 |
| eyes | 22.09 | 0.80 |
| palps | 21.63 | 6.83 |
| legs | 27.15 | 4.31 |
| chelicerae | 1.31 | 19.61 |
| stridulatory organ | 1.24 | 15.97 |
| Males | ||
| Group | Dim.1 | Dim.2 |
| qualitative | 11.48 | 15.36 |
| propeltidium | 11.72 | 32.44 |
| eyes | 11.40 | 10.03 |
| palps | 12.86 | 8.67 |
| legs | 14.59 | 9.09 |
| chelicerae | 11.37 | 12.39 |
| stridulatory organ | 9.51 | 3.09 |
| flagellum | 17.07 | 8.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pertegal, C.; Barranco, P.; De Mas, E.; Moya-Laraño, J. More Than 200 Years Later: Gluvia brunnea sp. nov. (Solifugae, Daesiidae), a Second Species of Camel Spider from the Iberian Peninsula. Insects 2024, 15, 284. https://doi.org/10.3390/insects15040284
Pertegal C, Barranco P, De Mas E, Moya-Laraño J. More Than 200 Years Later: Gluvia brunnea sp. nov. (Solifugae, Daesiidae), a Second Species of Camel Spider from the Iberian Peninsula. Insects. 2024; 15(4):284. https://doi.org/10.3390/insects15040284
Chicago/Turabian StylePertegal, Cristian, Pablo Barranco, Eva De Mas, and Jordi Moya-Laraño. 2024. "More Than 200 Years Later: Gluvia brunnea sp. nov. (Solifugae, Daesiidae), a Second Species of Camel Spider from the Iberian Peninsula" Insects 15, no. 4: 284. https://doi.org/10.3390/insects15040284
APA StylePertegal, C., Barranco, P., De Mas, E., & Moya-Laraño, J. (2024). More Than 200 Years Later: Gluvia brunnea sp. nov. (Solifugae, Daesiidae), a Second Species of Camel Spider from the Iberian Peninsula. Insects, 15(4), 284. https://doi.org/10.3390/insects15040284