
Validation of Diets with Tomato Pomace in Complete Cycle Breeding of Tenebrio molitor (L.) (Coleoptera: Tenebrionidae)
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Compositions
2.2. Insects and Experimental Set-Up
2.2.1. Experiment A
2.2.2. Experiment B
2.3. Statistical Analysis
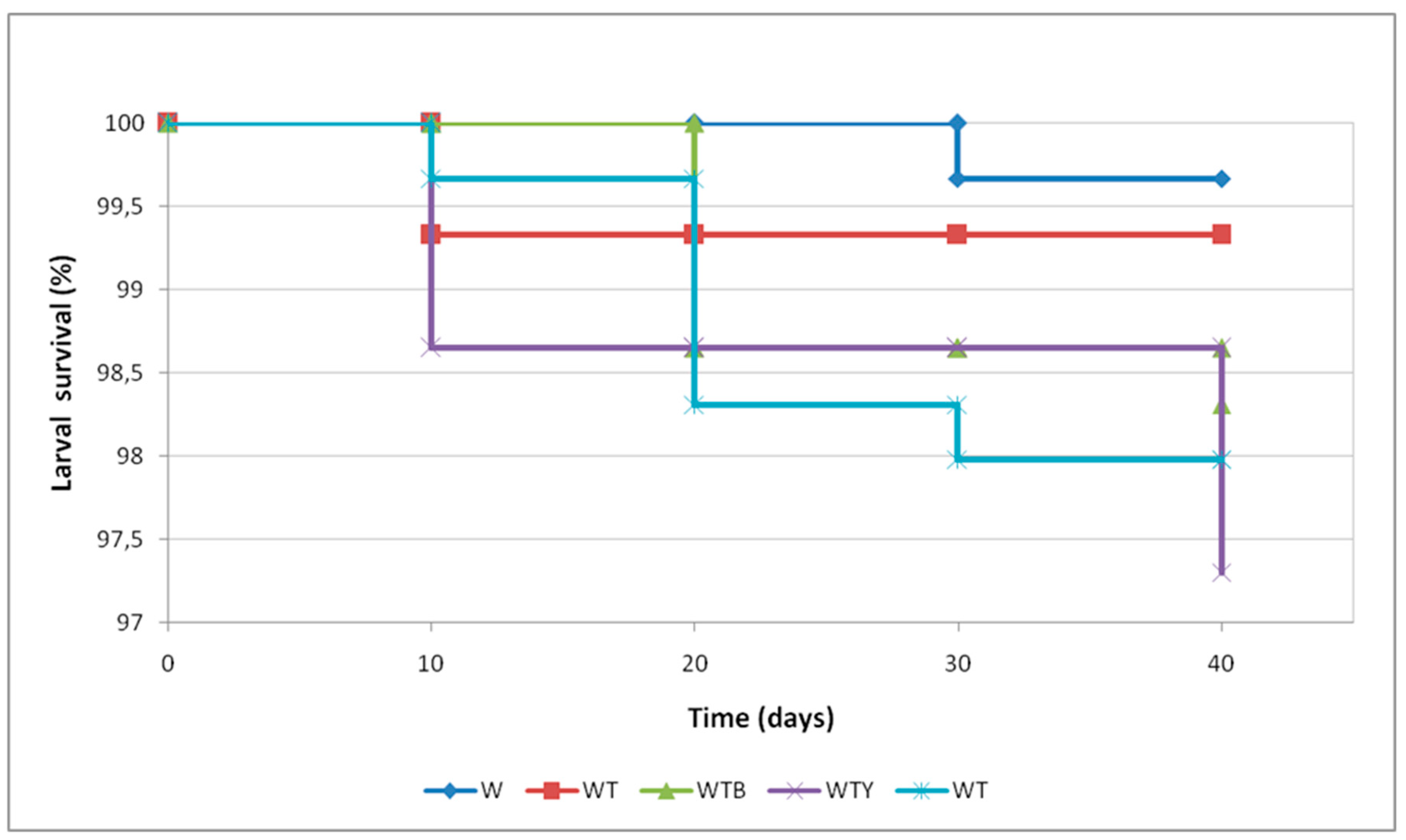
3. Results
3.1. Experiment A
3.2. Experiment B
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security. Rome Food Agric. Organ. United Nations 2013, 171, 187. [Google Scholar]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable Farming of the Mealworm Tenebrio Molitor for the Production of Food and Feed. Z. Für Naturforsch. C 2017, 72, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect Rearing: Potential, Challenges, and Circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Nava, A.L.; Higareda, T.E.; Barreto, C.; Rodríguez, R.; Márquez, I.; Palacios, M.L. Circular Economy Approach for Mealworm Industrial Production for Human Consumption. IOP Conf. Ser. Earth Environ. Sci. 2020, 463, 012087. [Google Scholar] [CrossRef]
- Smetana, S.; Palanisamy, M.; Mathys, A.; Heinz, V. Sustainability of Insect Use for Feed and Food: Life Cycle Assessment Perspective. J. Clean. Prod. 2016, 137, 741–751. [Google Scholar] [CrossRef]
- Mouhrim, N.; Peguero, D.A.; Green, A.; Silva, B.; Bhatia, A.; Ristic, D.; Tonda, A.; Mathys, A.; Smetana, S. Optimization Models for Sustainable Insect Production Chains. J. Insects Food Feed 2023, 1, 1–19. [Google Scholar] [CrossRef]
- Niyonsaba, H.H.; Groeneveld, I.L.; Vermeij, I.; Höhler, J.; van der Fels-Klerx, H.J.; Meuwissen, M.P.M. Profitability of Insect Production for T. Molitor Farms in The Netherlands. J. Insects Food Feed 2023, 1, 1–8. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Bliamplias, D.; Gourgouta, M.; Michail, V.; Athanassiou, C.G. Rearing Tenebrio molitor and Alphitobius diaperinus Larvae on Seed Cleaning Process Byproducts. Insects 2021, 12, 293. [Google Scholar] [CrossRef] [PubMed]
- Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas Morio, Tenebrio Molitor, and Acheta Domesticus Larvae as Alternative Feed Supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef]
- Bordiean, A.; Krzyżaniak, M.; Aljewicz, M.; Stolarski, M.J. Influence of Different Diets on Growth and Nutritional Composition of Yellow Mealworm. Foods 2022, 11, 3075. [Google Scholar] [CrossRef]
- Kröncke, N.; Benning, R. Influence of Dietary Protein Content on the Nutritional Composition of Mealworm Larvae (Tenebrio molitor L.). Insects 2023, 14, 261. [Google Scholar] [CrossRef]
- Kröncke, N.; Neumeister, M.; Benning, R. Near-Infrared Reflectance Spectroscopy for Quantitative Analysis of Fat and Fatty Acid Content in Living Tenebrio molitor Larvae to Detect the Influence of Substrate on Larval Composition. Insects 2023, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Boukid, F.; Riudavets, J.; del Arco, L.; Castellari, M. Impact of Diets Including Agro-Industrial By-Products on the Fatty Acid and Sterol Profiles of Larvae Biomass from Ephestia Kuehniella, Tenebrio molitor and Hermetia Illucens. Insects 2021, 12, 672. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Spagnoletta, A.; Lamaj, F.; Vitale, M.L.; Verrastro, V. First Optimization of Tomato Pomace in Diets for Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects 2023, 14, 854. [Google Scholar] [CrossRef] [PubMed]
- Kotsou, K.; Chatzimitakos, T.; Athanasiadis, V.; Bozinou, E.; Rumbos, C.I.; Athanassiou, C.G.; Lalas, S.I. Enhancing the Nutritional Profile of Tenebrio molitor Using the Leaves of Moringa Oleifera. Foods 2023, 12, 2612. [Google Scholar] [CrossRef] [PubMed]
- Nesci, S.; Spagnoletta, A.; Oppedisano, F. Inflammation, Mitochondria and Natural Compounds Together in the Circle of Trust. Int. J. Mol. Sci. 2023, 24, 6106. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, M.; Spranghers, T.; De Clercq, P.; Cooreman-Algoed, M.; Couchement, T.; De Clercq, G.; Verbeke, S.; Spanoghe, P. Pesticide Contamination of Tenebrio molitor (Coleoptera: Tenebrionidae) for Human Consumption. Food Chem. 2016, 201, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Naccarato, A.; Vommaro, M.L.; Amico, D.; Sprovieri, F.; Pirrone, N.; Tagarelli, A.; Giglio, A. Triazine Herbicide and NPK Fertilizer Exposure: Accumulation of Heavy Metals and Rare Earth Elements, Effects on Cuticle Melanization, and Immunocompetence in the Model Species Tenebrio molitor. Toxics 2023, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Candian, V.; Tedeschi, R. Impact of the Diet on the Mortality and on Gene Expression of the Antimicrobial Peptide Tenecin 3 in Tenebrio Molitor Larvae Infected by Beauveria Bassiana. Insects 2023, 14, 359. [Google Scholar] [CrossRef] [PubMed]
- Dahal, S.; Jensen, A.B.; Lecocq, A. Effect of Probiotics on Tenebrio molitor Larval Development and Resistance against the Fungal Pathogen Metarhizium Brunneum. Insects 2022, 13, 1114. [Google Scholar] [CrossRef]
- Morales Ramos, J.; Rojas, M.-G.; Shapiro Ilan, D.; TEDDERS, W. Developmental Plasticity in Tenebrio molitor (Coleoptera: Tenebrionidae): Analysis of Instar Variation in Number and Development Time under Different Diets. J. Entomol. Sci. 2010, 45, 75–90. [Google Scholar] [CrossRef]
- Ribeiro, N.; Abelho, M.; Costa, R. A Review of the Scientific Literature for Optimal Conditions for Mass Rearing Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2018, 53, 434–454. [Google Scholar] [CrossRef]
- Kröncke, N.; Benning, R. Self-Selection of Feeding Substrates by Tenebrio Molitor Larvae of Different Ages to Determine Optimal Macronutrient Intake and the Influence on Larval Growth and Protein Content. Insects 2022, 13, 657. [Google Scholar] [CrossRef]
- Rumbos, C.I.; Karapanagiotidis, I.T.; Mente, E.; Psofakis, P.; Athanassiou, C.G. Evaluation of Various Commodities for the Development of the Yellow Mealworm, Tenebrio Molitor. Sci. Rep. 2020, 10, 11224. [Google Scholar] [CrossRef] [PubMed]
- Lienhard, A.; Rehorska, R.; Pöllinger-Zierler, B.; Mayer, C.; Grasser, M.; Berner, S. Future Proteins: Sustainable Diets for Tenebrio molitor Rearing Composed of Food By-Products. Foods 2023, 12, 4092. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ramos, J.A.; Rojas, M.G.; Shapiro-llan, D.I.; Tedders, W.L. Use of Nutrient Self-Selection as a Diet Refining Tool in Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2013, 48, 206–221. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Kelstrup, H.C.; Emery, V. Self-Selection of Agricultural By-Products and Food Ingredients by Tenebrio molitor (Coleoptera: Tenebrionidae) and Impact on Food Utilization and Nutrient Intake. Insects 2020, 11, 827. [Google Scholar] [CrossRef]
- Errico, S.; Dimatteo, S.; Moliterni, S.; Baldacchino, F. Effects of Long-Lasting Cold Storage on Tenebrio molitor Larvae (Coleoptera: Tenebrionidae). J. Insects Food Feed 2021, 7, 1111–1116. [Google Scholar] [CrossRef]
- Manojlović, B. A Contribution to the Study of the Influence of the Feeding of Imagos and of Climatic Factors on the Dynamics of Oviposition and on the Embryonal Development of Yellow Mealworm Tenebrio molitor L. Coleoptera Tenebrionidae. Biol. Environ. Sci. 1987, 38, 337–448. [Google Scholar]
- El Deen, S.N.; Lamaj, F.; Verrastro, V.; Al Bitar, L.; Baldacchino, F. Effects of Two Diets on Adults’ Survival and Productivity in Mass-Rearing of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Insects Food Feed 2021, 7, 1149–1157. [Google Scholar] [CrossRef]
- Berggreen, I.E.; Offenberg, J.; Calis, M.; Heckmann, L.-H. Impact of Density, Reproduction Period and Age on Fecundity of the Yellow Mealworm Tenebrio molitor (Coleoptera: Tenebrionidae). J. Insects Food Feed 2018, 4, 43–50. [Google Scholar] [CrossRef]
- Veldkamp, T.; Claeys, J.; Haenen, O.L.M.; van Loon, J.J.A.; Spranghers, T. (Eds.) The Basics of Edible Insect Rearing; Wageningen Academic Publishers: Amsterdam, The Netherlands, 2021; ISBN 978-90-8686-365-5. [Google Scholar]
- Naser El Deen, S.; Spranghers, T.; Baldacchino, F.; Deruytter, D. The Effects of the Particle Size of Four Different Feeds on the Larval Growth of Tenebrio molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2022, 119, 242–249. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists International, 18th ed.; AOAC International: Gaitherburg, MD, USA, 2005. [Google Scholar]
- Bhattacharya, A.; Ameel, J.; Waldbauer, G. A Method for Sexing Living Pupal and Adult Yellow Mealworms. Ann. Entomol. Soc. Am. 1970, 63, 1783. [Google Scholar] [CrossRef]
- Weaver, D.K.; McFarlane, J.E. The Effect of Larval Density on Growth and Development of Tenebrio molitor. J. Insect Physiol. 1990, 36, 531–536. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Kay, S.; Shapiro-Ilan, D.I.; Tedders, W.L. Impact of Adult Weight, Density, and Age on Reproduction of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Entomol. Sci. 2012, 47, 208–220. [Google Scholar] [CrossRef]
- Urrejola, S.; Nespolo, R.; Lardies, M.A. Diet-Induced Developmental Plasticity in Life Histories and Energy Metabolism in a Beetle. Rev. Chil. Hist. Nat. 2011, 84, 523–533. [Google Scholar] [CrossRef]
- Giuffrè, A.M.; Capocasale, M. Physicochemical Composition of Tomato Seed Oil for an Edible Use: The Effect of Cultivar. Int. Food Res. J. 2016, 23, 583–591. [Google Scholar]
- Lazos, E.S.; Tsaknis, J.; Lalas, S. Characteristics and Composition of Tomato Seed Oil. Grasas Aceites 1998, 49, 440–445. [Google Scholar] [CrossRef]
- Haas, E.; Kim, Y.; Stanley, D. Why Can Insects Not Biosynthesize Cholesterol? Arch. Insect Biochem. Physiol. 2023, 112, e21983. [Google Scholar] [CrossRef] [PubMed]
- Clayton, R.B. The Utilization of Sterols by Insects. J. Lipid Res. 1964, 5, 3–19. [Google Scholar] [CrossRef]
- Monroe, R.E. Effect of Dietary Cholesterol on House Fly Reproduction1. Ann. Entomol. Soc. Am. 1960, 53, 821–824. [Google Scholar] [CrossRef]
- Elbadrawy, E.; Sello, A. Evaluation of Nutritional Value and Antioxidant Activity of Tomato Peel Extracts. Arab. J. Chem. 2016, 9, S1010–S1018. [Google Scholar] [CrossRef]
- Sangeetha, K.; Ramyaa, R.B.; Mousavi Khaneghah, A.; Radhakrishnan, M. Extraction, Characterization, and Application of Tomato Seed Oil in the Food Industry: An Updated Review. J. Agric. Food Res. 2023, 11, 100529. [Google Scholar] [CrossRef]
- Haeri, F.; Nouri, M.; Sadrmanesh, O.; Shirani, M.; Ghiasvand, R. The Relationship between the Intake of Dietary Fatty Acids and Minerals with Sperm Parameters in Infertile Men. Clin. Nutr. ESPEN 2023, 58, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Stoffel, W.; Schmidt-Soltau, I.; Binczek, E.; Thomas, A.; Thevis, M.; Wegner, I. Dietary Ω3-and Ω6-Polyunsaturated Fatty Acids Reconstitute Fertility of Juvenile and Adult Fads2-Deficient Mice. Mol. Metab. 2020, 36, 100974. [Google Scholar] [CrossRef] [PubMed]
- Roszkos, R.; Tóth, T.; Mézes, M. Review: Practical Use of n-3 Fatty Acids to Improve Reproduction Parameters in the Context of Modern Sow Nutrition. Animals 2020, 10, 1141. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.; Wang, J.; Lu, W. Regulation of Semen Quality by Fatty Acids in Diets, Extender, and Semen. Front. Vet. Sci. 2023, 10, 1119153. [Google Scholar] [CrossRef] [PubMed]
- Hoxha, M.; Barbonetti, A.; Zappacosta, B. Arachidonic Acid Pathways and Male Fertility: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 8207. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential Effects of Prostaglandin Derived from ω-6 and ω-3 Polyunsaturated Fatty Acids on COX-2 Expression and IL-6 Secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D. Prostaglandins and other eicosanoids in insects: Biological Significance. Annu. Rev. Entomol. 2006, 51, 25–44. [Google Scholar] [CrossRef]
- Stanley-Samuelson, D.W.; Peloquin, J.J.; Loher, W. Egg-Laying in Response to Prostaglandin Injections in the Australian Field Cricket, Teleogryllus Commodus. Physiol. Entomol. 1986, 11, 213–219. [Google Scholar] [CrossRef]
- Destephano, D.B.; Brady, U.E. Prostaglandin and Prostaglandin Synthetase in the Cricket, Acheta Domesticus. J. Insect Physiol. 1977, 23, 905–911. [Google Scholar] [CrossRef]
- Stanley, D.W. Eicosanoids in Invertebrate Signal Transduction Systems; Princeton University Press: Princeton, NJ, USA, 2000. [Google Scholar] [CrossRef]
- de Medeiros, M.N.; Belmonte, R.; Soares, B.C.C.; de Medeiros, L.N.; Canetti, C.; Freire-de-Lima, C.G.; Maya-Monteiro, C.M.; Bozza, P.T.; Almeida, I.C.; Masuda, H.; et al. Arrest of Oogenesis in the Bug Rhodnius Prolixus Challenged with the Fungus Aspergillus Niger Is Mediated by Immune Response-Derived PGE2. J. Insect Physiol. 2009, 55, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.A.; Ahmed, S.; Kim, Y. Biosynthetic Pathway of Arachidonic Acid in Spodoptera Exigua in Response to Bacterial Challenge. Insect Biochem. Mol. Biol. 2019, 111, 103179. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Ramírez, A.; Rocha-Ortega, M.; Córdoba-Aguilar, A. Dietary Macronutrient Balance and Fungal Infection as Drivers of Spermatophore Quality in the Mealworm Beetle. Curr. Res. Insect Sci. 2021, 1, 100009. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Stanley-Samuelson, D.W. Phospholipid Fatty Acid Composition and Arachidonic Acid Metabolism in Selected Tissues of Adult Tenebrio molitor (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 1990, 83, 975–981. [Google Scholar] [CrossRef]
- Zaidi, Z.S.; Ansari, H.J.; Khan, M.A. Phospholipid Concentration in the Fat Body and Gonads of Red Cotton Bug, Dysdercus Cingulatus Fabr. (Heteroptera: Pyrrhocoridae) in Relation to Growth, Metamorphosis, and Reproduction. Bolletino Zool. 1985, 52, 305–308. [Google Scholar] [CrossRef]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 Synthesis and Secretion: The Role of PGE2 Synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Stanley-Samuelson, D.W.; Jurenka, R.A.; Cripps, C.; Blomquist, G.J.; de Renobales, M. Fatty Acids in Insects: Composition, Metabolism, and Biological Significance. Arch. Insect Biochem. Physiol. 1988, 9, 1–33. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, S.; Seo, S.; Kim, K. Fatty Acid Composition of Different Tissues of Spodoptera Exigua Larvae and a Role of Cellular Phospholipase A2. Korean J. Appl. Entomol. 2016, 55, 129–138. [Google Scholar] [CrossRef]
- Kim, Y.; Ahmed, S.; Stanley, D.; An, C. Eicosanoid-Mediated Immunity in Insects. Dev. Comp. Immunol. 2018, 83, 130–143. [Google Scholar] [CrossRef]
- Cortes Ortiz, J.A.; Ruiz, A.T.; Morales-Ramos, J.A.; Thomas, M.; Rojas, M.G.; Tomberlin, J.K.; Yi, L.; Han, R.; Giroud, L.; Jullien, R.L. Insect Mass Production Technologies. In Insects as Sustainable Food Ingredients; Elsevier: London, UK, 2016; pp. 153–201. [Google Scholar]
- Han, J.-H.; Yang, Y.-X.; Feng, M.-Y. Contents of Phytosterols in Vegetables and Fruits Commonly Consumed in China. Biomed. Environ. Sci. 2008, 21, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Veen, M.; Lang, C. Production of Lipid Compounds in the Yeast Saccharomyces Cerevisiae. Appl. Microbiol. Biotechnol. 2004, 63, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Poerschmann, J.; Górecki, T. Characterization of the Lipid Soluble Fraction of Brewer’s Spent Grain. Curr. Chromatogr. 2016, 3, 86–98. [Google Scholar] [CrossRef]
- del Río, J.C.; Prinsen, P.; Gutiérrez, A. Chemical Composition of Lipids in Brewer’s Spent Grain: A Promising Source of Valuable Phytochemicals. J. Cereal Sci. 2013, 58, 248–254. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Dry Matter (%) | Crude Protein (%) | Crude Fat (%) | Crude Fiber (%) | Ash (%) | Carbohydrate (%) |
|---|---|---|---|---|---|---|
| Wheat bran | 91.2 | 16.7 | 6.5 | 36.1 | 4.2 | 30.2 |
| Tomato pomace | 92.1 | 9.5 | 3.2 | 67.1 | 3.9 | 8.9 |
| Brewer’s spent grain | 93.4 | 24.7 | 4.8 | 42.0 | 2.6 | 24.0 |
| Yeast | 93.0 | 47.6 | 2.4 | 6.8 | 8.0 | 13.8 |
| Diet | Wheat Bran (%) | Tomato Pomace (%) | Brewer’s Spent Grain (%) | Yeast (%) | Protein Value (%) | Carbohydrate (%) | Crude Fiber (%) | Fat (%) | Energy (kcal/100 g) |
|---|---|---|---|---|---|---|---|---|---|
| W | 100 | - | - | - | 16.7 | 30.2 | 36.1 | 6.5 | 318.2 |
| WY | 95 | - | - | 5 | 18.2 | 29.4 | 34.6 | 6.3 | 316.5 |
| WTB | 50 | 27 | 23 | - | 16.6 | 23.0 | 45.8 | 5.2 | 297.1 |
| WTY | 50 | 41 | - | 9 | 16.5 | 20.0 | 46.2 | 4.8 | 281.4 |
| WT | 50 | 50 | - | - | 13.1 | 19.6 | 51.6 | 4.9 | 277.5 |
| Diets | T1–10 | T11–20 | T21–30 | T31–40 | T41–50 | T51–60 | T61–70 |
|---|---|---|---|---|---|---|---|
| W | 85.6 ± 13.7 a | 59.4 ± 4.0 a | 57.7 ± 5.8 a | 100.2 ± 6.5 a | 83.7 ± 7.1 a | 94.6 ± 5.1 ab | 41.0 ± 3.0 a |
| WY | 71.5 ± 10.3 a | 74.4 ± 7.0 ab | 104.6 ± 10.8 bc | 95.7 ± 10.8 a | 86.7 ± 7.9 a | 107.4 ± 6.1 ab | 69.6 ± 5.6 b |
| WTB | 114.9 ± 9.7 ab | 98.5 ± 10.7 bc | 95.4 ± 10.7 ab | 128.2 ± 11.0 ab | 86.0 ± 6.4 a | 121.8 ± 9.2 ab | 46.4 ± 53 a |
| WTY | 151.6 ± 13.0 b | 140.4 ± 12.3 d | 140.5 ± 13.0 c | 157.9 ± 9.9 b | 88.5 ± 7.4 a | 125.8 ± 8.2 b | 71.8 ± 7.5 b |
| WT | 97.8 ± 13.2 a | 112.7 ± 8.3 cd | 117.1 ± 7.8 bc | 127.6 ± 8.0 ab | 109.2 ± 6.6 a | 92.2 ± 10.4 a | 83.3 ± 6.9 b |
| Diets | Larval Development Time (Days) | Early Pupation 1 (Days) | Weight Pupae (mg) | Sex Ratio (Male:Female) | Ingested Diet at T30 (g/replicate) |
|---|---|---|---|---|---|
| W | 45.1 ± 0.7 b | 0 | 116.1 ± 3.0 a | 50/50 | 9.1 ± 0.2 c |
| WY | 40.2 ± 0.7 a | −4.9 | 116.5 ± 3.2 a | 47/53 | 8.9 ± 0.2 c |
| WTB | 42.3 ± 0.9 ab | −2.8 | 125.0 ± 2.7 a | 51/49 | 10.0 ± 0.2 d |
| WTY | 39.5 ± 1.1 a | −5.6 | 121.7 ± 3.8 a | 42/58 | 7.9 ± 0.2 b |
| WT | 43.9 ± 0.9 b | −1.2 | 122.6 ± 2.9 a | 46/54 | 7.1 ± 0.2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldacchino, F.; Spagnoletta, A.; Lamaj, F.; Vitale, M.L.; Verrastro, V. Validation of Diets with Tomato Pomace in Complete Cycle Breeding of Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects 2024, 15, 287. https://doi.org/10.3390/insects15040287
Baldacchino F, Spagnoletta A, Lamaj F, Vitale ML, Verrastro V. Validation of Diets with Tomato Pomace in Complete Cycle Breeding of Tenebrio molitor (L.) (Coleoptera: Tenebrionidae). Insects. 2024; 15(4):287. https://doi.org/10.3390/insects15040287
Chicago/Turabian StyleBaldacchino, Ferdinando, Anna Spagnoletta, Flutura Lamaj, Maria Luisa Vitale, and Vincenzo Verrastro. 2024. "Validation of Diets with Tomato Pomace in Complete Cycle Breeding of Tenebrio molitor (L.) (Coleoptera: Tenebrionidae)" Insects 15, no. 4: 287. https://doi.org/10.3390/insects15040287