
Insecticide Resistance in Aedes aegypti Mosquitoes: Possible Detection of kdr F1534C, S989P, and V1016G Triple Mutation in Benin, West Africa

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Mosquito Sampling
2.2. Aedes Morphological Identification
2.3. Insecticide Resistance Bioassays
2.4. Measurement of Detoxification Enzyme Activity
2.5. Ae. aegypti kdr Genotyping
2.6. Data Analysis
3. Results
3.1. Mosquito Sampling
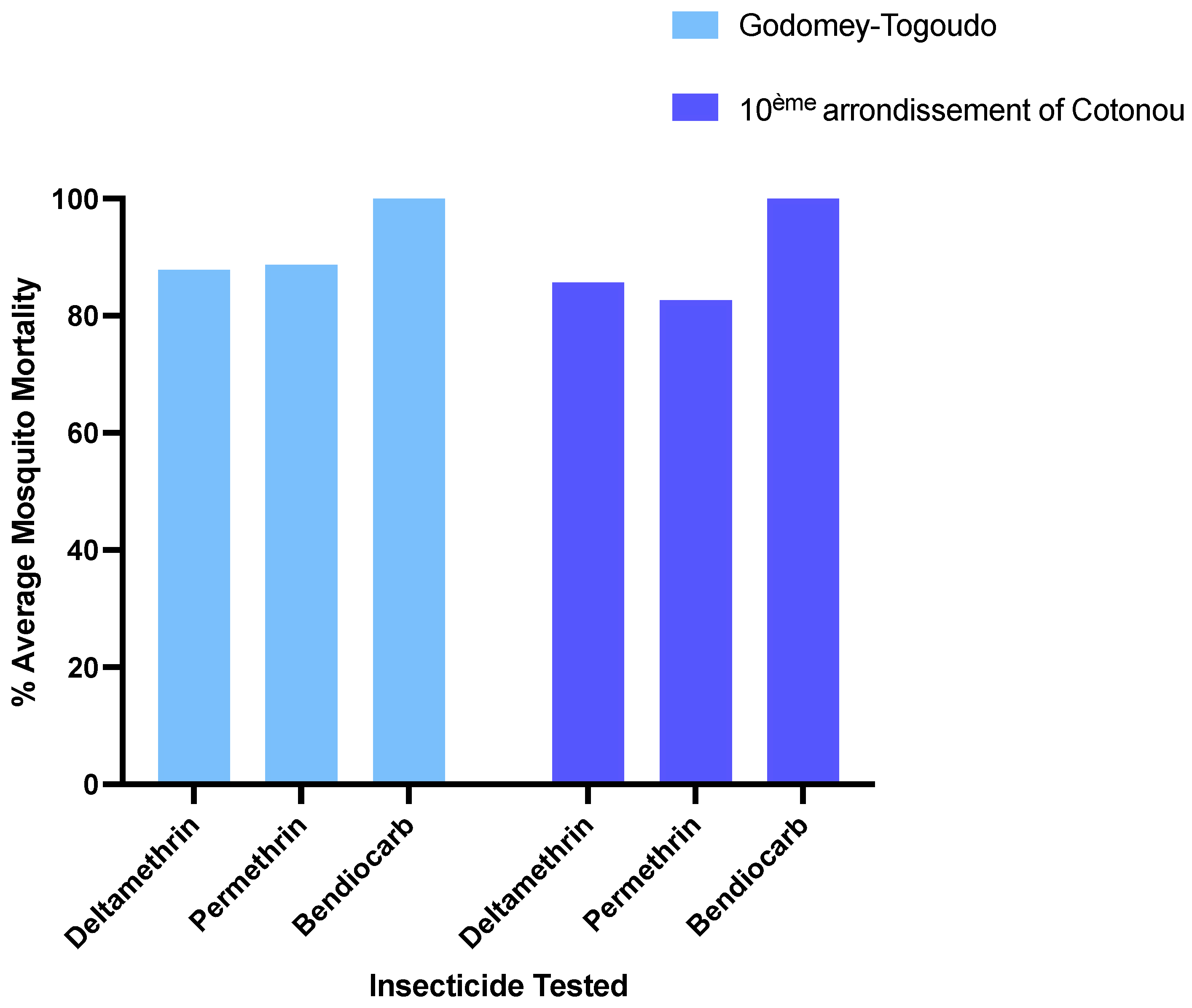
3.2. Insecticide Resistance Profiles
3.3. Expression of Detoxification Enzymes
3.4. kdr Mutation Screening
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dengue et Dengue Sévère. Available online: https://www.who.int/fr/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 20 January 2022).
- Padonou, G.G.; Ossè, R.; Salako, A.S.; Aikpon, R.; Sovi, A.; Kpanou, C.; Sgbaohan HAkadiri, Y.; Lamine, B.; Akogbeto, C.M. Entomological assessment of the risk of dengue outbreak in Abomey-Calavi Commune, Benin. Trop. Med. Health 2020, 48, 20. [Google Scholar] [CrossRef] [PubMed]
- Esu, E.; Lenhart, A.; Smith, L.; Horstick, O. Effectiveness of peridomestic space spraying with insecticide on dengue transmission; systematic review. Trop. Med. Int. Health 2010, 15, 619–631. [Google Scholar] [CrossRef]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Liu, N. Résistance aux insecticides chez les moustiques: Impact, mécanismes et directions de recherche. Ann. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Brogdon, W.G.; McAllister, J.C. Résistance aux insecticides et lutte antivectorielle. Urgence Infect. Dis. 1998, 4, 605–613. [Google Scholar]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.E.; Albeldaño-Vázquez, W.; Salas, I.F.; Badii, M.H.; Becerra, H.L.; Garcia, G.P.; Fuentes, S.L.; Brogdon, W.G.; Black Iv, W.C.; Beaty, B. Elevated α-esterase levels associated with permethrin tolerance in Aedes aegypti (L.) from Baja California, México. Pestic. Biochem. Phys. 2005, 82, 66–78. [Google Scholar] [CrossRef]
- Bisset, J.A.; Marin, R.; Rodriguez, M.M.; Severson, D.W.; Ricardo, Y.; French, L.; Díaz, M.; Perez, O. Insecticide resistance in two Aedes aegypti (Diptera: Culicidae) strains from Costa Rica. J. Med. Entomol. 2013, 50, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.R.; Longnecker, M.; Fredregill, C.L.; Debboun, M.; Pietrantonio, P.V. Kdr genotyping (V1016I, F1534C) of the Nav channel of Aedes aegypti (L.) mosquito populations in Harris County (Houston), Texas, USA, after Permanone 31–66 field tests and its influence on probability of survival. PLoS Negl. Trop. Dis. 2021, 15, e0009833. [Google Scholar] [CrossRef]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef]
- Li, C.X.; Kaufman, P.E.; Xue, R.D.; Zhao, M.H.; Wang, G.; Yan, T.; Guo, X.X.; Zhang, Y.M.; Dong, Y.D.; Xing, D.; et al. Relationship between insecticide resistance and kdr mutations in the dengue vector Aedes aegypti in Southern China. Parasites Vectors 2015, 8, 325. [Google Scholar] [CrossRef]
- Kawada, H.; Oo, S.Z.M.; Thaung, S.K.E.; Maung, Y.N.M.; Thu, H.M.; Thant, K.Z.; Minakawa, N. Co-occurrence of point mutations in the voltage-gated sodium channel of pyrethroid-resistant Aedes aegypti populations in Myanmar. PLoS Negl. Trop. Dis. 2014, 8, e3032. [Google Scholar] [CrossRef] [PubMed]
- Kawada, H.; Higa, Y.; Futami, K.; Muranami, Y.; Kawashima, E.; Osei, J.H.; Sakyi, K.Y.; Dadzie, S.; de Souza, D.K.; Appawu, M. Discovery of point mutations in the voltag egated sodium channel from African Aedes aegypti populations: Potential phylogenetic reasons for gene introgression. PLoS Negl. Trop. Dis. 2016, 10, e0004780. [Google Scholar] [CrossRef] [PubMed]
- Sombié, A.; Saiki, E.; Yaméogo, F.; Sakurai, T.; Shirozu, T.; Fukumoto, S.; Sanon, A.; Weetman, D.; McCall, P.; Kanuka, H.; et al. Fréquences élevées des mutations F1534C et V1016I kdr et association avec la résistance aux pyréthroïdes chez Aedes aegypti de Somgandé (Ouagadougou), Burkina Faso. Trop. Med. Santé 2019, 47, 2. [Google Scholar]
- Agbohun, I.K.; Idowu, E.T.; Oyeniyi, T.A.; Adeogun, A.O.; Adesalu, K.; Nwanya, O.; Okonkwo, F.; Oladosu, Y.; Otubanjo, O. First Detection and Co-occurrence of kdr (F1534C and S989P) Mutations in Multiple Insecticides Resistant Aedes aegypti in Nigeria. Preprints 2021, 2021070302. [Google Scholar] [CrossRef]
- Ayres, C.F.J.; Seixas, G.; Borrego, S.; Marques, C.; Monteiro, I.; Marques, C.S.; Gouveia, B.; Leal, S.; Troco, A.D.; Fortes, F.; et al. The V410L knockdown resistance mutation occurs in island and continental populations of Aedes aegypti in West and Central Africa. PLoS Negl. Trop. Dis. 2020, 14, e0008216. [Google Scholar] [CrossRef] [PubMed]
- Fontenille, D.; Traore-Lamizana, M.; Diallo, M.; Thonnon, J.; Digoutte, J.P.; Zeller, H.G. New vectors of Rift Valley fever in West Africa. Emerg. Infect. Dis. 1998, 4, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Guideline for Evaluating Insecticide Resistance in Vectors Using the CDC Bottle Bioassay. Available online: https://stacks.cdc.gov/view/cdc/21777/cdc_21777_DS1.pdf (accessed on 20 January 2022).
- Hemingway, J. Insecticide Resistance Mechanisms (Field and Laboratory Manual); World Health Organization: Geneva, Switzerland, 1998; pp. 1–39. [Google Scholar]
- Rigby, L.M.; Rašić, G.; Peatey, C.L.; Hugo, L.E.; Beebe, N.W.; Devine, G.J. Identifying the fitness costs of a pyrethroid-resistant genotype in the major arboviral vector Aedes Aegypti. Parasites Vectors 2020, 13, 358. [Google Scholar] [CrossRef]
- Zuharah, W.F.; Sufian, M. The discovery of a novel knockdown resistance (kdr) mutation A1007G on Aedes aegypti (Diptera: Culicidae) from Malaysia. Sci. Rep. 2021, 11, 5180. [Google Scholar] [CrossRef] [PubMed]
- AhbiRami, R.; Ishak, I.H.; Yahya, Z.S.; Zuharah, W.F. Knockdown resistance (kdr) in dengue vectors, Aedes aegypti and Aedes albopictus from Malaysia: A post-flood risk assessment. Gen. Mol. Biol. 2020, 19, gmr18604. [Google Scholar]
- Naw, H.; Võ, T.C.; Lê, H.G.; Kang, J.-M.; Mya, Y.Y.; Myint, M.K.; Kim, T.-S.; Shin, H.-J.; Na, B.-K. Knockdown Resistance Mutations in the Voltage-Gated Sodium Channel of Aedes aegypti (Diptera: Culicidae) in Myanmar. Insects 2022, 13, 322. [Google Scholar] [CrossRef]
- Fan, Y.; Scott, J.G. The F1534C voltage-sensitive sodium channel mutation confers 7- to 16-fold resistance to pyrethroid insecticides in Aedes aegypti. Pest Manag. Sci. 2020, 76, 2251–2259. [Google Scholar] [CrossRef] [PubMed]
- Derilus, D.; Impoinvil, L.M.; Muturi, E.J.; McAllister, J.; Kenney, J.; Massey, S.E.; Hemme, R.; Kothera, L.; Lenhart, A. Comparative Transcriptomic Analysis of Insecticide-Resistant Aedes aegypti from Puerto Rico Reveals Insecticide-Specific Patterns of Gene Expression. Genes 2023, 14, 1626. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| kdr Mutation | Study Site | # Mosquitoes Tested | Homozygote Mutation (RR) | Heterozygote Mutation (RS) | Homozygote Wild Type (SS) | Allele Frequency | χ2 Test | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dead | Alive | Dead | Alive | Dead | Alive | R | S | |||||||||
| Dead | Alive | Dead | Alive | Dead | Alive | Dead | Alive | |||||||||
| S989P | 10ème arrondissement of Cotonou | 24 | 0 | 9 | 9 | 3 | 3 | 0 | 0.38 | 0.88 | 0.62 | 0.12 | 4.32 | 0.245 | 0.038 | 0.621 |
| Godomey-Togoudo | 54 | 15 | 21 | 8 | 4 | 6 | 0 | 0.66 | 0.92 | 0.34 | 0.08 | 4.40 | 0.189 | 0.036 | 0.664 | |
| F1534C | 10ème arrondissement of Cotonou | 78 | 33 | 15 | 12 | 9 | 5 | 4 | 0.78 | 0.70 | 0.22 | 0.30 | 4.52 | 1.61 | 0.033 | 0.204 |
| Godomey-Togoudo | 111 | 31 | 23 | 21 | 12 | 17 | 7 | 0.60 | 0.69 | 0.40 | 0.31 | 9.20 | 4.62 | 0.0024 | 0.032 | |
| V1016G | 10ème arrondissement of Cotonou | 33 | 5 | 7 | 5 | 7 | 6 | 3 | 0.47 | 0.62 | 0.53 | 0.38 | 2.22 | 0.279 | 0.136 | 0.597 |
| Godomey-Togoudo | 16 | 1 | 8 | 1 | 3 | 1 | 2 | 0.50 | 0.73 | 0.50 | 0.27 | 0.333 | 2.22 | 0.564 | 0.136 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokponnon, T.F.; Ossè, R.; Zoulkifilou, S.D.; Amos, G.; Festus, H.; Idayath, G.; Sidick, A.; Messenger, L.A.; Akogbeto, M. Insecticide Resistance in Aedes aegypti Mosquitoes: Possible Detection of kdr F1534C, S989P, and V1016G Triple Mutation in Benin, West Africa. Insects 2024, 15, 295. https://doi.org/10.3390/insects15040295
Tokponnon TF, Ossè R, Zoulkifilou SD, Amos G, Festus H, Idayath G, Sidick A, Messenger LA, Akogbeto M. Insecticide Resistance in Aedes aegypti Mosquitoes: Possible Detection of kdr F1534C, S989P, and V1016G Triple Mutation in Benin, West Africa. Insects. 2024; 15(4):295. https://doi.org/10.3390/insects15040295
Chicago/Turabian StyleTokponnon, Tatchémè Filémon, Razaki Ossè, Sare Dabou Zoulkifilou, Gbenouga Amos, Houessinon Festus, Gounou Idayath, Aboubakar Sidick, Louisa A. Messenger, and Martin Akogbeto. 2024. "Insecticide Resistance in Aedes aegypti Mosquitoes: Possible Detection of kdr F1534C, S989P, and V1016G Triple Mutation in Benin, West Africa" Insects 15, no. 4: 295. https://doi.org/10.3390/insects15040295