
Molecular Correlates of Diapause in Aphidoletes aphidimyza
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Effects of Temperature and Photoperiod on Diapause Induction of A. aphidimyza
2.3. Insect Stage Sensitive to Photoperiod Induction of Diapause in A. aphidimyza
2.4. Transcriptomics Analysis and Quantitative Real-Time PCR (qRT-PCR) Validation
2.5. Physiological and Biochemical Substance Determination
2.6. Data Analysis
3. Results
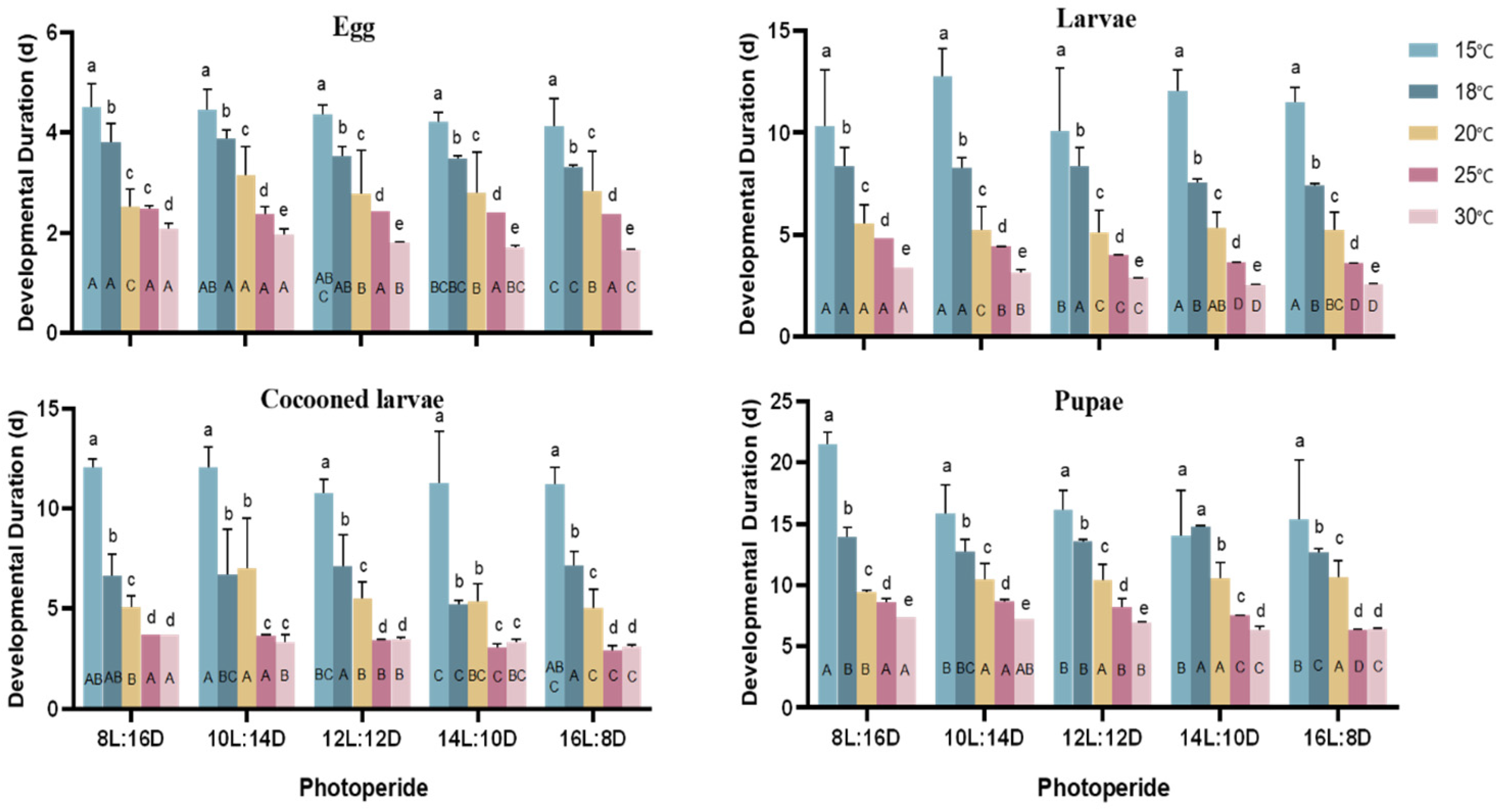
3.1. Effects of Temperature and Photoperiod on the Developmental Periods of A. aphidimyza
3.2. Effects of Temperature and Photoperiod on Diapause in A. aphidimyza
3.3. Photoperiod Sensitive Stage of A. aphidimyza to Diapause Induction
3.4. Transcriptomic Analysis of A. aphidimyza
3.5. Differential Expression Gene Analysis
3.6. GO Enrichment Analysis of Differentially Expressed Genes
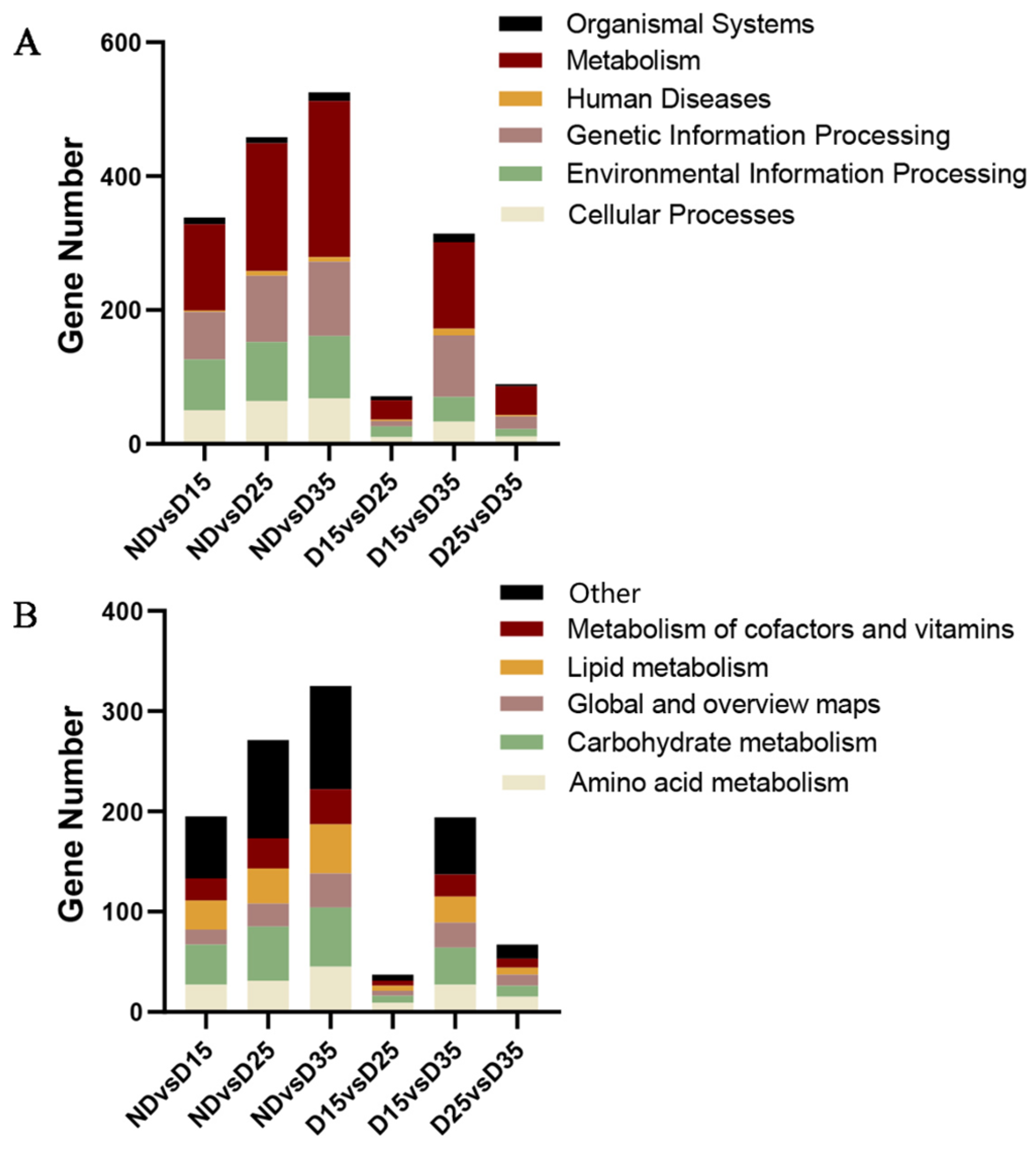
3.7. KEGG Enrichment Analysis of Differentially Expressed Genes
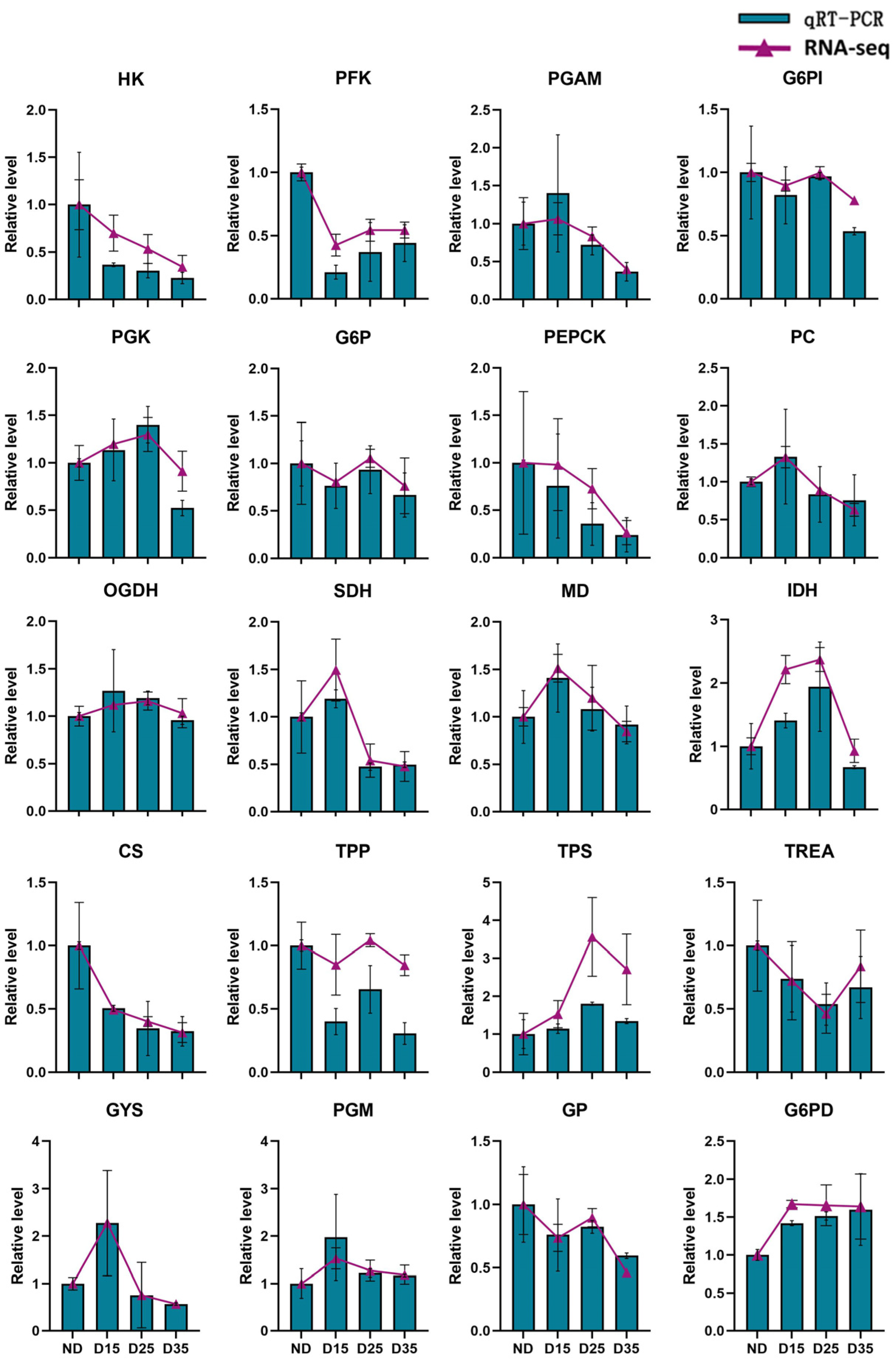
3.8. Real-Time Quantitative PCR Analysis
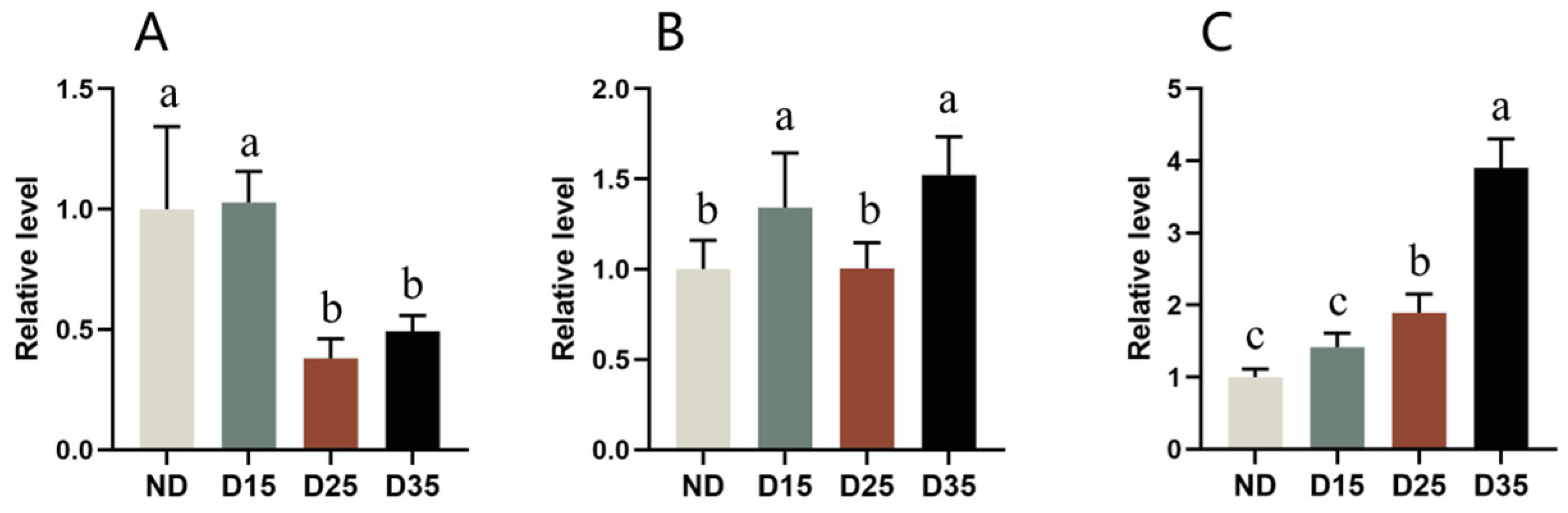
3.9. Water and Fat Contents in A. aphidimyza
3.10. Glycogen, Trehalose, and Sorbitol Contents in A. aphidimyza
4. Discussion
4.1. Effects of Temperature and Photoperiod on the Developmental Duration of A. aphidimyza
4.2. Effects of Temperature and Photoperiod on Diapause Induction of A. aphidimyza
4.3. Life Stage of A. aphidimyza Sensitive to Diapause Induction by Photoperiod
4.4. Transcriptome Sequence and Analysis of Differentially Expressed Genes in the Transcriptome
4.5. Analysis of Differentially Expressed Genes in Glucose Metabolism Pathways
4.6. Changes in Physiological and Biochemical Substances
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lenteren, J.C.V. The state of commercial augmentative biological control: Plenty of natural enemies, but a frustrating lack of uptake. BioControl 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Jandricic, S.E.; Wraight, S.P.; Gillespie, D.R.; Sanderson, J.P. Biological control outcomes using the generalist aphid predator Aphidoletes aphidimyza under multi-prey conditions. Insects 2016, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q. Research progress on Aphidoletes aphidimyza. Anhui Agric. Sci. Bull. 2007, 6, 136–137. [Google Scholar] [CrossRef]
- François-Xavier, B.; Sarah, J.; Karel, B.L.W.F.; Apostolos, P. Optimizing aphid biocontrol with the predator Aphidoletes aphidimyza, based on biology and ecology. Pest Manag. Sci. 2019, 75, 1479–1493. [Google Scholar] [CrossRef]
- Xie, M.; Cheng, H.K.; Qiu, W.L. Application of life table to evaluate the reproduction system of Aphidoletes aphidimyza. Acta Entomol. Sin. 2000, S1, 151–156. [Google Scholar]
- Yano, E. Ecological considerations for biological control of aphids in protected culture. Popul. Ecol. 2006, 48, 333–339. [Google Scholar] [CrossRef]
- Lin, Q.C.; Chen, H.; Yin, Y.Y.; Zhang, S.C.; Yi, Y.; Zhuang, Q.Y.; Zheng, L.; Zhai, Y.F. Effects of temperature on the development and predation of Aphidoletes aphidimyza (Rondani) larvae. Chin. J. Appl. Entomol. 2019, 56, 79–84. [Google Scholar]
- Gong, Y.J.; Shi, B.C.; Lu, H. A study on the optimum cold storage conditions of Aphidoletes aphidimyza. Plant Prot. 2007, 33, 48–50. [Google Scholar]
- Kostal, V.; Sula, J.; Simek, P. Physiology of drought tolerance and cold hardiness of the Mediterranean tiger moth Cymbalophora pudica during summer diapause. J. Insect Physiol. 1998, 44, 165–173. [Google Scholar] [CrossRef]
- Havelka, J.; Zemek, R. Intraspecific variability of aphidophagous gall midge Aphidoletes aphidimyza (Rondani) (Dipt., Cecidomyiidae) and its importance for biological control of aphids. J. Appl. Entomol. 1988, 105, 280–288. [Google Scholar] [CrossRef]
- Havelka, J. Effect of temperature on the developmental rate of preimaginal stages of Aphidoletes aphidimyza (Diptera, Cecidomyiidae). Entomol. Exp. Appl. 1980, 27, 83–90. [Google Scholar] [CrossRef]
- Yamane, M.; Yano, E.; Matsumoto, Y.; Yoshioka, S.; Kawai, T.; Toyonishi, H.; Nakamura, T. Effect of photoperiod and temperature on the induction of diapause in a Japanese strain of Aphidoletes aphidimyza (Diptera: Cecidomyiidae). Appl. Entomol. Zool. 2012, 47, 17–26. [Google Scholar] [CrossRef]
- Yocum, G.D.; Buckner, J.S.; Fatland, C.L. A comparison of internal and external lipids of nondiapausing and diapause initiation phase adult Colorado potato beetles, Leptinotarsa decemlineata. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 159, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Wipking, W.; Viebahn, M.; Neumann, D. Oxygen consumption, water, lipid and glycogen contentof early and late diapause and non-diapause larvae of the burnet moth Zygaena trifolii. J. Insect Physiol. 1995, 41, 47–56. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Wu, G.Q.; Wang, J.; Liu, C.L.; Qiu, L.H. Next-generation sequencing-based transcriptome analysis of Helicoverpa armigera Larvae immune-primed with Photorhabdus luminescens TT01. PLoS ONE 2017, 8, e80146. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.F.; Zhang, Z.M.; Gao, H.H.; Chen, H.; Sun, M.; Zhang, W.; Zheng, L. Hormone signaling regulates nymphal diapause in Laodelphax striatellus (Hemiptera: Delphacidae). Sci. Rep. 2017, 7, 13370. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Song, C.X.; Huang, L.F. Effects of temperature and humidity on the growth and development of Anabremia sp. J. Anhui Agric. Sci. 2019, 47, 113–115. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Zhou, Z.C.; Chen, X.S. Characterization of the complete mitochondrial genome of Epicauta impressicornis (Coleoptera: Meloidae) and Its phylogenetic implications for the infraorder cucujiformia. J. Insect Sci. 2020, 20, 16. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, M.F.; Wang, L.S. Influence of temperature on the development of Aphidoletes aphidimyza. Chin. Bull. Entomol. 2008, 45, 256–259. [Google Scholar] [CrossRef]
- Saunders, D.S. Dormancy, diapause, and the role of the circadian system in insect photoperiodism. Annu. Rev. Entomol. 2020, 65, 373–389. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, S.; Li, N.; Chen, J.; He, Y.Z.; Qin, Q.J. Effects of photoperiods on adult diapause of Harmonia axyridis (Pallas). Chin. J. Plant Prot. 2014, 41, 495–500. [Google Scholar] [CrossRef]
- Xu, Y.Y.; Mou, J.Y.; Hu, C.; Xi, D. The effects of photoperiod and temperature on the reproduction of adults of Chrysoperla sinica Tjeder. Entomol. J. East China 2002, 01, 39–43. [Google Scholar]
- Wang, X.X.; Fan, J.L.; Zhou, M.; Gao, G.; Wei, L.Y.; Kang, L. Interactive effect of photoperiod and temperature on the induction and termination of embryonic diapause in the migratory locust. Pest Manag. Sci. 2021, 77, 2854–2862. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.H. Studies on effects of temperature and phtotoperiod on diapause regulation in Hippodamia variegate (Goeze). Chin. Nanjing Agric. Univ. 2013. [Google Scholar] [CrossRef]
- Gilkeson, L.A. Cold storage of the predatory midge Aphidoletes aphidimyza (Diptera: Cecidomyiidae). J. Econ. Entomol. 1990, 83, 965–970. [Google Scholar] [CrossRef]
- Chen, P.Y.; Feng, H.Q.; Li, G.P.; Qiang, X.S.; Niu, Y.T.; Qiu, F. Preliminary study on variation in diapause rate for different geographic populations of Apolygus lucorum (Meyer-Dur). Chin. J. Plant Prot. 2016, 42, 178–180. [Google Scholar] [CrossRef]
- Feng, H.Q.; Jin, Y.L.; Zhang, Y.F.; Huang, J.R.; Feng, H.; Hou, Y.M. Modelling the combined effects of photoperiod and temperature on diapause induction in Apolygus lucorum (Meyer-Dür) through different latitudes. Pest Manag. Sci. 2020, 77, 2231–2237. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B. Studies on diapause induced by photoperiod and temperature in Hyphantria cunea (Lepidoptera: Arctiidae) and transcriptomics analysis. Henan Univ. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Zhuo, D.G.; Li, Z.H.; Men, X.Y.; Yu, Y.; Zhang, A.S.; Li, L.L. Effects of photoperiod and temperature on diapause induction of Apolygus lucorum Meyer-Dür (Hemiptera: Miridae). Acta Ecol. Sin. 2011, 54, 1082–1086. [Google Scholar] [CrossRef]
- Zhai, Y.F.; Zhang, X.Y.; Yin, Z.J.; Zhu, Q.S.; Tao, M.; Yu, Y.; Zheng, L. Nymphal Diapause in Laodelphax striatellus (Hemiptera: Delphacidae). J. Entomol. Sci. 2018, 53, 107–122. [Google Scholar] [CrossRef]
- Chen, Y.S.; Chen, C.; Liu, X.P.; Kuang, X.J.; Xue, F.S. Comparisons of photosensitivity of diapause during larval development among different geographic populations of Helicoverpa armigera. Acta Agric. Univ. Jiangxiensis 2012, 34, 288–293. [Google Scholar] [CrossRef]
- Mansingh, A.; Smallman, B.N. The influence of temperature on the photoperiodic regulation of diapause in saturniids. J. Insect Physiol. 1971, 17, 1735–1739. [Google Scholar] [CrossRef]
- Shi, C.H. Studies on effects of temperature and photoperiod on diapause regulation in Orius nagaii. Shandong Agric. Univ. 2022. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, Y.X.; Xu, W.H. Proteomic and metabolomic profiles of larval hemolymph associated with diapause in the cotton bollworm, Helicoverpa armigera. BMC Genom. 2013, 14, 751. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.C.; Yang, H.J.; Shen, B.H. Effect of Photoperiod on diapause of Diaphania pyloalis in North District of Zhejiang Province. Acta Sericologica Sin. 2002, 28, 329–332. [Google Scholar] [CrossRef]
- Shi, A.J.; Xu, H.F.; Liu, Z.D.; Zhao, J.; Zhang, F.; Xu, Y.Y. Effect of photoperiod on induction of prepupal diapause and larval developmentin Chrysopa pallens (Rambur) (Neuroptera: Chrysopidae). Acta Ecol. Sin. 2008, 28, 3854–3859. [Google Scholar] [CrossRef]
- Li, W.X.; Li, J.C.; Lu, Z.Y.; Liu, X.X.; Zhang, Q.W. Critical photoperiod and sensitive stage of diapause induction in Microplitis mediator (Haliday) (Hymenoptera: Braconidae). Acta Ecol. Sin. 2008, 51, 635–639. [Google Scholar] [CrossRef]
- Sun, S.H.; Zhao, L.W.; Qi, J.Y. Diapause induction and post-diapause development in Chouioia cunea Yang. Acte Ecol. Sin. 2009, 52, 1307–1311. [Google Scholar] [CrossRef]
- Marteaux, L.D. Regulation of Diapause Entry and Termination in the Swede Midge, Contarinia nasturtii (Diptera: Cecidomyiidae). Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2012. Available online: https://hdl.handle.net/10214/3657 (accessed on 12 December 2023).
- Xu, J.; Jiang, T.; Xue, P.; Shen, G.S.; Huang, J.Y.; Zhu, J. Changes in the mRNA levels and activities of carbohydrate metabolism-related enzymes in the diapause-destined, non-diapause-destined and immediately acid-treated eggs of the bivoltine silkworm (Bombyx mori). Acta Ecol. Sin. 2020, 63, 1431–1440. [Google Scholar] [CrossRef]
- Cheng, W.N.; Li, X.L.; Li, Y.P.; Li, J.J.; Wu, J.X. Activities of four sugar metabolic enzymes in Sitodiplosis mosellana (Gehin) (Diptera: Cecidomyiidae) larvae at different diapause stages. Acta Ecol. Sin. 2009, 52, 133–139. [Google Scholar]
- Xu, W.H.; Lu, Y.X.; Denlinger, D.L. Cross-talk between the fat body and brain regulates insect developmental arrest. Proc. Natl. Acad. Sci. USA 2012, 109, 14687–14692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Han, H.B.; Xu, L.B.; Song, M.X.; Liu, A.P. Transcriptomic analysis of diapause-associated genes in Exorista civilis Rondani (Diptera: Tachinidae). Arch. Insect Biochem. Physiol. 2021, 107, e21789. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Z.; Wang, M.Q.; Xie, Y.Q.; Xiang, M.; Li, P.; Li, Y.Y.; Zhang, L.S. Gene cloning and expression analysis of trehalose-6-phosphate synthase, glycogen synthase and glycogen phosphorylase reveal the glycometabolism in the diapause process of aphidius gifuensis. J. Asia-Pacific Entomol. 2020, 23, 641–645. [Google Scholar] [CrossRef]
- Poelchau, M.F.; Reynolds, J.A.; Denlinger, D.L.; Elsik, C.G.; Armbruster, P.A. A de novo transcriptome of the Asian tiger mosquito, Aedes albopictus, to identify candidate transcripts for diapause preparation. BMC Genom. 2011, 12, 619. [Google Scholar] [CrossRef] [PubMed]
- Ragland, G.J.; Denlinger, D.L.; Hahn, D.A. Mechanisms of suspended animation are revealed by transcript profiling of diapause in the flesh fly. Proc. Natl. Acad. Sci. USA 2010, 107, 14909–14914. [Google Scholar] [CrossRef] [PubMed]
- Besteiro, S.; Barrett, M.P.; Rivière, L.; Bringaud, F. Energy generation in insect stages of Trypanosoma brucei: Metabolism in flux. Trends Parasitol. 2005, 21, 185–191. [Google Scholar] [CrossRef]
- Wu, Z.P. Respiratory & survival dynamics and differential expression of trehalases in diapausing pupae of Pieris melete. Jiangxi Agric. Univ. 2019. [Google Scholar] [CrossRef]
- Zhang, X.; Du, W.; Zhang, J.; Zou, Z.; Ruan, C. High-throughput profiling of diapause regulated genes from Trichogramma dendrolimi (Hymenoptera: Trichogrammatidae). BMC Genom. 2020, 21, 864. [Google Scholar] [CrossRef]
- Nelson, A.; Alfredo, M.V.; Enrique, M.; Gabriel, L. Insights on the evolution of trehalose biosynthesis. BMC Evol. Biol. 2006, 6, 109. [Google Scholar] [CrossRef]
- Pellerone, F.I.; Archer, S.K.; Behm, C.A.; Grant, W.N.; Lacey, M.J.; Somerville, A.C. Trehalose metabolism genes in Caenorhabditis elegans and filarial nematodes. Int. J. Parasitol. 2023, 33, 1195–1206. [Google Scholar] [CrossRef]
- Zhang, Q.; Denlinger, D.L. Dynamics of diapause hormone and prothoracicotropic hormone transcript expression at diapause termination in pupae of the corn earworm, Helicoverpa zea. Peptides 2012, 34, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.Y.; Qi, X.Y.; Jiang, S.; Liu, Y.; Zhang, L.S. Differential expression and function of glucose metabolism regulating enzymes in the diapause Coccinella septempunctata Linnaeus. In Proceedings of the 2014 Annual Conference of the Chinese Plant Protection Society, Xiamen, China, 6–7 November 2014. [Google Scholar]
- Huang, Q.; Zhang, G.; Nan, J.; Cheng, W.; Zhu-Salzman, K. Characterization of trehalose metabolic genes and corresponding enzymatic activities during diapause of Sitodiplosis mosellana. J. Insect Physiol. 2021, 135, 104324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Ma, J. Insect supercooling point and its influence factors. Tianjin Agric. Sci. 2013, 19, 76–84. [Google Scholar] [CrossRef]
- Hou, F.; Ma, J.; Liu, X.; Wang, Y.; Liu, X.N.; Zhang, F.C. Seasonal changes in antifreeze protein gene transcription and water content of beetle Microdera punctipennis (Coleoptera, Tenebrionidae) from Gurbantonggut desert in Central Asia. Cryo Lett. 2010, 31, 359–370. [Google Scholar]
- Colinet, H.; Vernon, P.; Hance, T. Does thermal-related plasticity in size and fat reserves influence supercooling abilities and cold-tolerance in Aphidius colemani (Hymenoptera: Aphidiinae) mummies? J. Therm. Biol. 2007, 32, 374–382. [Google Scholar] [CrossRef]
- Li, X.C.; Cui, J.; Tang, J.W.; Xia, T.T.; Xu, W.; Shi, S.C. Comparison of the supercooling capacity and contents of main biochemical substances between the diapausing and non-diapausing larvae of Leguminivora glycinivorella (Lepidoptera: Olethreutidae). Acta Ecol. Sin. 2019, 62, 1009–1016. [Google Scholar] [CrossRef]
- Guo, T.T.; Ling, F.; Zhang, S.Y.; Yu, Y.; Li, L.L.; Xu, Y.Y. The super-cooling ability and biochemical substances in diapausing and non-diapausing larvae of Grapholitha molesta (Busck) (Lepidoptera: Tortricidae). Chin. J. Appl. Entomol. 2013, 50, 1514–1518. [Google Scholar] [CrossRef]
- Lv, B.; Sun, M.; Zhai, Y.F.; Zheng, L.; Chen, H. Changes of cold tolerance and biochemical substances in diapause and post-diapause Orius sauteri. Chin. J. Biol. Control 2020, 36, 885–890. [Google Scholar] [CrossRef]
- Sinclair; Brent, J.; Marshall, K.E. The many roles of fats in overwintering insects. J. Exp. Biol. 2018, 221, 161836. [Google Scholar] [CrossRef]
- Koczor, S.; Szentkirályi, F.; Tóth, M. New perspectives for simultaneous attraction of Chrysoperla and Chrysopa lacewing species for enhanced biological control (Neuroptera: Chrysopidae). Sci. Rep. 2019, 9, 10303. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilization. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.F.; Zhou, W.Z.; Zhao, S.; Jin, D.C.; Guo, J.J. Changes in the fat, carbohydrate and protein content of Aspongopus chinensis during diapause. Chin. J. Appl. Entomol. 2019, 56, 1019. [Google Scholar]
- Chen, Z.Z.; Zhao, N.; Yin, X.C.; Zhang, F.; Xu, Y.Y. Physiological and biochemical changes in naturally overwintering adults of Chrysoperla sinica (Neuroptera: Chrysopidae) during diapause termination under two different photoperiods. Acta Ecol. Sin. 2013, 56, 120–130. [Google Scholar] [CrossRef]
- Yi, C.H.; Chen, X.M.; Shi, J.Y.; Zhou, C.L. Change of fat contents in the non-diapause and over-winter adult of Kallima inachus. Shandong For. Technol. 2009, 39, 65–66. [Google Scholar] [CrossRef]
- Hedan, L.; Daqing, X.; Xin, T.; Huang, D.Y.; Huang, Y.; Zhao, G.H.; Hu, X.Q.; Wang, X.Y. The role of trehalose biosynthesis on mycolate composition and L-glutamate production in Corynebacterium glutamicum. Microbiol. Res. 2022, 267, 127260. [Google Scholar] [CrossRef]
- Khani, A.; Moharramipour, S.; Barzegar, M. Cold tolerance and trehalose accumulation in overwintering larvae of the codling moth, Cydia pomonella (Lepidoptera: Tortricidae). Eur. J. Entomol. 2007, 104, 385–392. [Google Scholar] [CrossRef]
- Wang, X.Y.; Hou, J.M.; Bo, L.Q.; Wei, D.X.; Ruan, R.L.; Zhang, B.H. Effects of low temperature stress on trehalose and trehalase in muscle larvae of Trichinella spiralis. Chin. Vet. Sci. 2023, 53, 78–84. [Google Scholar] [CrossRef]
- Sun, L.Z.; Wang, G.B.; Zhu, X.W.; Zhang, Y.; Jiang, Y.; Yang, R. Effect of different light treatments on diapause termination and glycometabolism of Antheraea pernyi pupa. Sci. Seric. 2019, 45, 75–80. [Google Scholar] [CrossRef]
- Jiang, H.; Yuan, Z.L.; Wang, W.X.; Liu, Z.L.; Luo, L. Changes in activity of four kinds of metabolic enzymes and three kinds of protective enzymes in female adults of Ceroplastes japonicus before and after overwintering. China Plant Prot. 2018, 38, 11–16. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, H.; Li, X.J.; Wu, J.X.; Cheng, W.N.; Zhu, K.Y. Changes in sorbitol content and the expression level of sorbitol dehydrogenase gene in Sitodiplosis mosellana (Diptera: Cecidomyiidae) larvae at different diapause stages. Acta Ecol. Sin. 2016, 59, 119–126. [Google Scholar] [CrossRef]
- Zhao, L.N.; Zhang, W.X.; Qin, M.; Wang, Y.; Wang, H.F.; Liu, Z.G. Effect of exogenous trehalose and sorbitol on cold resistance of Apis mellifera ligustica. Chin. J. Anim. Nutr. 2021, 33, 1652–1662. [Google Scholar] [CrossRef]
- Zhou, Y.C.; He, H.M.; Xue, F.S. The effects of photoperiod and temperature on diapause induction in the Taian population of cabbage beetle, Colaphellus bowringi Baly. Environ. Entomol. 2018, 40, 123–128. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diapause Standard | Development Duration of Cocooned Larvae |
|---|---|
| non-diapause | ≤5 d |
| incomplete diapause | 6–15 d |
| complete diapause | >15 d |
| Treatment | Different Developmental Stages | ||||
|---|---|---|---|---|---|
| Egg | 1st Instar Larva | 2nd Instar Larva | 3rd Instar Larva | Cocooned Larvae | |
| 1 | S | L | L | L | L |
| 2 | S | S | L | L | L |
| 3 | S | S | S | L | L |
| 4 | S | S | S | S | L |
| 5 | S | S | S | S | S |
| 6 | L | S | S | S | S |
| 7 | L | L | S | S | S |
| 8 | L | L | L | S | S |
| 9 | L | L | L | L | S |
| 10 | L | L | L | L | L |
| Biological Parameters | Source | df | F | p |
|---|---|---|---|---|
| Egg | Temperatures | 4 | 1207.353 | <0.05 |
| Photoperiods | 4 | 18.181 | <0.05 | |
| Temperatures × Photoperiods | 16 | 8.168 | <0.05 | |
| Error | 3811 | |||
| Larvae | Temperatures | 4 | 6648.596 | <0.05 |
| Photoperiods | 4 | 92.349 | <0.05 | |
| Temperatures × Photoperiods | 16 | 28.389 | <0.05 | |
| Error | 3876 | |||
| Cocooned larvae | Temperatures | 4 | 2131.563 | <0.05 |
| Photoperiods | 4 | 26.552 | <0.05 | |
| Temperatures × Photoperiods | 16 | 16.991 | <0.05 | |
| Error | 3721 | |||
| Pupae | Temperatures | 4 | 2089.923 | <0.05 |
| Photoperiods | 4 | 102.040 | <0.05 | |
| Temperatures × Photoperiods | 16 | 45.208 | <0.05 | |
| Error | 3721 |
| Treatment | Diapause Rate (%) |
|---|---|
| 1 | 16.25 ± 5.70 c |
| 2 | 59.38 ± 12.04 b |
| 3 | 83.33 ± 4.41 ab |
| 4 | 100.00 ± 0.00 a |
| 5 | 100.00 ± 0.00 a |
| 6 | 83.25 ± 3.98 ab |
| 7 | 17.84 ± 2.60 c |
| 8 | 16.78 ± 3.51 c |
| 9 | 13.81 ± 6.92 c |
| 10 | 0.00 ± 0.00 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, X.; Wang, Y.; Liu, Y.; Wang, R.; Su, L.; Yin, Z.; Zhao, S.; Chen, H.; Zheng, L.; Dong, X.; et al. Molecular Correlates of Diapause in Aphidoletes aphidimyza. Insects 2024, 15, 299. https://doi.org/10.3390/insects15050299
Dai X, Wang Y, Liu Y, Wang R, Su L, Yin Z, Zhao S, Chen H, Zheng L, Dong X, et al. Molecular Correlates of Diapause in Aphidoletes aphidimyza. Insects. 2024; 15(5):299. https://doi.org/10.3390/insects15050299
Chicago/Turabian StyleDai, Xiaoyan, Yu Wang, Yan Liu, Ruijuan Wang, Long Su, Zhenjuan Yin, Shan Zhao, Hao Chen, Li Zheng, Xiaolin Dong, and et al. 2024. "Molecular Correlates of Diapause in Aphidoletes aphidimyza" Insects 15, no. 5: 299. https://doi.org/10.3390/insects15050299