Variations in Mating and Reproduction in Oriental Fruit Moth Caused by Adult Physiological State in Laboratory Tests

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Experimental Designs Applicable to All Experiments
2.2.1. Exp. #1. Mating Selection
2.2.2. Exp. #2. Anatomical and Morphological Observations of Reproductive System
2.2.3. Exp. #3. Mating and Reproductive Traits
2.3. Statistical Analysis
3. Results
3.1. Effect of Adult Physiological States on Selection of One Sex for the Opposite Sex
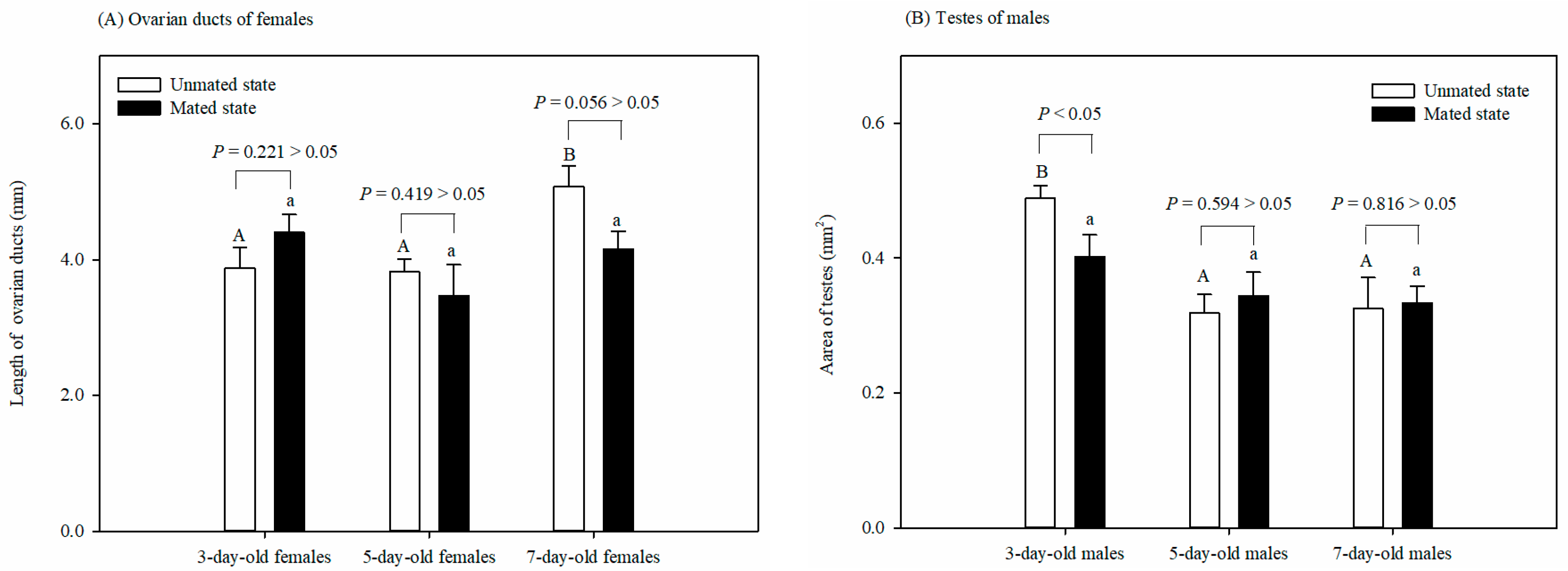
3.2. Effect of Adult Physiological States on Reproductive System
3.3. Effect of Adult Physiological States on Mating and Reproductive Success
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, Z.G.; Li, Y.L.; He, X.H.; Rong, E.H.; Zhang, L.J.; Wang, M.L.; Ma, R.Y. Research on the voltinism of oriental fruit moths based on GIS. J. Shanxi Agr. Univ. 2019, 39, 81–86. [Google Scholar] [CrossRef]
- Du, J.; Li, G.; Xu, X.; Wu, J. Development and fecundity performance of oriental fruit moth (Lepidoptera: Tortricidae) reared on shoots and fruits of peach and pear in different seasons. Environ. Entomol. 2015, 44, 1522–1530. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Liu, Z.F.; Guo, Y.F.; Zhao, F.; Fan, R.J. The effect of different hosts on the oviposition preferences of Grapholita molesta (Busck) (Lepidoptera: Tortricidae). Chin. J. Appl. Entomol. 2018, 55, 104–111. [Google Scholar]
- Kanga, L.H.B.; Pree, D.J.; van Lier, J.L.; Whitty, K.J. Mechanisms of resistance to organophosphorus and carbamate insecticides in oriental fruit moth populations (Grapholita molesta Busck). Pestic. Biochem. Phys. 1997, 59, 11–23. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Chai, Y.P.; Zhang, L.J.; Zhao, Z.G.; Gao, L.L.; Ma, R.Y. Transcriptome analysis and identification of major detoxification gene families and insecticide targets in Grapholita molesta (Busck) (Lepidoptera: Tortricidae). J. Insect. Sci. 2017, 17, 43. [Google Scholar] [CrossRef]
- Cui, G.Z.; Zhu, J.J. Pheromone-based pest management in China: Past, present, and future prospects. J. Chem. Ecol. 2016, 42, 557–570. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Sanchez-González, Á.; Ponce-Escudero, F.; Martín-Vertedor, D.; Ferrero-García, J.J. Assessing mass trapping efficiency and population density of Cerambyx welensii Küster by mark-recapture in dehesa open woodlands. Eur. J. For. Res. 2012, 131, 1103–1116. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Jia, X.T.; Gao, Y.; Fan, R.J.; Li, J.; Ma, R.Y. Emergence and mating behavior of the Oriental fruit moth Cydia molesta (Lepidoptera: Tortricidae) and its potential for reproduction. Ann. Soc. Entomol. 2019, 55, 446–453. [Google Scholar] [CrossRef]
- Sadek, M.M. Polyandry in field-collected Spodoptera littoralis moths and laboratory assessment of the effects of male mating history. Entomol. Exp. Appl. 2001, 98, 165–172. [Google Scholar] [CrossRef]
- Blyth, J.E.; Gilburn, A.S. Extreme promiscuity in a mating system dominated by sexual conflict. J. Insect Behav. 2006, 19, 447–455. [Google Scholar] [CrossRef]
- Parker, G.A. Sexual conflict over mating and fertilization: An overview. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 235–259. [Google Scholar] [CrossRef] [PubMed]
- Thornhill, R.; Gwynne, D.T. The evolution of sexual differences in insects. Am. Sci. 1986, 74, 382–389. [Google Scholar]
- Candolin, U. Reproduction under predation risk and the trade-off between current and future reproduction in the three-spine stickleback. Proc. R. Soc. London. Ser. B Biol. Sci. 1998, 265, 1171–1175. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Guo, Y.F.; Chai, X.H.; Li, J.; Ma, R.Y. Effects of operational sex ratio, mating age, and male mating history on mating and reproductive behavior in Grapholita molesta. Bull. Entomol. Res. 2021, 111, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Zhang, X.H.; Zhang, J.L.; Huang, B.J.; Jian, C.Z.; Peng, X.; Vreysen, M.J.B.; Chen, M.H. flight performance, fecundity, and ovary development of Grapholita molesta (Lepidoptera: Torticidae) at different ages. Insects 2022, 13, 837. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Men, L.N.; Peng, Y.F.; Li, J.; Deng, A.; Chen, Y.; Liu, X.Q.; Ma, R.Y. Morphological differences of the reproductive system could be used to predict the optimum Grapholita molesta (Busck) control period. Sci. Rep. 2017, 7, e8198. [Google Scholar] [CrossRef]
- Du, J.; Wang, Y.R.; Wu, J.X. Effect of four different artificial diets on development and reproduction of Grapholita molesta (Lepidoptera: Tortricidae). J. Shanxi Agr. Univ. 2010, 30, 229–231. [Google Scholar] [CrossRef]
- Kong, W.N.; Wang, Y.; Guo, Y.F.; Chai, X.H.; Li, J.; Ma, R.Y. Importance of preovipositional period of an oligophagous moth in predicting host suitability. J. Econ. Entomol. 2020, 113, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Jia, X.T.; Xiang, H.M.; Diao, H.L.; Yan, Y.; Wang, Y.; Ma, R.Y. The effect of photoperiods and light intensity on mating behavior and reproduction of Grapholita molesta (Lepidoptera: Tortricidae). Environ. Entomol. 2019, 48, 1035–1041. [Google Scholar] [CrossRef]
- Cao, Y.; Benelli, G.; Germinara, G.S.; Germinara, G.S.; Maggi, F.; Zhang, Y.J.; Luo, S.L.; Yang, H.; Li, C. Innate positive chemotaxis to paeonal from highly attractive Chinese medicinal herbs in the cigarette beetle, Lasioderma serricorne. Sci. Rep. 2019, 9, 6995. [Google Scholar] [CrossRef]
- Cao, Y.; Pistillo, O.M.; Lou, Y.B.; D’Isita, I.; Maggi, F.; Hu, Q.Q.; Germinara, G.S.; Li, C. Electrophysiological and behavioural responses of Stegobium paniceum to volatile compounds from Chinese medicinal plant materials. Pest. Manag. Sci. 2022, 78, 3697–3703. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R.; Baker, T.C.; Roelofs, W.L. Hairpencil pheromone components of male oriental fruit moths, Grapholitha molesta. J. Chem. Ecol. 1982, 8, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Kuang, X.Q.; Calvin, D.; Knapp, M.; Poston, F. Female European corn borer (Lepidoptera: Crambidae) ovarian developmental stages: Their association with oviposition and use in a classification system. J. Econ. Entomol. 2004, 97, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, H.; Song, J.H.; Li, R.G.; Zhang, Y.B.; Yang, W.J. Function of vitellogenin receptor gene TaVgR in the regulation of reproductive development in Tuta absoluta (Lepidoptera: Gelechiidae). Acta Entomol. Sin. 2022, 65, 675–683. [Google Scholar] [CrossRef]
- Force, E.; Couzi, P.; Dacher, M.; Debernard, S. Diet impacts the reproductive system’s maturation in the male moth Agrotis ipsilon (Noctuidae, Lepidoptera). J. Insect Physiol. 2023, 148, 104532. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.S.; Zaky Shoaib, A.A.; Saad Elabasy, A.S.; Cheng, X.L.; Zhang, Q.; Shi, Z.H. Effects of delayed mating on male mating success and female reproductive performance of Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae). Crop Prot. 2020, 132, 105135. [Google Scholar] [CrossRef]
- Gunathilaka, R.A.K.M.; Jayatunga, D.P.W.; Ganehiarachchi, G.A.S.M. Effect of delayed mating on reproductive performance and life-history parameters of dengue vector Aedes aegypti. Bull. Entomol. Res. 2023, 113, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? Annu. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Dufour, M.C.; Anton, S. Transient post-mating inhibition of behavioural and central nervous responses to sex pheromone in an insect. Proc. R. Soc. London. Ser. B Biol. Sci. 2001, 268, 1631–1635. [Google Scholar] [CrossRef]
- Shukla, S.; Shilpa, M.C.; Gadagkar, R. Virgin wasps develop ovaries on par with mated females, but lay fewer eggs. Insect. Soc. 2013, 60, 345–350. [Google Scholar] [CrossRef]
- Perez-Mendoza, J.; Throne, J.E.; Baker, J.E. Ovarian physiology and age-grading in the rice weevil, Sitophilus oryzae (Coleoptera: Curculionidae). J. Stored Prod. Res. 2004, 40, 179–196. [Google Scholar] [CrossRef]
- Shahid, M.; Siddiqui, A.; Mishra, G. Mating alters the rate of development of ovarioles in the ladybird, Propylea dissecta (Coleoptera: Coccinellidae). Eur. J. Entomol. 2016, 113, 44–50. [Google Scholar] [CrossRef]
- Li, W.J.; Yuan, W.; Zhao, X.C.; Li, Y.H.; Wu, K.M. Effects of mating on ovarian development and oviposition of Apolygus lucorum. J. Asia-Pac. Entomol. 2017, 20, 1442–1446. [Google Scholar] [CrossRef]
- Liu, F.; Liao, H.J.; Shi, L.; Du, T.; Zho, C.L.; Deng, J.; Ma, Y.Q. Morphology and development of reproductive system of female adult Tirumala limniace (Cramer). For. Res. 2017, 30, 125–130. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; Hu, T.Y.; Yang, H.L.; Wei, J.H.; Jin, L.; Hao, D.J. Morphology and development of the internal reproductive system of Monochamus alternatus (Coleoptera: Cerambycidae) adults. Acta Entomol. Sin. 2023, 66, 1210–1220. [Google Scholar] [CrossRef]
- Muzzi, M.; Di Giulio, A.; Mancini, E.; Fratini, E.; Cervelli, M.; Gasperi, T.; Mariottini, P.; Persichini, T.; Bologna, M.A. The male reproductive accessory glands of the blister beetle Meloe proscarabaeus Linnaeus, 1758 (Coleoptera: Meloidae): Anatomy and ultrastructure of the cantharidin-storing organs. Arthropod Struct. 2020, 59, 100980. [Google Scholar] [CrossRef]
- Klein, C.D.; Kozii, I.V.; Wood, S.C.; Koziy, R.V.; Zabrodski, M.W.; Dvylyuk, I.; de Mattos, I.M.; Moshynskyy, I.; Honaramooz, A.; Simko, E. Testicular changes of honey bee drones, Apis mellifera (Hymenoptera: Apidae), during sexual maturation. J. Insect Sci. 2021, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Makee, H.; Saour, G. Factors influencing mating success, mating frequency, and fecundity in Phthorimaea operculella (Lepidoptera: Gelechiidae). Environ. Entomol. 2001, 30, 31–36. [Google Scholar] [CrossRef]
- Dunn, D.W.; Sumner, J.P.; Goulson, D. The benefits of multiple mating to female seaweed flies, Coelopa frigida (Diptera: Coelpidae). Behav. Ecol. Sociobiol. 2005, 58, 128–135. [Google Scholar] [CrossRef]
- Colares, F.; Michaud, J.P.; Torres, J.B.; Silva-Torres, C.S.A. Polyandry and male mating history affect the reproductive performance of Eriopis connexa (Coleoptera: Coccinellidae). Ann. Entomol. Soc. Am. 2015, 108, 736–742. [Google Scholar] [CrossRef]
- Fraser, H.W.; Trimble, R.M. Effect of delayed mating on reproductive biology of the Oriental fruit moth (Lepidoptera: Tortricidae). Can. Entomol. 2001, 133, 219–227. [Google Scholar] [CrossRef]
- de Morais, R.M.D.; Redaelli, L.R.; Sant’Ana, J. Age and multiple mating effects on reproductive success of Grapholita molesta (Busck) (Lepidoptera, Tortricidae). Rev. Bras. Entomol. 2012, 56, 319–324. [Google Scholar] [CrossRef]
- de Morais, M.; Sant’Ana, J.; Redaelli, L.R.; Lorscheiter, R. Effects of aging and polygamy on the reproductive performance of Grapholita molesta (Lepidoptera: Tortricidae). Rev. Colomb. Entomol. 2011, 37, 67–70. Available online: http://hdl.handle.net/10183/108524 (accessed on 30 May 2024). [CrossRef]


{kind=link}
{kind=link}
| Unmated Female | Male | Number of Matings per Cage | Duration of Matings per Cage (min) | Number of Eggs Laid by a Female per Cage | Percentage of Eggs Hatched per Cage (%) | |
|---|---|---|---|---|---|---|
| 3-day-old | 3-day-old | Mated | 2.40 ± 0.45 ab | 63.20 ± 11.90 a | 135.80 ± 18.28 d | 61.73 ± 10.81 ab |
| Unmated | 1.10 ± 0.10 ab | 26.60 ± 3.93 a | 37.70 ± 14.95 abc | 81.02 ± 10.30 ab | ||
| 5-day-old | Mated | 1.00 ± 0.19 a | 31.50 ± 5.45 a | 91.25 ± 10.16 bcd | 82.74 ± 3.67 ab | |
| Unmated | 1.60 ± 0.40 ab | 55.30 ± 12.92 a | 93.00 ± 25.47 abcd | 95.95 ± 2.46 ab | ||
| 7-day-old | Mated | 1.33 ± 0.37 ab | 33.11 ± 8.20 a | 125.44 ± 18.88 d | 79.03 ± 7.79 ab | |
| Unmated | 1.10 ± 0.10 ab | 34.70 ± 2.26 a | 128.30 ± 21.51 d | 96.48 ± 1.59 b | ||
| 5-day-old | 3-day-old | Mated | 2.50 ± 0.56 b | 68.40 ± 15.49 a | 134.90 ± 14.37 d | 78.32 ± 8.94 ab |
| Unmated | 1.30 ± 0.15 ab | 35.50 ± 4.27 a | 124.90 ± 10.54 d | 98.00 ± 0.79 b | ||
| 5-day-old | Mated | 1.80 ± 0.36 ab | 50.60 ± 9.85 a | 144.40 ± 20.60 d | 84.76 ± 5.10 b | |
| Unmated | 1.00 ± 0.00 ab | 35.00 ± 4.69 a | 25.67 ± 11.14 ab | 76.63 ± 11.73 ab | ||
| 7-day-old | Mated | 1.20 ± 0.13 ab | 39.70 ± 4.00 a | 114.10 ± 18.45 cd | 83.81 ± 6.85 ab | |
| Unmated | 1.50 ± 0.27 ab | 46.60 ± 6.56 a | 107.80 ± 19.42 cd | 97.16 ± 1.20 b | ||
| 7-day-old | 3-day-old | Mated | 1.30 ± 0.15 ab | 36.80 ± 5.61 a | 100.80 ± 13.67 cd | 62.29 ± 11.88 ab |
| Unmated | 1.10 ± 0.10 ab | 30.90 ± 2.96 a | 94.00 ± 17.55 bcd | 86.90 ± 7.40 b | ||
| 5-day-old | Mated | 1.60 ± 0.31 ab | 47.30 ± 11.81 a | 69.10 ± 19.91 abcd | 49.96 ± 14.00 a | |
| Unmated | 1.00 ± 0.00 ab | 35.90 ± 4.07 a | 68.10 ± 18.62 abcd | 81.41 ± 8.79 ab | ||
| 7-day-old | Mated | 1.17 ± 0.40 ab | 34.67 ± 13.33 a | 121.00 ± 22.86 d | 76.21 ± 8.38 ab | |
| Unmated | 1.00 ± 0.00 ab | 42.33 ± 6.68 a | 13.00 ± 4.18 a | 77.56 ± 7.90 ab | ||
| Mated Female | Male | Number of Matings per Cage | Duration of Matings per Cage (min) | Number of Eggs Laid by a Female per Cage after Pairing | Percentage of Eggs Hatched per Cage after Pairing (%) | Total Number of Eggs Laid by a Female per Cage | Total Percentage of Eggs Hatched per Cage (%) | |
|---|---|---|---|---|---|---|---|---|
| 3-day-old | 3-day-old | Mated | 0.50 ± 0.31 a | 12.90 ± 7.16 a | 141.10 ± 12.31 def | 94.38 ± 1.04 c | 148.20 ± 14.22 a | 94.08 ± 1.10 b |
| Unmated | 0.50 ± 0.17 a | 15.80 ± 6.27 a | 170.00 ± 14.54 ef | 91.25 ± 1.39 bc | 170.60 ± 14.71 a | 90.99 ± 1.45 b | ||
| 5-day-old | Mated | 0.30 ± 0.15 a | 11.60 ± 6.57 a | 171.00 ± 12.75 f | 92.15 ± 1.89 bc | 175.10 ± 12.61 a | 89.87 ± 1.74 b | |
| Unmated | 0.50 ± 0.17 a | 22.90 ± 8.31 a | 170.70 ± 13.35 f | 71.09 ± 10.43 abc | 170.80 ± 13.34 a | 71.10 ± 10.43 ab | ||
| 7-day-old | Mated | 0.22 ± 0.15 a | 6.33 ± 4.26 a | 129.56 ± 19.79 def | 88.31 ± 5.85 bc | 134.44 ± 20.10 a | 87.96 ± 5.40 b | |
| Unmated | 0.50 ± 0.17 a | 18.50 ± 6.27 a | 131.90 ± 9.77 def | 91.73 ± 2.53 bc | 134.00 ± 9.61 a | 90.15 ± 2.47 b | ||
| 5-day-old | 3-day-old | Mated | 0.10 ± 0.10 a | 3.70 ± 3.70 a | 89.90 ± 14.29 bcd | 90.53 ± 2.34 bc | 119.20 ± 14.98 a | 75.67 ± 6.86 ab |
| Unmated | 0.20 ± 0.13 a | 5.90 ± 3.93 a | 94.70 ± 9.75 bcde | 88.24 ± 3.19 bc | 119.90 ± 12.44 a | 87.98 ± 2.46 b | ||
| 5-day-old | Mated | 1.20 ± 0.47 a | 52.70 ± 30.46 a | 120.90 ± 7.64 def | 81.58 ± 6.91 abc | 161.60 ± 11.62 a | 82.06 ± 5.41 b | |
| Unmated | 0.10 ± 0.10 a | 6.00 ± 6.00 a | 107.30 ± 9.81 cdef | 89.44 ± 3.36 bc | 135.80 ± 10.24 a | 87.67 ± 3.38 b | ||
| 7-day-old | Mated | 0.40 ± 0.22 a | 15.10 ± 8.29 a | 89.90 ± 14.38 abcd | 62.50 ± 11.15 a | 118.40 ± 18.80 a | 59.81 ± 10.13 a | |
| Unmated | 0.20 ± 0.13 a | 8.00 ± 5.54 a | 93.10 ± 4.81 bcde | 94.19 ± 1.52 c | 118.40 ± 8.92 a | 88.27 ± 4.04 b | ||
| 7-day-old | 3-day-old | Mated | 0.80 ± 0.25 a | 29.90 ± 9.73 a | 65.00 ± 13.21 abc | 77.20 ± 8.80 abc | 135.10 ± 10.87 a | 83.15 ± 9.29 ab |
| Unmated | 0.40 ± 0.16 a | 13.00 ± 5.43 a | 52.20 ± 7.40 ab | 68.59 ± 10.75 ab | 135.50 ± 20.85 a | 72.01 ± 10.20 ab | ||
| 5-day-old | Mated | 0.90 ± 0.43 a | 19.30 ± 8.85 a | 85.10 ± 12.08 abcd | 66.80 ± 8.29 abc | 151.80 ± 15.53 a | 67.78 ± 8.39 ab | |
| Unmated | 0.40 ± 0.16 a | 16.90 ± 8.03 a | 37.10 ± 6.50 a | 58.81 ± 9.88 a | 114.30 ± 21.22 a | 53.96 ± 9.60 a | ||
| 7-day-old | Mated | 0.80 ± 0.29 a | 25.90 ± 9.27 a | 103.80 ± 10.64 bcdef | 91.52 ± 1.01 bc | 157.90 ± 12.12 a | 92.03 ± 1.36 b | |
| Unmated | 0.44 ± 0.18 a | 20.00 ± 8.36 a | 85.78 ± 9.22 bcd | 80.02 ± 9.67 abc | 152.89 ± 12.87 a | 81.10 ± 9.56 ab | ||
| Unmated Male | Female | Number of Matings per Cage | Duration of Matings per Cage (min) | Number of Eggs Laid by a Female per Cage after Pairing | Percentage of Eggs Hatched per Cage after Pairing (%) | Total Number of Eggs Laid by a Female per Cage | Total Percentage of Eggs Hatched per Cage (%) | |
|---|---|---|---|---|---|---|---|---|
| 3-day-old | 3-day-old | Mated | 0.50 ± 0.17 ab | 15.80 ± 6.27 abcd | 170.00 ± 14.54 g | 91.25 ± 1.39 a | 170.60 ± 14.71 e | 90.99 ± 1.45 a |
| Unmated | 1.10 ± 0.10 cd | 26.60 ± 3.93 bcdef | 37.70 ± 14.95 abc | 81.02 ± 10.30 a | 37.70 ± 14.95 abc | 81.02 ± 10.30 a | ||
| 5-day-old | Mated | 0.20 ± 0.13 a | 5.90 ± 3.93 ab | 94.70 ± 9.75 defg | 88.24 ± 3.19 a | 119.90 ± 12.44 de | 87.98 ± 2.46 a | |
| Unmated | 1.30 ± 0.15 d | 35.50 ± 4.27 cdef | 124.90 ± 10.54 efg | 98.00 ± 0.79 a | 124.90 ± 10.54 de | 98.00 ± 0.79 a | ||
| 7-day-old | Mated | 0.40 ± 0.16 a | 13.00 ± 5.43 abc | 52.20 ± 7.40 abcde | 68.59 ± 10.75 a | 135.50 ± 20.85 de | 72.01 ± 10.20 a | |
| Unmated | 1.10 ± 0.10 cd | 30.90 ± 2.96 cdef | 94.00 ± 17.55 cdefg | 86.90 ± 7.40 a | 94.00 ± 17.55 cde | 86.90 ± 7.40 a | ||
| 5-day-old | 3-day-old | Mated | 0.50 ± 0.17 ab | 22.90 ± 8.31 abcdef | 170.70 ± 13.35 g | 71.09 ± 10.43 a | 170.80 ± 13.34 e | 71.10 ± 10.43 a |
| Unmated | 1.60 ± 0.40 d | 55.30 ± 12.92 f | 93.00 ± 25.47 bcdef | 95.95 ± 2.46 a | 93.00 ± 25.47 bcde | 95.95 ± 2.46 a | ||
| 5-day-old | Mated | 0.10 ± 0.10 a | 6.00 ± 6.00 a | 107.30 ± 9.81 efg | 89.44 ± 3.36 a | 135.80 ± 10.24 de | 87.67 ± 3.38 a | |
| Unmated | 1.00 ± 0.00 abc | 35.00 ± 4.69 cdef | 25.67 ± 11.14 ab | 76.63 ± 11.73 a | 25.67 ± 11.14 ab | 76.63 ± 11.73 a | ||
| 7-day-old | Mated | 0.40 ± 0.16 a | 16.90 ± 8.03 abcd | 37.10 ± 6.50 abcd | 58.81 ± 9.88 a | 114.30 ± 21.22 de | 53.96 ± 9.60 a | |
| Unmated | 1.00 ± 0.00 abc | 35.90 ± 4.07 cdef | 68.10 ± 18.62 bcdef | 81.41 ± 8.79 a | 68.10 ± 18.62 abcd | 81.41 ± 8.79 a | ||
| 7-day-old | 3-day-old | Mated | 0.50 ± 0.17 ab | 18.50 ± 6.27 abcde | 131.90 ± 9.77 fg | 91.73 ± 2.53 a | 134.00 ± 9.61 de | 90.15 ± 2.47 a |
| Unmated | 1.10 ± 0.10 cd | 34.70 ± 2.26 cdef | 128.30 ± 21.51 efg | 96.48 ± 1.59 a | 128.30 ± 21.51 de | 96.48 ± 1.59 a | ||
| 5-day-old | Mated | 0.20 ± 0.13 a | 8.00 ± 5.54 ab | 93.10 ± 4.81 defg | 94.19 ± 1.52 a | 118.40 ± 8.92 de | 88.27 ± 4.04 a | |
| Unmated | 1.50 ± 0.27 d | 46.60 ± 6.5 6 ef | 107.80 ± 19.42 defg | 97.16 ± 1.20 a | 107.80 ± 19.42 de | 97.16 ± 1.20 a | ||
| 7-day-old | Mated | 0.44 ± 0.18 ab | 20.00 ± 8.36 abcde | 85.78 ± 9.22 cdefg | 80.02 ± 9.67 a | 152.89 ± 12.87 e | 81.10 ± 9.56 a | |
| Unmated | 1.00 ± 0.00 abc | 42.33 ± 6.68 def | 13.00 ± 4.18 a | 77.56 ± 7.90 a | 13.00 ± 4.18 a | 77.56 ± 7.90 a | ||
| Mated Male | Female | Number of Matings per Cage | Duration of Matings per Cage (min) | Number of Eggs Laid by a Female per Cage after Pairing | Percentage of Eggs Hatched per Cage after Pairing (%) | Total Number of Eggs Laid by a Female per Cage | Total Percentage of Eggs Hatched per Cage (%) | |
|---|---|---|---|---|---|---|---|---|
| 3-day-old | 3-day-old | Mated | 0.50 ± 0.31 abcd | 12.90 ± 7.16 abcd | 141.10 ± 12.31 bc | 94.38 ± 1.04 b | 148.20 ± 14.22 b | 94.08 ± 1.10 b |
| Unmated | 2.40 ± 0.45 ef | 63.20 ± 11.90 ef | 135.80 ± 18.28 abc | 61.73 ± 10.81 ab | 135.80 ± 18.28 ab | 61.73 ± 10.81 ab | ||
| 5-day-old | Mated | 0.10 ± 0.10 a | 3.70 ± 3.70 a | 89.90 ± 14.29 abc | 90.53 ± 2.34 b | 119.20 ± 14.98 ab | 75.67 ± 6.86 ab | |
| Unmated | 2.50 ± 0.56 f | 68.40 ± 15.49 f | 134.90 ± 14.37 abc | 78.32 ± 8.94 ab | 134.90 ± 14.37 b | 78.32 ± 8.94 ab | ||
| 7-day-old | Mated | 0.80 ± 0.25 abcdef | 29.90 ± 9.73 abcdef | 65.00 ± 13.21 ab | 77.20 ± 8.80 ab | 135.10 ± 10.87 b | 83.15 ± 9.29 ab | |
| Unmated | 1.30 ± 0.15 cdef | 36.80 ± 5.61 bcdef | 100.80 ± 13.67 abc | 62.29 ± 11.88 ab | 100.80 ± 13.67 ab | 62.29 ± 11.88 ab | ||
| 5-day-old | 3-day-old | Mated | 0.30 ± 0.15 abc | 11.60 ± 6.57 abc | 171.00 ± 12.75 c | 92.15 ± 1.89 b | 175.10 ± 12.61 b | 89.87 ± 1.74 b |
| Unmated | 1.00 ± 0.19 abcdef | 31.50 ± 5.45 abcdef | 91.25 ± 10.16 abc | 82.74 ± 3.67 b | 91.25 ± 10.16 ab | 82.74 ± 3.67 b | ||
| 5-day-old | Mated | 1.20 ± 0.47 abcdef | 52.70 ± 30.46 abcdef | 120.90 ± 7.64 abc | 81.58 ± 6.91 b | 161.60 ± 11.62 b | 82.06 ± 5.41 b | |
| Unmated | 1.80 ± 0.36 ef | 50.60 ± 9.85 def | 144.40 ± 20.60 bc | 84.76 ± 5.10 b | 144.40 ± 20.60 b | 84.76 ± 5.10 b | ||
| 7-day-old | Mated | 0.90 ± 0.43 abcde | 19.30 ± 8.85 abcde | 85.10 ± 12.08 abc | 66.80 ± 8.29 ab | 151.80 ± 15.53 b | 67.78 ± 8.39 ab | |
| Unmated | 1.60 ± 0.31 def | 47.30 ± 11.81 cdef | 69.10 ± 19.91 a | 49.96 ± 14.00 a | 69.10 ± 19.91 a | 49.96 ± 14.00 a | ||
| 7-day-old | 3-day-old | Mated | 0.22 ± 0.15 ab | 6.33 ± 4.26 ab | 129.56 ± 19.79 abc | 88.31 ± 5.85 b | 134.44 ± 20.10 ab | 87.96 ± 5.40 b |
| Unmated | 1.33 ± 0.37 bcdef | 33.11 ± 8.20 abcdef | 125.44 ± 18.88 abc | 79.03 ± 7.79 b | 125.44 ± 18.88 ab | 79.03 ± 7.79 b | ||
| 5-day-old | Mated | 0.40 ± 0.22 abc | 15.10 ± 8.29 abcd | 89.90 ± 14.38 abc | 62.50 ± 11.15 ab | 118.40 ± 18.80 ab | 59.81 ± 10.13 ab | |
| Unmated | 1.20 ± 0.13 cdef | 39.70 ± 4.00 cdef | 114.10 ± 18.45 abc | 83.81 ± 6.85 b | 114.10 ± 18.45 ab | 83.81 ± 6.85 b | ||
| 7-day-old | Mated | 0.80 ± 0.29 abcde | 25.90 ± 9.27 abcdef | 103.80 ± 10.64 abc | 91.52 ± 1.01 b | 157.90 ± 12.12 b | 92.03 ± 1.36 b | |
| Unmated | 1.17 ± 0.40 abcdef | 34.67 ± 13.33 abcdef | 121.00 ± 22.86 abc | 76.21 ± 8.38 ab | 121.00 ± 22.86 ab | 76.21 ± 8.38 ab | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, W.; Wang, Y.; Li, N.; Cao, W.; Hu, X.; Liu, C.; Niu, G.; Li, J.; Ma, R. Variations in Mating and Reproduction in Oriental Fruit Moth Caused by Adult Physiological State in Laboratory Tests. Insects 2024, 15, 457. https://doi.org/10.3390/insects15060457
Kong W, Wang Y, Li N, Cao W, Hu X, Liu C, Niu G, Li J, Ma R. Variations in Mating and Reproduction in Oriental Fruit Moth Caused by Adult Physiological State in Laboratory Tests. Insects. 2024; 15(6):457. https://doi.org/10.3390/insects15060457
Chicago/Turabian StyleKong, Weina, Yi Wang, Na Li, Weiye Cao, Xuefeng Hu, Changnian Liu, Guofei Niu, Jie Li, and Ruiyan Ma. 2024. "Variations in Mating and Reproduction in Oriental Fruit Moth Caused by Adult Physiological State in Laboratory Tests" Insects 15, no. 6: 457. https://doi.org/10.3390/insects15060457
APA StyleKong, W., Wang, Y., Li, N., Cao, W., Hu, X., Liu, C., Niu, G., Li, J., & Ma, R. (2024). Variations in Mating and Reproduction in Oriental Fruit Moth Caused by Adult Physiological State in Laboratory Tests. Insects, 15(6), 457. https://doi.org/10.3390/insects15060457