

Human–Environment Interactions Shape Mosquito Seasonal Population Dynamics
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Mosquito Sampling
2.3. Biological Control
2.4. Statistical Analysis
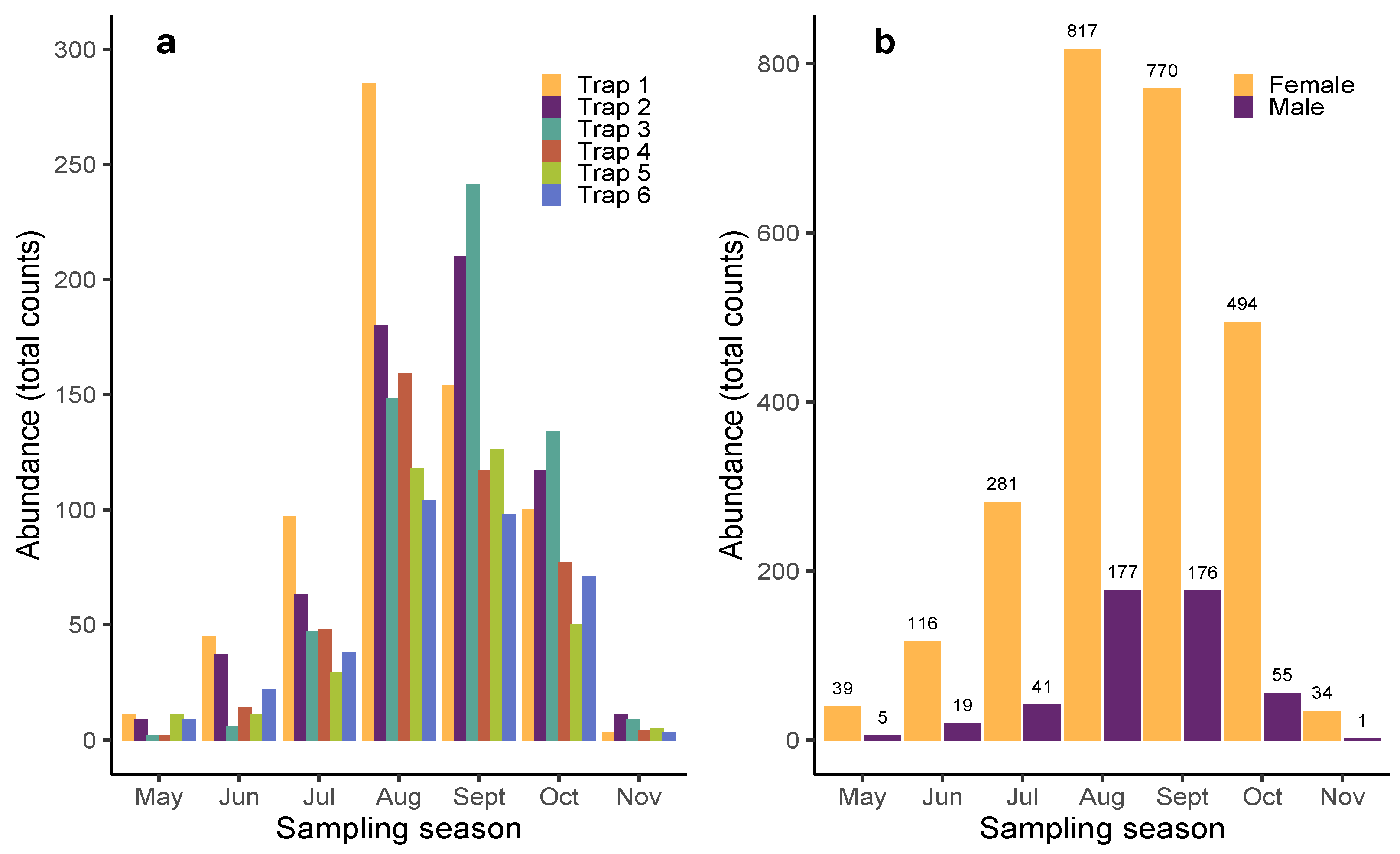
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, T.; Braack, L.; Guarido, M.; Venter, M.; Gouveia Almeida, A.P. Mosquito community composition and abundance at contrasting sites in northern South Africa, 2014–2017. J. Vector Ecol. 2020, 45, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Mayer, S.V.; Tesh, R.B.; Vasilakis, N. The emergence of arthropod-borne viral diseases: A global prospective on dengue, chikungunya and Zika fevers. Acta Trop. 2017, 166, 155–163. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Brady, O.J.; Golding, N.; Pigott, D.M.; Kraemer, M.U.; Messina, J.P.; Reiner, R.C., Jr.; Scott, T.W.; Smith, D.L.; Gething, P.W.; Hay, S.I. Global temperature constraints on Aedes aegypti and Aedes albopictus persistence and competence for dengue virus transmission. Parasit. Vectors 2014, 7, 338. [Google Scholar] [CrossRef]
- Liu-Helmersson, J.; Quam, M.; Wilder-Smith, A.; Stenlund, H.; Ebi, K.; Massad, E.; Rocklöv, J. Climate change and Aedes Vectors: 21st century projections for dengue transmission in Europe. EBioMedicine 2016, 7, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Sousa, A.R.; Ceretti-Júnior, W.; de Carvalho, G.C.; Nardi, M.S.; Araujo, A.B.; Vendrami, D.P.; Marrelli, M.T. Diversity and abundance of mosquitoes (Diptera: Culicidae) in an urban park: Larval habitats and temporal variation. Acta Trop. 2015, 150, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Chowell, G.; Mizumoto, K.; Banda, J.M.; Poccia, S.; Perrings, C. Assessing the potential impact of vector-borne disease transmission following heavy rainfall events: A mathematical framework. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180272. [Google Scholar] [CrossRef]
- Monaghan, A.J.; Morin, C.W.; Steinhoff, D.F.; Wilhelmi, O.; Hayden, M.; Quattrochi, D.A.; Reiskind, M.; Lloyd, A.L.; Smith, K.; Schmidt, C.A.; et al. On the Seasonal Occurrence and Abundance of the Zika Virus Vector Mosquito Aedes Aegypti in the Contiguous United States. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef]
- Caldwell, J.M.; LaBeaud, A.D.; Lambin, E.F.; Stewart-Ibarra, A.M.; Ndenga, B.A.; Mutuku, F.M.; Krystosik, A.R.; Ayala, E.B.; Anyamba, A.; Borbor-Cordova, M.J.; et al. Climate predicts geographic and temporal variation in mosquito-borne disease dynamics on two continents. Nat. Commun. 2021, 12, 1233. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The global compendium of Aedes aegypti and Aedes albopictus occurrence. Sci. Data 2015, 2, e08347. [Google Scholar] [CrossRef]
- Delatte, H.; Toty, C.; Boyer, S.; Bouetard, A.; Bastien, F.; Fontenille, D. Evidence of habitat structuring Aedes albopictus populations in Réunion Island. PLoS Negl. Trop. Dis. 2013, 7, e2111. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kamara, F.; Zhou, G.; Puthiyakunnon, S.; Li, C.; Liu, Y.; Zhou, Y.; Yao, L.; Yan, G.; Chen, X.G. Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship. PLoS Negl. Trop. Dis. 2014, 8, e3301. [Google Scholar] [CrossRef] [PubMed]
- Kolimenakis, A.; Heinz, S.; Wilson, M.L.; Winkler, V.; Yakob, L.; Michaelakis, A.; Papachristos, D.; Richardson, C.; Horstick, O. The role of urbanisation in the spread of Aedes mosquitoes and the diseases they transmit—A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0009631. [Google Scholar] [CrossRef] [PubMed]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control Assoc. Suppl. 1988, 1, 1–39. [Google Scholar] [PubMed]
- Petrić, M.; Ducheyne, E.; Gossner, C.M.; Marsboom, C.; Nicolas, G.; Venail, R.; Hendrickx, G.; Schaffner, F. Seasonality and timing of peak abundance of Aedes albopictus in Europe: Implications to public and animal health. Geospat. Health 2021, 16, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Dieng, H.; Rahman, G.S.; Abu Hassan, A.; Che Salmah, M.; Satho, T.; Miake, F.; Boots, M.; Sazaly, A. The effects of simulated rainfall on immature population dynamics of Aedes albopictus and female oviposition. Int. J. Biometeorol. 2012, 56, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Guzzetta, G.; Montarsi, F.; Baldacchino, F.A.; Metz, M.; Capelli, G.; Rizzoli, A.; Pugliese, A.; Rosà, R.; Poletti, P.; Merler, S. Potential risk of dengue and chikungunya outbreaks in northern Italy based on a population model of Aedes albopictus (Diptera: Culicidae). PLoS Negl. Trop. Dis. 2016, 10, e0004762. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, M.; Martínez-de La Puente, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effects of landscape anthropization on mosquito community composition and abundance. Sci. Rep. 2016, 6, 29002. [Google Scholar] [CrossRef]
- de Souza, W.M.; Weaver, S.C. Effects of climate change and human activities on vector-borne diseases. Nat. Rev. Microbiol. 2024. [Google Scholar] [CrossRef]
- Camargo, C.; Alfonso-Parra, C.; Díaz, S.; Rincon, D.F.; Ramírez-Sánchez, L.F.; Agudelo, J.; Barrientos, L.M.; Villa-Arias, S.; Avila, F.W. Spatial and temporal population dynamics of male and female Aedes albopictus at a local scale in Medellín, Colombia. Parasit. Vectors 2021, 14, 312. [Google Scholar] [CrossRef]
- Roiz, D.; Rosa, R.; Arnoldi, D.; Rizzoli, A. Effects of temperature and rainfall on the activity and dynamics of host-seeking Aedes albopictus females in northern Italy. Vector Borne Zoonotic Dis. 2010, 10, 811–816. [Google Scholar] [CrossRef]
- Tran, A.; L’ambert, G.; Lacour, G.; Benoît, R.; Demarchi, M.; Cros, M.; Cailly, P.; Aubry-Kientz, M.; Balenghien, T.; Ezanno, P. A rainfall-and temperature-driven abundance model for Aedes albopictus populations. Int. J. Environ. Res. Public Health 2013, 10, 1698–1719. [Google Scholar] [CrossRef]
- Erguler, K.; Smith-Unna, S.E.; Waldock, J.; Proestos, Y.; Christophides, G.K.; Lelieveld, J.; Parham, P.E. Large-scale modelling of the environmentally-driven population dynamics of temperate Aedes albopictus (Skuse). PLoS ONE 2016, 11, e0149282. [Google Scholar] [CrossRef]
- Manica, M.; Filipponi, F.; D’Alessandro, A.; Screti, A.; Neteler, M.; Rosà, R.; Solimini, A.; della Torre, A.; Caputo, B. Spatial and Temporal Hot Spots of Aedes albopictus Abundance inside and outside a South European Metropolitan Area. PLoS Negl. Trop. Dis. 2016, 10, e0004758. [Google Scholar] [CrossRef]
- Ravasi, D.; Monton, D.P.; Tanadini, M.; Campana, V.; Flacio, E. Efficacy of biological larvicide VectoMax® FG against Aedes albopictus and Culex pipiens under field conditions in urban catch basins. J. Eur. Mosq. Control Assoc. 2022, 1, 10. [Google Scholar] [CrossRef]
- Lacey, L.A. Bacillus thuringiensis serovariety israelensis and Bacillus sphaericus for mosquito control. J. Am. Mosq. Control Assoc. 2007, 23, 133–163. [Google Scholar] [CrossRef]
- Roiz, D.; Boussès, P.; Simard, F.; Paupy, C.; Fontenille, D. Autochthonous chikungunya Transmission and Extreme Climate Events in Southern France. PLoS Negl. Trop. Dis. 2015, 9, e0003854. [Google Scholar] [CrossRef]
- Nowosad, J. pollen: Analysis of Aerobiological Data. R Package Version 0.71. 2019. Available online: https://CRAN.R-project.org/package=pollen (accessed on 10 July 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level /Mixed) Regression Models. R Package Version 0.4.6. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 10 July 2024).
- QGIS Development Team. QGIS Geographic Information System; QGIS Association: Zurich, Switzerland, 2023. [Google Scholar]
- Medlock, J.; Hansford, K.; Versteirt, V.; Cull, B.; Kampen, H.; Fontenille, D.; Hendrickx, G.; Zeller, H.; Van Bortel, W.; Schaffner, F. An entomological review of invasive mosquitoes in Europe. Bull. Entom. Res. 2015, 105, 637–663. [Google Scholar] [CrossRef]
- Benelli, G.; Wilke, A.B.; Beier, J.C. Aedes albopictus (Asian tiger mosquito). Trends Parasitol. 2020, 36, 942–943. [Google Scholar] [CrossRef]
- WHO. A Global Brief on Vector-Borne Diseases; Technical report; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef]
- Rocklöv, J.; Dubrow, R. Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 479–483. [Google Scholar] [CrossRef]
- Collantes, F.; Delacour, S.; Alarcón-Elbal, P.M.; Ruiz-Arrondo, I.; Delgado, J.A.; Torrell-Sorio, A.; Bengoa, M.; Eritja, R.; Ángel Miranda, M.; Molina, R.; et al. Review of ten-years presence of Aedes albopictus in Spain 2004–2014: Known distribution and public health concerns. Parasit. Vectors 2015, 8, 655. [Google Scholar] [CrossRef]
- Lührsen, D.S.; Zavitsanou, E.; Cerecedo-Iglesias, C.; Pardo-Araujo, M.; Palmer, J.R.; Bartumeus, F.; Montalvo, T.; Michaelakis, A.; Lowe, R. Adult Aedes albopictus in winter: Implications for mosquito surveillance in Southern Europe. Lancet Planet Health 2023, 7, e729–e731. [Google Scholar] [CrossRef]
- Alto, B.W.; Juliano, S.A. Precipitation and temperature effects on populations of Aedes albopictus (Diptera: Culicidae): Implications for range expansion. J. Med. Entomol. 2001, 38, 646–656. [Google Scholar] [CrossRef]
- Delatte, H.; Gimonneau, G.; Triboire, A.; Fontenille, D. Influence of temperature on immature development, survival, longevity, fecundity, and gonotrophic cycles of Aedes albopictus, vector of chikungunya and dengue in the Indian Ocean. J. Med. Entomol. 2009, 46, 33–41. [Google Scholar] [CrossRef]
- Calado, D.C.; Silva, M.A.N.d. Evaluation of the temperature influence on the development of Aedes albopictus. Rev. Saude Publica 2002, 36, 173–179. [Google Scholar] [CrossRef]
- Richards, S.L.; Apperson, C.S.; Ghosh, S.K.; Cheshire, H.M.; Zeichner, B.C. Spatial analysis of Aedes albopictus (Diptera: Culicidae) oviposition in suburban neighborhoods of a Piedmont community in North Carolina. J. Med. Entomol. 2006, 43, 976–989. [Google Scholar] [CrossRef]
- Lourenço-de Oliveira, R.; Castro, M.G.; Braks, M.A.; Lounibos, L.P. The invasion of urban forest by dengue vectors in Rio de Janeiro. J. Vector Ecol. 2004, 29, 94–100. [Google Scholar]
- Wilke, A.B.B.; Medeiros-Sousa, A.R.; Ceretti-Junior, W.; Marrelli, M.T. Mosquito populations dynamics associated with climate variations. Acta Trop. 2017, 166, 343–350. [Google Scholar] [CrossRef]
- Luciano, T.; Severini, I.F.; Di Luca, I.M.; Bella, I.A.; ryP Roberto, R. Seasonal patterns of oviposition and egg hatching rate of Aedes albopictus in Rome. J. Am. Mosq. Control Assoc. 2003, 19, 100. [Google Scholar]
- Higa, Y.; Toma, T.; Araki, Y.; Onodera, I.; Miyagi, I. Seasonal changes in oviposition activity, hatching and embryonation rates of eggs of Aedes albopictus (Diptera: Culicidae) on three islands of the Ryukyu Archipelago, Japan. Med. Entomol. Zool. 2007, 58, 1–10. [Google Scholar] [CrossRef]
- Kache, P.A.; Eastwood, G.; Collins-Palmer, K.; Katz, M.; Falco, R.C.; Bajwa, W.I.; Armstrong, P.M.; Andreadis, T.G.; Diuk-Wasser, M.A. Environmental determinants of Aedes albopictus abundance at a northern limit of its range in the United States. Am. J. Trop. Med. Hyg. 2020, 102, 436. [Google Scholar] [CrossRef]
- Carrieri, M.; Albieri, A.; Angelini, P.; Soracase, M.; Dottori, M.; Antolini, G.; Bellini, R. Effects of the Weather on the Seasonal Population Trend of Aedes albopictus (Diptera: Culicidae) in Northern Italy. Insects 2023, 14, 879. [Google Scholar] [CrossRef]
- Montalvo, T.; Higueros, A.; Valsecchi, A.; Realp, E.; Vila, C.; Ortiz, A.; Peracho, V.; Figuerola, J. Effectiveness of the Modification of Sewers to Reduce the Reproduction of Culex pipiens and Aedes albopictus in Barcelona, Spain. Pathogens 2022, 11, 423. [Google Scholar] [CrossRef]
- Guidi, V.; Lüthy, P.; Tonolla, M. Comparison between diflubenzuron and a Bacillus thuringiensis israelensis–and Lysinibacillus sphaericus–based formulation for the control of mosquito larvae in urban catch basins in Switzerland. J. Am. Mosq. Control Assoc. 2013, 29, 138–145. [Google Scholar] [CrossRef]
- Souza, R.L.; Mugabe, V.A.; Paploski, I.A.D.; Rodrigues, M.S.; Moreira, P.S.d.S.; Nascimento, L.C.J.; Roundy, C.M.; Weaver, S.C.; Reis, M.G.; Kitron, U.; et al. Effect of an intervention in storm drains to prevent Aedes aegypti reproduction in Salvador, Brazil. Parasit. Vectors 2017, 10, 328. [Google Scholar] [CrossRef]
- Marini, F.; Caputo, B.; Pombi, M.; Tarsitani, G.; Torre, A.D. Study of Aedes albopictus dispersal in Rome, Italy, using sticky traps in mark-release-recapture experiments. Med. Vet. Entomol. 2010, 24, 361–368. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Response Variable | Explanatory Variables | Coefficient (±SE) | (95% CI) | IRR (95% CI) | Wald Test | df | p-Value | |
|---|---|---|---|---|---|---|---|---|
| Abundance of | Intercept | 2.78 ± 0.06 | (2.66, 2.91) | 16.25 (14.33, 18.43) | 43.392 | 1882.87 | 1 | <0.0001 |
| mosquitoes | MaxRH | 0.23 ± 0.05 | (0.13, 0.32) | 1.25 (1.14, 1.38) | 4.74 | 22.54 | 1 | <0.0001 |
| MinTemp—21 | 0.58 ± 0.06 | (0.47, 0.70) | 1.80 (1.60, 2.01) | 10.04 | 100.87 | 1 | <0.0001 | |
| N of Visitors | 0.23 ± 0.05 | (0.13, 0.34) | 1.26 (1.14, 1.41) | 4.43 | 19.62 | 1 | <0.0001 | |
| Number of water drains with water | 0.27 ± 0.06 | (0.15, 0.39) | 1.31 (1.16, 1.48) | 4.37 | 19.13 | 1 | <0.0001 | |
| Weeks since treatments * Acc. Rainfall 3w | 0.07 ± 0.03 | (0.003, 0.14) | 1.07 (1.00, 1.15) | 2.05 | 4.23 | 1 | 0.038 | |
| Acc. Rainfall 3w | 0.35 ± 0.05 | (0.25, 0.45) | 1.42 (1.28, 1.57) | 6.90 | 47.67 | 1 | <0.0001 | |
| Weeks since treatments | −0.13 ± 0.05 | (−0.24, −0.029) | 0.87 (0.78, 0.97) | −2.49 | 6.23 | 1 | 0.012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco-Sierra, L.; Bellver-Arnau, J.; Escartin, S.; Mariani, S.; Bartumeus, F. Human–Environment Interactions Shape Mosquito Seasonal Population Dynamics. Insects 2024, 15, 527. https://doi.org/10.3390/insects15070527
Blanco-Sierra L, Bellver-Arnau J, Escartin S, Mariani S, Bartumeus F. Human–Environment Interactions Shape Mosquito Seasonal Population Dynamics. Insects. 2024; 15(7):527. https://doi.org/10.3390/insects15070527
Chicago/Turabian StyleBlanco-Sierra, Laura, Jesús Bellver-Arnau, Santi Escartin, Simone Mariani, and Frederic Bartumeus. 2024. "Human–Environment Interactions Shape Mosquito Seasonal Population Dynamics" Insects 15, no. 7: 527. https://doi.org/10.3390/insects15070527