
New COI-COII mtDNA Region Haplotypes in the Endemic Honey Bees Apis mellifera intermissa and Apis mellifera sahariensis (Hymenoptera: Apidae) in Algeria
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
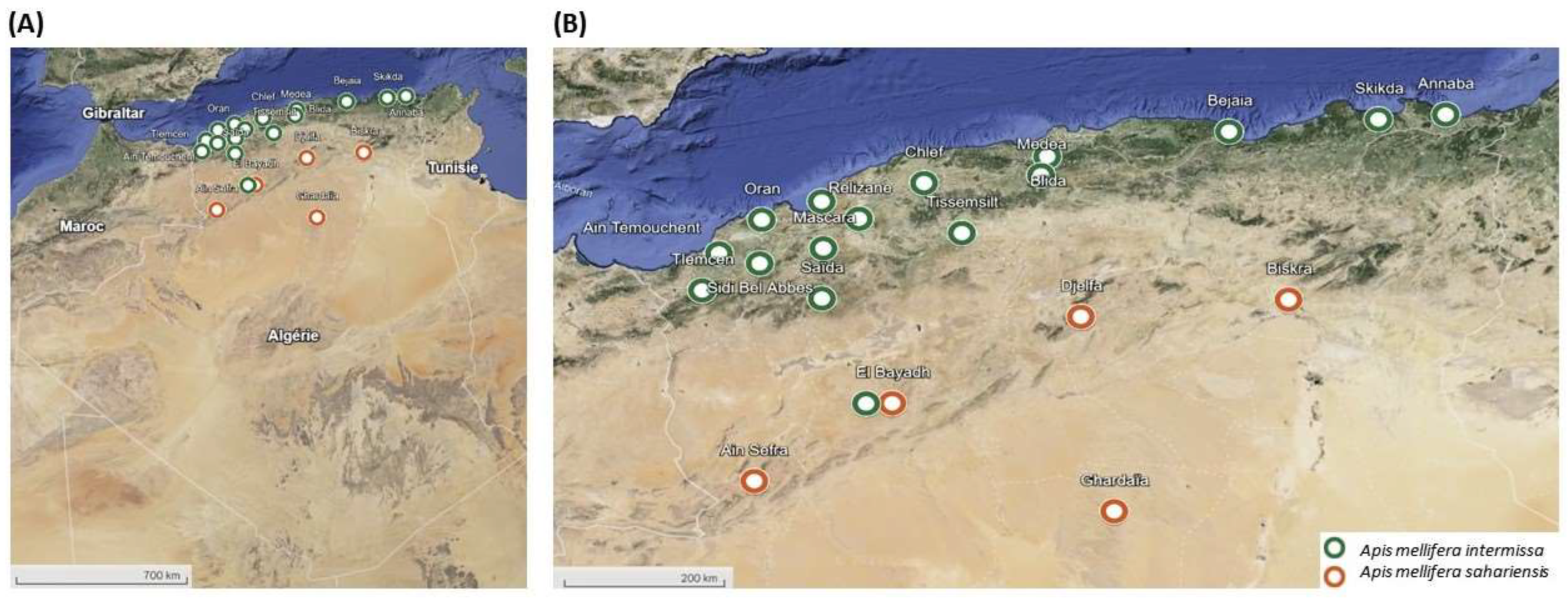
2.1. Honey Bee Sampling
2.2. DNA Extraction
2.3. Mitochondrial DNA Analysis
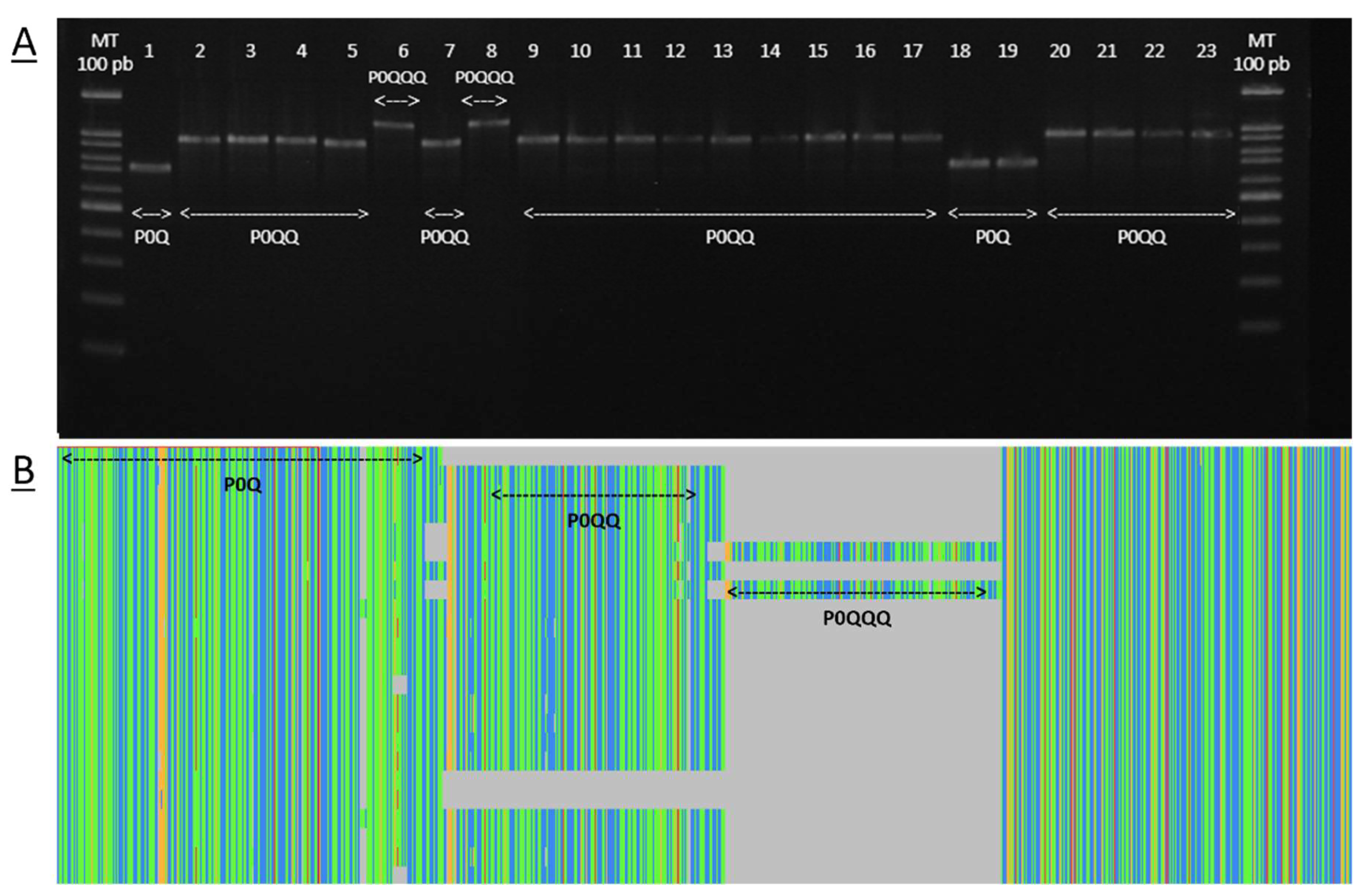
2.4. Nomenclature of Novel Haplotypes
2.5. Phylogenetic Analyses
3. Results
3.1. Honey Bee Evolutionary Lineage and Haplotype Identification
3.2. COI-COII Haplotypes’ Distribution and Comparison
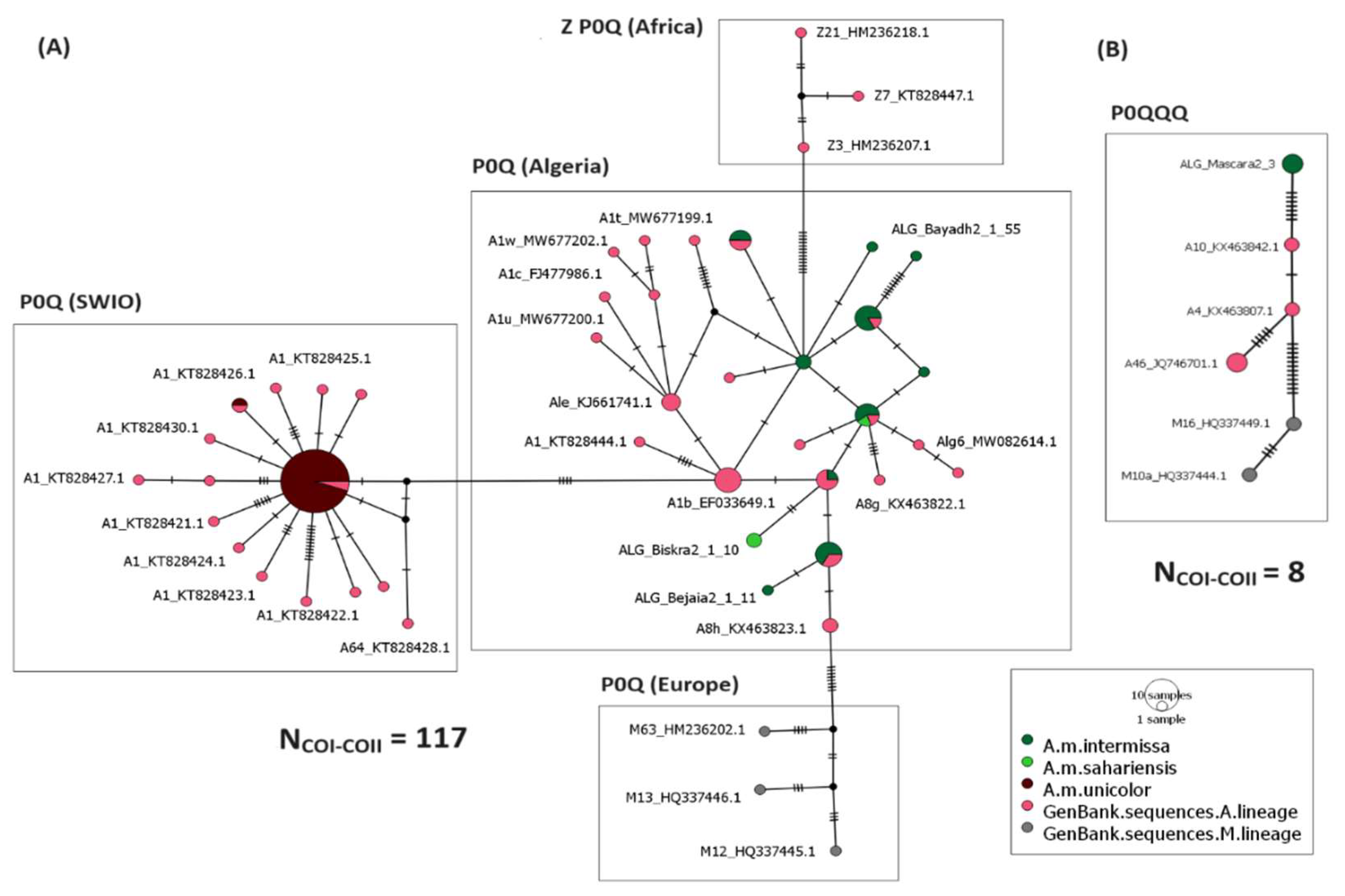
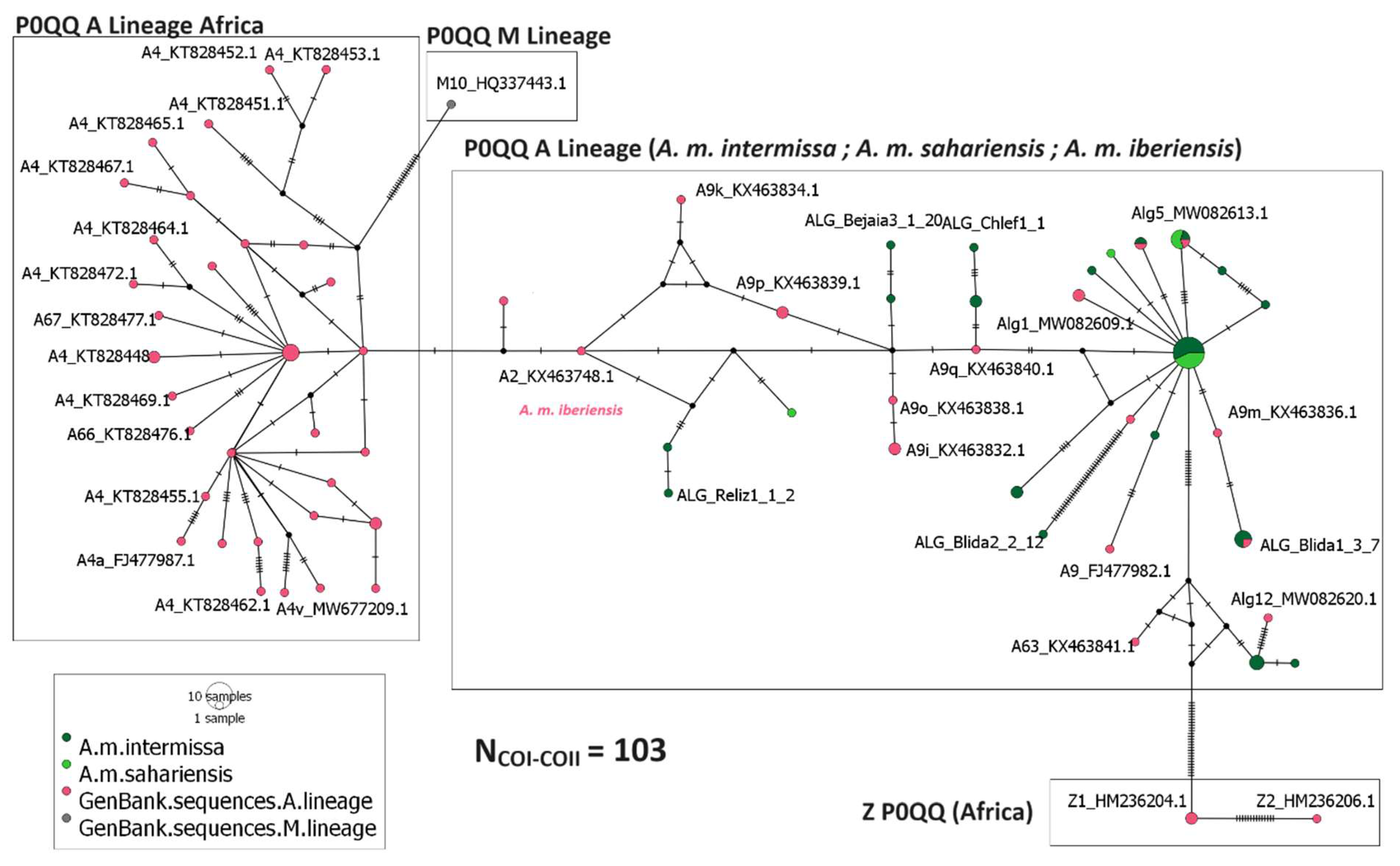
3.3. Evolutionary Lineages Distribution and Relationships among Haplotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.S. The Taxonomy of Recent and Fossil Honey Bees (Hymenoptera: Apidae; Apis). J. Hymenopt. Res. 1999, 8, 165–196. [Google Scholar]
- Ruttner, F. Biogeography and Taxonomy of Honey Bees; Springer: Berlin, Germany, 1988. [Google Scholar]
- Cridland, J.M.; Tsutsui, N.D.; Ramírez, S.R. The Complex Demographic History and Evolutionary Origin of the Western Honey Bee, Apis Mellifera. Genome Biol. Evol. 2017, 9, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Franck, P.; Garnery, L.; Loiseau, A.; Oldroyd, B.P.; Hepburn, H.R.; Solignac, M.; Cornuet, J.M. Genetic diversity of the honey bee in Africa: Microsatellite and mitochondrial data. Heredity 2001, 86, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Chahbar, N.; Muñoz, I.; Dall’Olio, R.; De la Rúa, P.; Serrano, J.; Doumandji, S. Population Structure of North African Honey Bees Is Influenced by Both Biological and Anthropogenic Factors. J. Insect Conserv. 2013, 17, 385–392. [Google Scholar] [CrossRef]
- Chavez-Galarza, J.; Garnery, L.; Henriques, D.; Neves, C.J.; Loucif-Ayad, W.; Jonhston, J.S.; Pinto, M.A. Mitochondrial DNA variation of Apis mellifera iberiensis: Further insights from a large-scale study using sequence data of the tRNAleu-cox2 intergenic region. Apidologie 2017, 48, 533–544. [Google Scholar] [CrossRef]
- Henriques, D.; Chavez-Galarza, J.; Quaresma, A.; Neves, C.J.; Lopes, A.R.; Costa, C.; Costa, F.O.; Rufino, J.; Pinto, M.A. From the popular tRNAleu-COX2 intergenic region to the mitogenome: Insights from diverse honey bee populations of Europe and North Africa. Apidologie 2019, 50, 215–229. [Google Scholar] [CrossRef]
- Buttel-Reepen, H. Contributions to the systematics biology as well as the historical and geographical distribution of honey bees (Apis mellifica L.) their variability and other Apis species. Mitt Zool Mus Berl 1906, 3, 117–201. [Google Scholar]
- Baldensperger, J. North African Bees.—II. Bee World 1923, 5, 189–190. [Google Scholar] [CrossRef]
- Barour, C.; Ali, T.; Radloff, S.; Randall, H. Multivariate analysis of honey bees, Apis mellifera Linnaeus (Hymenoptera: Apidae) of the northeastern and southern regions of Algeria. Afr. Entomol. 2005, 13, 17–23. [Google Scholar]
- Haddad, N.J.; Adjlane, N.; Saini, D.; Menon, A.; Krishnamurthy, V.; Jonklaas, D. Whole-genome sequencing of north African honey bee Apis mellifera intermissa to assess its benefcial traits. Entomol Res. 2018, 48, 174–186. [Google Scholar] [CrossRef]
- Haccour, P. Recherche sur la race d’abeille saharienne au Maroc. Comptes Rendus Société Sci. Nat. Phys. Maroc. 1960, 6, 96–98. [Google Scholar]
- Adjlane, N.; Doumandji, S.E.; Haddad, N. Beekeeping in Algeria: The factors threatening the survival of the honey bee Apis mellifera intermissa. Cah. Agric. 2012, 21, 235–241. [Google Scholar] [CrossRef]
- Mallek, S.; Ladjali, K.; Mohammedi, A. Les Menaces Naturelles et Anthropiques Mettant en Péril l’Existence de l’Abeille Saharienne en Algérie. Bull. Société D‘Histoire Nat. D‘Afrique Nord USTHB 2018, 74, 165–196. [Google Scholar]
- Avise, J.C.; Arnold, J.; Ball, R.M.; Bermingham, E.; Lamb, T.; Neigel, J.E.; Reeb, C.A.; Saunders, N.C. Intraspecific Phylogeography: The Mitochondrial DNA Bridge Between Population Genetics and Systematics. Annu. Rev. Ecol. Syst. 1987, 18, 489–522. [Google Scholar] [CrossRef]
- Garnery, L.; Cornuet, J.M.; Solignac, M. Evolutionary history of the honey bee Apis mellifera inferred from mitochondrial DNA analysis. Mol. Ecol. 1992, 1, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Hall, H.G.; Smith, D.R. Distinguishing African and European honey bee matrilines using amplified mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1991, 88, 4548–4552. [Google Scholar] [CrossRef]
- Alburaki, M.; Moulin, S.; Legout, H.; Alburaki, A.; Garnery, L. Mitochondrial structure of Eastern honey bee populations from Syria, Lebanon and Iraq. Apidologie 2011, 42, 628–641. [Google Scholar] [CrossRef]
- Loucif-Ayad, W.; Achou, M.; Legout, H.; Alburaki, M.; Garnery, L. Genetic Assessment of Algerian Honey bee Populations by Microsatellite Markers. Apidologie 2015, 46, 392–402. [Google Scholar] [CrossRef]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.M. A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Meixner, M.D.; Pinto, M.A.; Bouga, M.; Kryger, P.; Ivanova, E.; Fuchs, S. Standard methods for characterizing subspecies and ecotypes of Apis mellifera. J. Apic. Res. 2013, 52, 1–27. [Google Scholar] [CrossRef]
- El-Niweiri, M.A.A.; Moritz, R.F.A. Mitochondrial discrimination of honeybees (Apis mellifera) of Sudan. Apidologie 2008, 39, 566–573. [Google Scholar] [CrossRef]
- Legesse, G.; Getu, E. Geographical and population diversity of African honey bees, Apis mellifera L. (Hymenoptera: Apidae): A review. SINET Ethiop. J. Sci. 2022, 45, 235–248. [Google Scholar] [CrossRef]
- Fridi, R.; Tabet Aoul, N.; Catays, G.; Basso, B.; Bienefeld, K.; Gregorc, A.; Vignal, A.; Canale-Tabet, K. Genetic diversity and population genetic structure analysis of Apis mellifera subspecies in Algeria and Europe based on complementary sex determiner (CSD) gene. Apidologie 2022, 53, 4. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Rortais, A.; Arnold, G.; Alburaki, M.; Legout, H.; Garnery, L. Review of the DraI COI-COII test for the conservation of the black honeybee (Apis mellifera mellifera). Conserv. Genet. Resour. 2011, 3, 383–391. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. PopArt: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Wragg, D.; Marti-Marimon, M.; Basso, B.; Bidanel, J.P.; Labarthe, E.; Bouchez, O.; Le Conte, Y.; Vignal, A. Whole-genome resequencing of honey bee drones to detect genomic selection in a population managed for royal jelly. Sci. Rep. 2016, 6, 27168. [Google Scholar] [CrossRef]
- Techer, M.A.; Clémencet, J.; Simiand, C.; Preeaduth, S.; Azali, H.A.; Reynaud, B.; Hélène, D. Large-scale mitochondrial DNA analysis of native honey bee Apis mellifera populations reveals a new African subgroup private to the South West Indian Ocean islands. BMC Genet. 2017, 18, 53. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Garnery, L.; Solignac, M. Putative origin and function of the intergenic region between COI and COII of Apis mellifera L. mitochondrial DNA. Genetics 1991, 128, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Alburaki, M.; Madella, S.; Lopez, J.; Bouga, M.; Chen, Y.; vanEngelsdorp, D. Honey bee populations of the USA display restrictions in their mtDNA haplotype diversity. Front. Genet. 2023, 13, 1092121. [Google Scholar] [CrossRef] [PubMed]
- Wragg, D.; Techer, M.A.; Canale-Tabet, K.; Basso, B.; Bidanel, J.P.; Labarthe, E.; Bouchez, O.; Le Conte, Y.; Clémencet, J.; Delatte, H.; et al. Autosomal and Mitochondrial Adaptation Following Admixture: A Case Study on the Honey bees of Reunion Island. Genome Biol. Evol. 2018, 10, 220–238. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.S.; Chahbar, N.; Abed, F.; Oussalah, N.; Kandemir, I.; Ainouche, A.; Doumandji, S. Mitochondrial DNA variation of Algerian honey bees Apis mellifera intermissa and Apis mellifera sahariensis based on COI-COII RFLP and sequencing. Res. Sq. 2023; preprint. [Google Scholar] [CrossRef]
- Barour, C.; Tahar, A.; Baylac, M. Forewing Shape Variation in Algerian Honey Bee Populations of Apis mellifera intermissa (Buttel-Reepen, 1906) (Hymenoptera: Apidae): A Landmark-Based Geometric Morphometrics Analysis. Afr. Entomol. 2011, 19, 11–22. [Google Scholar] [CrossRef]
- Bendjedid, H.; Achou, M. Etude de la Diversité Morphométrique de Deux Populations d’Abeilles Domestiques (Apis mellifera intermissa et Apis mellifera sahariensis) du Sud Algérien. Synthèse Rev. Sci. Technol. 2014, 28, 84–95. [Google Scholar] [CrossRef]
- Bouzeraa, H.; Sellami, H.; Gdoura, R.; Achou, M.; Soltani, N. Genetic diversity of the Honey bee Apis mellifera Linnaeus, 1758 (Hymenoptera Apidae) from Jijel (Northeast Algeria). Biodivers. J. 2020, 11, 7–14. [Google Scholar] [CrossRef]
- Achou, M.; Loucif-Ayad, W.; Legout, H.; Hmidan, H.; Alburaki, M.; Garnery, L. An Insightful Molecular Analysis Reveals Foreign Honey bees Among Algerian Honey bee Populations (Apis mellifera L.). J. Data Min. Genom. Proteom. 2015, 6, 166. [Google Scholar] [CrossRef]
- Garnery, L.; Franck, P.; Baudry, E.; Vautrin, D.; Cornuet, J.-M.; Solignac, M. Genetic diversity of the west European honey bee (Apis mellifera mellifera and A. m. iberica). I. Mitochondrial DNA. Genet. Sel. Evol. 1998, 30, S31–S47. [Google Scholar] [CrossRef]
- Salvatore, G.; Chibani Bahi Amar, A.; Canale-Tabet, K.; Fridi, R.; Tabet Aoul, N.; Saci, S.; Labarthe, E.; Palombo, V.; D’Andrea, M.; Vignal, A.; et al. Natural clines and human management impact the genetic structure of Algerian honey bee populations. Genet. Sel. Evol. 2023, 55, 94. [Google Scholar] [CrossRef]
- Rasolofoarivao, H.; Clémencet, J.; Techer, M.A.; Ravaomanarivo, L.H.R.; Reynaud, B.; Delatte, H. Genetic diversity of the endemic honey bee: Apis mellifera unicolor (Hymenoptera: Apidae) in Madagascar. Apidologie 2015, 46, 735–747. [Google Scholar] [CrossRef]
- Hepburn, H.R.; Radloff, S.E. Morphometric and Pheromonal Analyses of Apis mellifera L. along a Transect from the Sahara to the Pyrenees. Apidologie 1996, 27, 35–45. [Google Scholar] [CrossRef]
- Smith, F.G. The Races of Honey bees in Africa. Bee World 1961, 42, 255–260. [Google Scholar] [CrossRef]
- Bertrand, B.; Alburaki, M.; Legout, H.; Moulin, S.; Mougel, F.; Garnery, L. MtDNA COI-COII marker and drone congregation area: An efficient method to establish and monitor honey bee (Apis mellifera L.) conservation centres. Mol. Ecol. Resour. 2015, 15, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Wragg, D.; Eynard, S.E.; Basso, B.; Canale-Tabet, K.; Labarthe, E.; Bouchez, O.; Bienefeld, K.; Bieńkowska, M.; Costa, C.; Gregorc, A.; et al. Complex population structure and haplotype patterns in the Western European honey bee from sequencing a large panel of haploid drones. Mol. Ecol. Resour. 2022, 22, 3068–3086. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Areas | Intergenic Variant | Percentage of Occurrence |
|---|---|---|
| Algeria | P0Q | 35.3% |
| P0QQ | 61.7% | |
| P0QQQ | 3% | |
| SWIO and Madagascar | P0Q | 92.9% |
| Q | 7.1% | |
| Europe | P0QQ | 3.4% |
| PQ | 13.8% | |
| PQQ | 27.6% | |
| Q | 55.2% |
| L 1 | GS 2 | Subspecies | OS 3 | Cities | Haplotypes | GB Accession 4 |
|---|---|---|---|---|---|---|
| A | P0Q | intermissa sahariensis | Algeria | Biskra; AinSefra; Bejaia; Chlef; Bayadh; Tissemssilet | A1x | PP667401 |
| A | P0Q | intermissa | Algeria | Bejaia | A8k | PP667402 |
| A | P0QQ | intermissa | Algeria | Mascara; Mostaganem; Relizane | A68 | PP667403 |
| A | P0QQ | intermissa | Algeria | Mostaganem | A69 | PP667404 |
| A | P0QQ | intermissa | Algeria | Bejaia | A70 | PP667405 |
| A | P0QQ | intermissa | Algeria | Sidibelabes | A71a | PP667406 |
| A | P0QQ | intermissa | Algeria | Sidibelabes; Relizane | A72 | PP667407 |
| A | P0QQ | intermissa | Algeria | Chlef; Sidibelabes; Mostaganem | A73 | PP667408 |
| A | P0QQ | intermissa | Algeria | Mascara | A70a | PP667409 |
| A | P0QQ | intermissa | Algeria | Mascara | A74a | PP667410 |
| A | P0QQ | intermissa | Algeria | Mostaganem | A73a | PP667411 |
| A | P0QQ | intermissa sahariensis | Algeria | Bayadh; Mostaganem; Tlemcen Oran; Relizane; Ghardaia; Djelfa | A74 | PP667412 |
| A | P0QQ | sahariensis | Algeria | Djelfa | A74b | PP667413 |
| A | P0QQ | sahariensis | Algeria | AinSefra | A75 | PP667414 |
| A | P0QQ | intermissa | Algeria | Sidibelabes | A76 | PP667415 |
| A | P0QQQ | intermissa | Algeria | Mascara | A77′ | PP667416 |
| M | PQQ | mellifera | France | NA 5 | M4r | PP667417 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chibani Bahi Amar, A.; Tabet Aoul, N.; Fridi, R.; Vignal, A.; Canale-Tabet, K. New COI-COII mtDNA Region Haplotypes in the Endemic Honey Bees Apis mellifera intermissa and Apis mellifera sahariensis (Hymenoptera: Apidae) in Algeria. Insects 2024, 15, 549. https://doi.org/10.3390/insects15070549
Chibani Bahi Amar A, Tabet Aoul N, Fridi R, Vignal A, Canale-Tabet K. New COI-COII mtDNA Region Haplotypes in the Endemic Honey Bees Apis mellifera intermissa and Apis mellifera sahariensis (Hymenoptera: Apidae) in Algeria. Insects. 2024; 15(7):549. https://doi.org/10.3390/insects15070549
Chicago/Turabian StyleChibani Bahi Amar, Amira, Nacera Tabet Aoul, Riad Fridi, Alain Vignal, and Kamila Canale-Tabet. 2024. "New COI-COII mtDNA Region Haplotypes in the Endemic Honey Bees Apis mellifera intermissa and Apis mellifera sahariensis (Hymenoptera: Apidae) in Algeria" Insects 15, no. 7: 549. https://doi.org/10.3390/insects15070549