
Safety of the Entomopathogenic Fungus Beauveria bassiana for Wild and Laboratory-Reared Chrysoperla lucasina Strains



Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Fungal Preparations
2.2. Insect Rearing
2.3. Insect Bioassays
2.3.1. Survival Bioassays
2.3.2. Bioassays on Life-Table Parameters
2.4. Data Analysis and Statistics
3. Results
3.1. Survival Bioassays
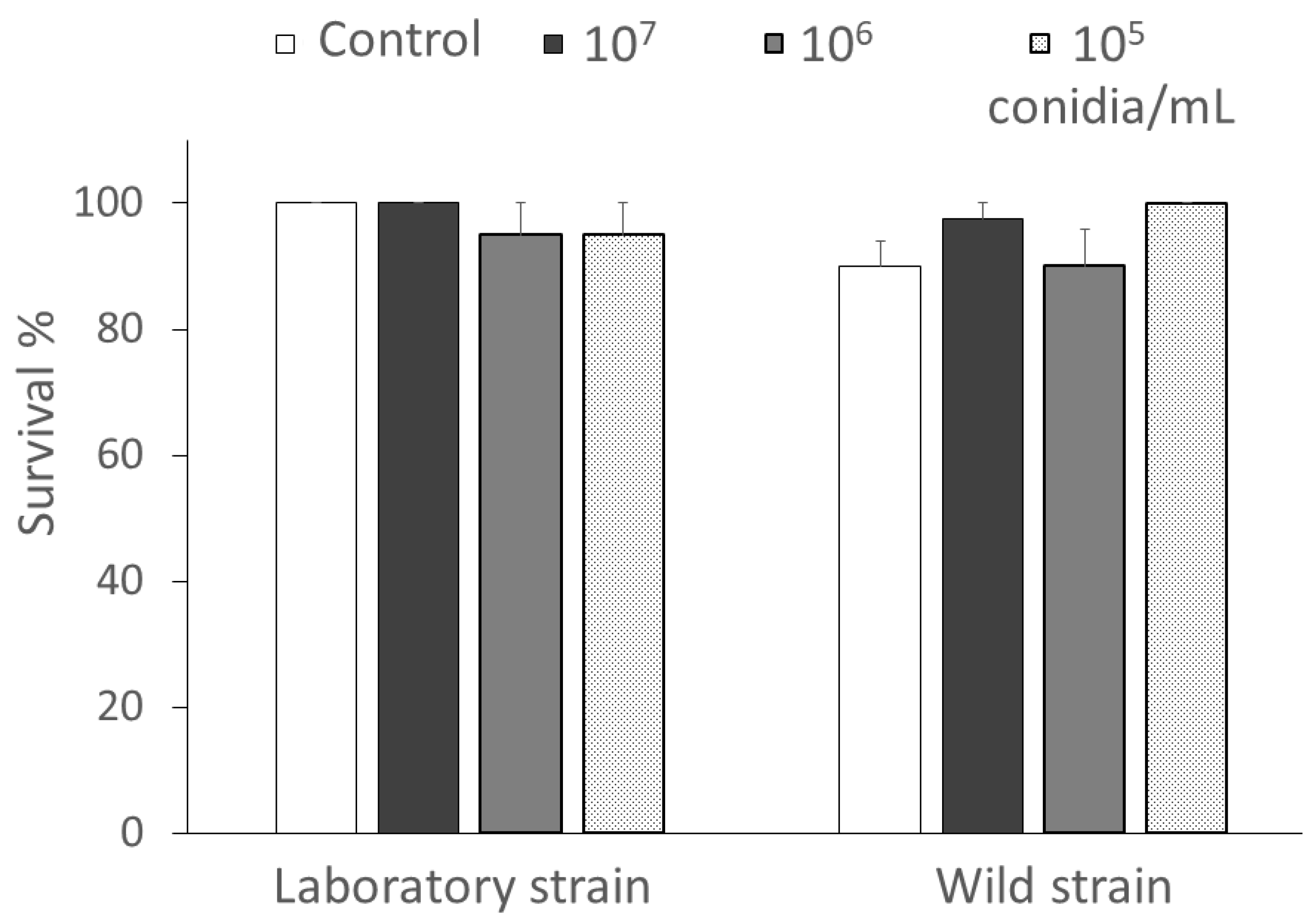
3.1.1. Bioassays with Chrysoperla lucasina
3.1.2. Bioassays with Tenebrio molitor
3.2. Bioassays on Life-Table Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baker, B.P.; Green, T.A.; Loker, A.J. Biological control and integrated pest management in organic and conventional systems. Biol. Control 2020, 140, 104095. [Google Scholar] [CrossRef]
- Pappas, M.L.; Broufas, G.D.; Koveos, D.S. Chrysopid predators and their role in biological control. J. Entomol. 2011, 8, 301–326. [Google Scholar] [CrossRef]
- Gentz, M.C.; Murdoch, G.; King, G.F. Tandem use of selective insecticides and natural enemies for effective, reduced-risk pest management. Biol. Control 2010, 52, 208–215. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Urquiza, A.; Riveiro-Miranda, L.; Santiago-Álvarez, C.; Quesada-Moraga, E. Insect-toxic secreted proteins and virulence of the entomopathogenic fungus Beauveria bassiana. J. Invertebr. Pathol. 2010, 105, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Moraga, E.; Garrido-Jurado, I.; González-Mas, N.; Yousef-Yousef, M. Ecosystem services of entomopathogenic ascomycetes. J. Invertebr. Pathol. 2023, 201, 108015. [Google Scholar] [CrossRef]
- Koller, J.; Sutter, L.; Gonthier, J.; Collatz, J.; Norgrove, L. Entomopathogens and parasitoids allied in biocontrol: A systematic review. Pathogens 2023, 12, 957. [Google Scholar] [CrossRef]
- Thungrabeab, M.; Tongma, S. Effect of entomopathogenic fungi, Beauveria bassiana (Balsam) and Metarhizium anisopliae (Metsch) on non-target insects. Curr. Appl. Sci. Technol. 2007, 7, 8–12. [Google Scholar]
- Aguila, L.C.R.; Akutse, K.S.; Ashraf, H.J.; Bamisile, B.S.; Lin, J.; Dai, J.; Wang, H.; Wang, L. The survival and parasitism rate of Tamarixia radiata (Hymenoptera: Eulophidae) on its host exposed to Beauveria bassiana (Ascomycota: Hypocreales). Agronomy 2021, 11, 1496. [Google Scholar] [CrossRef]
- Al-mazra’awi, M.S. Impact of the entomopathogenic fungus Beauveria bassiana on the honeybees, Apis mellifera (Hymenoptera: Apidae). J. Agric. Sci. 2007, 3, 7–11. [Google Scholar]
- Cappa, F.; Petrocelli, I.; Dani, F.R.; Dapporto, L.; Giovannini, M.; Silva-Castellari, J.; Turillazzi, S.; Cervo, R. Natural biocide disrupts nestmate recognition in honeybees. Sci. Rep. 2019, 9, 3171. [Google Scholar] [CrossRef] [PubMed]
- Atzeni, R.; Moro, G.; Marche, M.G.; Uva, P.; Ruiu, L. Genome sequence of Beauveria bassiana strain ATCC 74040, a widely employed insect pathogen. Microbiol. Res. Announc. 2020, 9, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, E. Prove di allevamento in ambiente condizionato di Chrysopa carnea Steph. (Neuroptera, Chrysopidae). Boll. Istit. Entomol. 1975, 32, 291–304. [Google Scholar]
- Loru, L.; Fois, X.; Sassu, A.; Pantaleoni, R.A. An individual marking technique for green lacewings (Neuroptera: Chrysopidae). Fla. Entomol. 2013, 96, 628–630. [Google Scholar] [CrossRef]
- Ruiu, L.; Lazzeri, A.M.; Nuvoli, M.T.; Floris, I.; Satta, A. Safety evaluation of the entomopathogenic bacterium Brevibacillus laterosporus for the green lacewing Chrysoperla agilis (Neuroptera: Chrysopidae). J. Invertebr. Pathol. 2020, 169, 107281. [Google Scholar] [CrossRef] [PubMed]
- Ehrenberg, S.; Lewkowski, O.; Erler, S. Dyeing but not dying: Colourful dyes as a non-lethal method of food labelling for in vitro-reared honey bee (Apis mellifera) larvae. J. Insect Physiol. 2019, 113, 1–8. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Ahrens, W.H.; Cox, D.J.; Budhwar, G. Use of the arcsine and square root transformations for subjectively determined percentage data. Weed Sci. 1990, 38, 452–458. [Google Scholar] [CrossRef]
- Keselman, H.J.; Rogan, J.C. The Tukey multiple comparison test: 1953–1976. Psychol. Bull. 1977, 84, 1050. [Google Scholar] [CrossRef]
- Collier, T.; Van Steenwyk, R. A critical evaluation of augmentative biological control. Biol. Control 2004, 31, 245–256. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Nicoli, G.; Heinz, K.M.; Parella, M.P. Quality control of mass-produced beneficial insects. In Biocontrol in Protected Culture; Heinz, K.M., Van Driesche, R., Parrella, M.P., Eds.; Ball Publishing: Batavia, IL, USA, 2004; pp. 503–527. [Google Scholar]
- Gilkeson, L.A. Ecology of rearing: Quality, regulation, and mass rearing. In Ecological Interactions and Biological Control, 1st ed.; Andow, D.A., Ragsdale, D.W., Nyvall, R.F., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 139–148. [Google Scholar]
- Grenier, S.; Clercq, P.D. Comparison of artificially vs. naturally reared natural enemies and their potential for use in biological control. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; Van Lenteren, J.C., Ed.; CABI Publishing: Wallingford, UK, 2003; pp. 115–131. [Google Scholar]
- Quesada-Moraga, E.; Yousef-Naef, M.; Garrido-Jurado, I. Advances in the use of entomopathogenic fungi as biopesticides in suppressing crop pests. In Biopesticides for Sustainable Agriculture, 1st ed.; Birch, N., Glare, T., Eds.; Burleigh Dodds Science Publishing: London, UK, 2020; pp. 63–98. [Google Scholar]
- Mannino, M.C.; Huarte-Bonnet, C.; Davyt-Colo, B.; Pedrini, N. Is the insect cuticle the only entry gate for fungal infection? Insights into alternative modes of action of entomopathogenic fungi. J. Fungi 2019, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Mingotti Dias, P.; de Souza Loureiro, E.; Amorim Pessoa, L.G.; Mendes de Oliveira Neto, F.; de Souza Tosta, R.A.; Teodoro, P.E. Interactions between fungal-infected Helicoverpa armigera and the predator Chrysoperla externa. Insects 2019, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Leyva, O.E.; Villalon, E.M.; Avila, R.A.; Bulet, D.B. Susceptibilidad de Chrysopa exterior Navas a Beauveria bassiana (Blasamo) Vuillemin cepa LBB-1 en condiciones de laboratorio. Fitosanidad 2011, 15, 51–57. [Google Scholar]
- Hamze, R.; Nuvoli, M.T.; Pirino, C.; Ruiu, L. Compatibility of the bacterial entomopathogen Pseudomonas protegens with the natural predator Chrysoperla carnea (Neuroptera: Chrysopidae). J. Invertebr. Pathol. 2022, 194, 107828. [Google Scholar] [CrossRef] [PubMed]
- Broza, M.; Pereira, R.M.; Stimac, J.L. The nonsusceptibility of soil Collembola to insect pathogens and their potential as scavengers of microbial pesticides. Pedobiologia 2001, 45, 523–534. [Google Scholar] [CrossRef]
- Dromph, K.M.; Vestergaard, S. Pathogenicity and attractiveness of entomopathogenic hyphomycete fungi to collembolans. Appl. Soil Ecol. 2002, 21, 197–210. [Google Scholar] [CrossRef]
- Amorim, L.G.; Sousa, R.; Moino, A.; Souza, B. Compatibility between Beauveria bassiana and the predator Chrysoperla externa in laboratory. Pesqui. Agropecu. Bras. 2005, 40, 6. [Google Scholar]
- Portilla, M.; Snodgrass, G.; Luttrell, R. Lethal and sub-lethal effects of Beauveria bassiana (Cordycipitaceae) strain NI8 on Chrysoperla rufilabris (Neuroptera: Chrysopidae). Fla. Entomol. 2017, 100, 627–633. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, Y.; Xu, W.; Sui, L.; Du, Q.; Wang, Y.; Zhao, Y.; Li, Q. Influence of genetic diversity of seventeen Beauveria bassiana isolates from different hosts on virulence by comparative genomics. BMC Genom. 2020, 21, 451. [Google Scholar] [CrossRef]
- Wakil, W.; Boukouvala, M.C.; Kavallieratos, N.G.; Naeem, A.; Ghazanfar, M.U.; Alhewairini, S.S. Impact of Three Entomopathogenic Fungal Isolates on the Growth of Tomato Plants—Ectoapplication to Explore Their Effect on Tetranychus urticae. Agronomy 2024, 14, 665. [Google Scholar] [CrossRef]
- Usman, M.; Wakil, W.; Piñero, J.C.; Wu, S.; Toews, M.D.; Shapiro-Ilan, D.I. Evaluation of locally isolated entomopathogenic fungi against multiple life stages of Bactrocera zonata and Bactrocera dorsalis (Diptera: Tephritidae): Laboratory and field study. Microorganisms 2021, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Kim, J.J. Target-oriented dissemination of Beauveria bassiana conidia by the predators, Harmonia axyridis (Coleoptera: Coccinellidae) and Chrysoperla carnea (Neuroptera: Chrysopidae) for biocontrol of Myzus persicae. Biocontrol Sci. Technol. 2012, 22, 393–406. [Google Scholar] [CrossRef]
- Donegan, K.; Lighthart, B. Effect of several stress factors on the susceptibility of the predatory insect, Chrysoperla carnea (Neuroptera: Chrysopidae), to the fungal pathogen Beauveria bassiana. J. Invertebr. Pathol. 2003, 54, 79–84. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beauveria bassiana Concentration (Conidia/mL) | Development Time (Days) | Adult Emergence c % | Sex Ratio (Male %) | |
|---|---|---|---|---|
| Larvae a | Pupae b | |||
| Laboratory strain | ||||
| 107 | 16.3 ± 1.26 a d | 11.5 ± 0.93 a | 92.5 ± 2.50 ab | 73.1 ± 5.33 b |
| 106 | 16.6 ± 1.62 a | 11.4 ± 1.19 a | 95.0 ± 2.89 a | 55.0 ± 5.49 a |
| 105 | 16.8 ± 1.42 a | 11.1 ± 1.01 a | 92.5 ± 2.50 ab | 59.4 ± 5.24 a |
| Control | 15.8 ± 1.37 a | 11.5 ± 1.03 a | 97.5 ± 2.50 a | 49.4 ± 12.48 a |
| Wild strain | ||||
| 107 | 17.5 ± 1.29 a | 11.71 ± 0.99 a | 80.0 ± 7.07 b | 51.6 ± 10.70 a |
| 106 | 19.0 ± 1.98 a | 11.83 ± 1.02 a | 82.5 ± 8.54 b | 37.5 ± 11.09 a |
| 105 | 17.8 ± 1.90 a | 12.0 ± 0.89 a | 85.0 ± 6.45 b | 31.5 ± 12.78 a |
| Control | 17.1 ±1.20 a | 11.7 ± 0.91 a | 85.0 ± 6.45 b | 54.6 ± 6.61 a |
| Beauveria bassiana Concentrations (Conidia/mL) | N a | Eggs/Female | Egg Hatching b % |
|---|---|---|---|
| Laboratory strain | |||
| 107 | 11 | 288.4 ± 46.03 c | 55.0 ± 8.66 |
| 106 | 17 | 355.0 ± 54.94 | 60.0 ± 4.08 |
| 105 | 16 | 234.8 ± 63.38 | 62.0 ± 6.29 |
| Control | 18 | 294.5 ± 64.42 | 70.0 ± 4.08 |
| Wild strain | |||
| 107 | 13 | 211.9 ± 54.27 | 72.5 ± 11.09 |
| 106 | 16 | 182.7 ± 31.75 | 77.5 ± 7.50 |
| 105 | 16 | 237.6 ± 14.44 | 67.5 ± 8.53 |
| Control | 14 | 254.1 ± 57.32 | 55.0 ± 6.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morda, W.; Nuvoli, M.T.; Ruiu, L. Safety of the Entomopathogenic Fungus Beauveria bassiana for Wild and Laboratory-Reared Chrysoperla lucasina Strains. Insects 2024, 15, 576. https://doi.org/10.3390/insects15080576
Morda W, Nuvoli MT, Ruiu L. Safety of the Entomopathogenic Fungus Beauveria bassiana for Wild and Laboratory-Reared Chrysoperla lucasina Strains. Insects. 2024; 15(8):576. https://doi.org/10.3390/insects15080576
Chicago/Turabian StyleMorda, Walaa, Maria Tiziana Nuvoli, and Luca Ruiu. 2024. "Safety of the Entomopathogenic Fungus Beauveria bassiana for Wild and Laboratory-Reared Chrysoperla lucasina Strains" Insects 15, no. 8: 576. https://doi.org/10.3390/insects15080576
APA StyleMorda, W., Nuvoli, M. T., & Ruiu, L. (2024). Safety of the Entomopathogenic Fungus Beauveria bassiana for Wild and Laboratory-Reared Chrysoperla lucasina Strains. Insects, 15(8), 576. https://doi.org/10.3390/insects15080576