
Linear Morphometry of Male Genitalia Distinguishes the Ant Genera Monomorium and Syllophopsis (Hymenoptera: Formicidae) in Madagascar
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source
2.2. Slide Preparation and Measurements
2.3. Morphometric Data
2.4. Data Analyses
3. Results
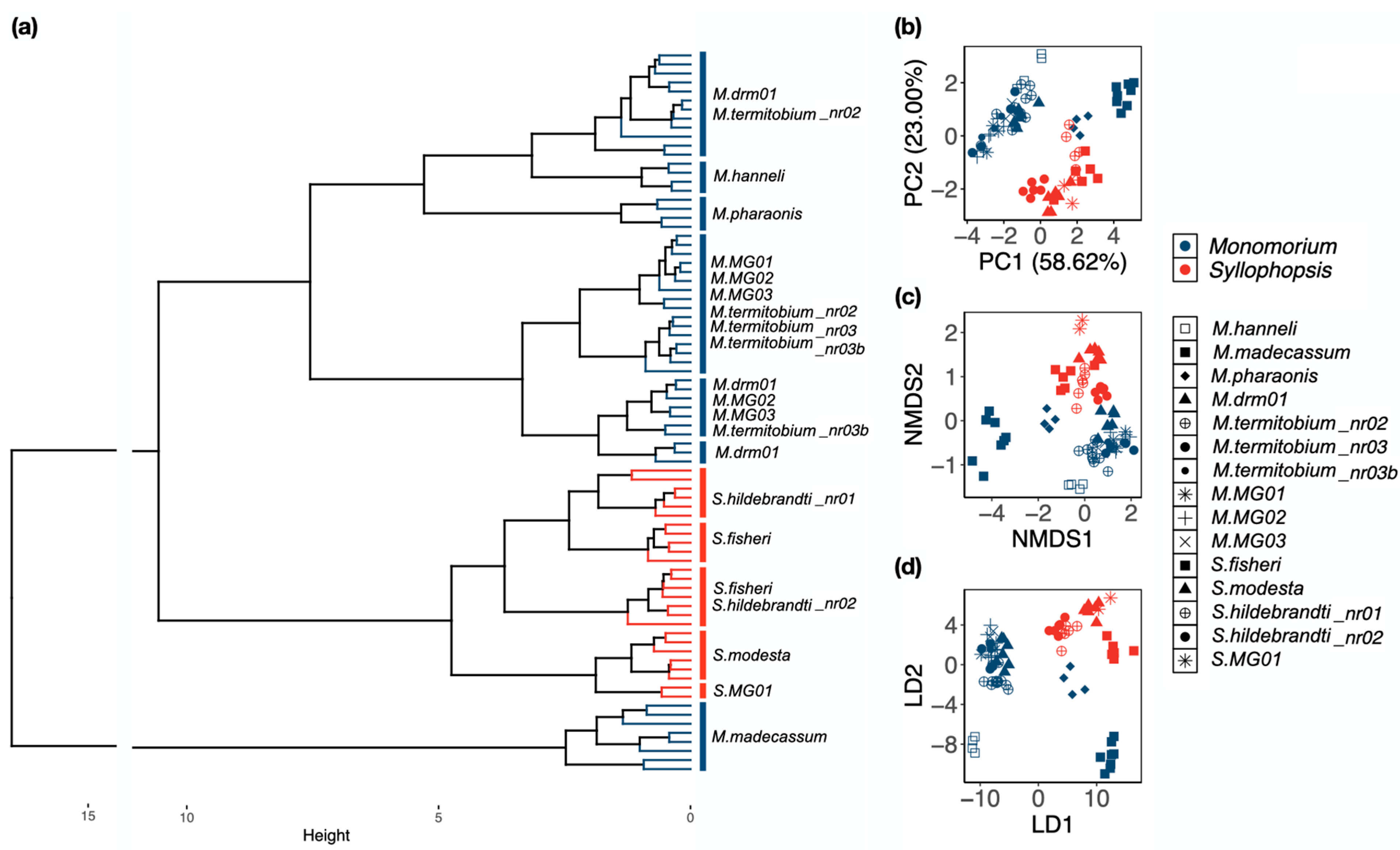
3.1. Multivariate Analysis of Morphometric Data
3.1.1. Raw Data
3.1.2. Ratio Data
3.1.3. RAV Data (Removal Allometric Variance)
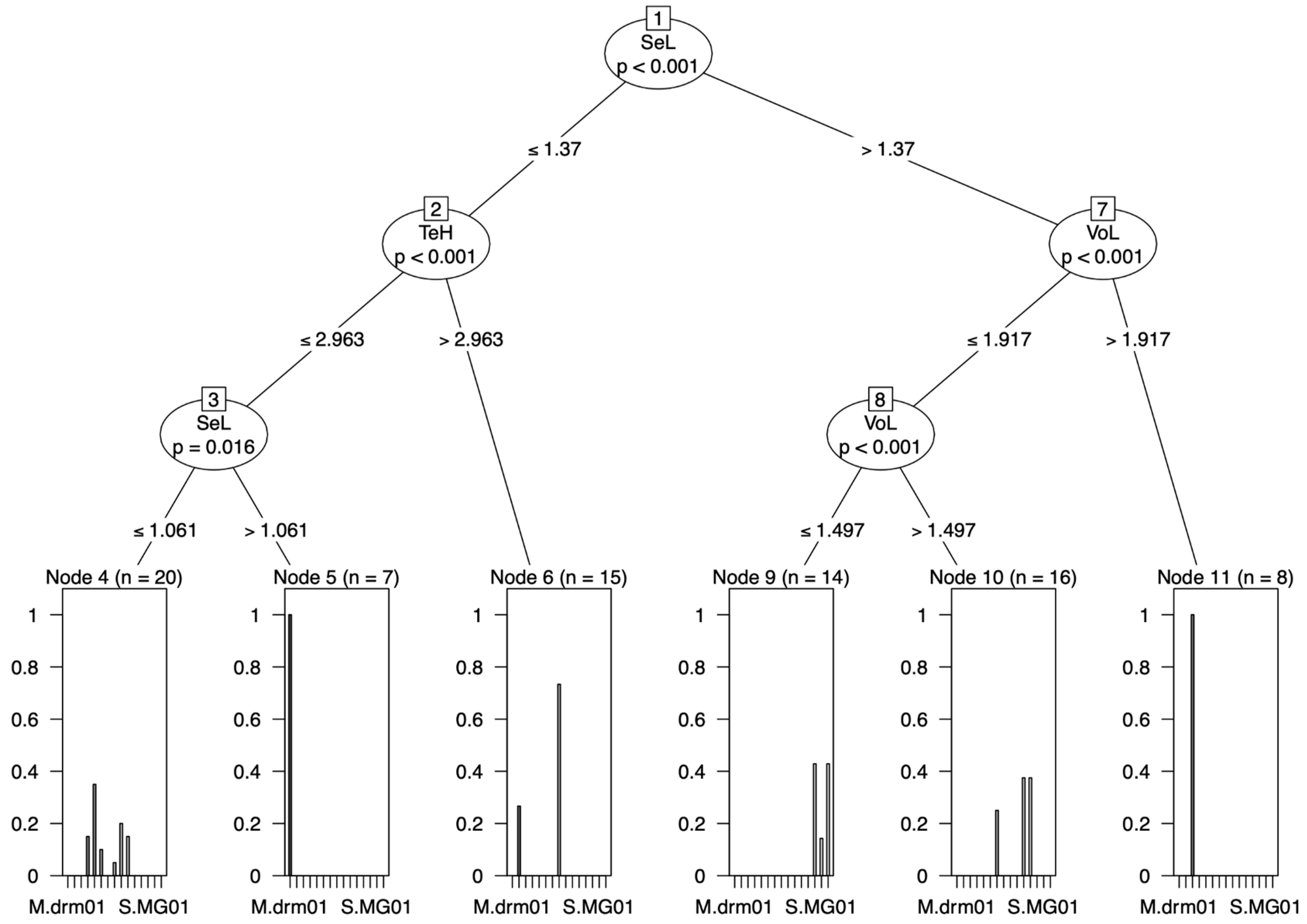
3.2. Conditional Inference Trees (CITs)
3.2.1. Raw Data
3.2.2. Ratio Data
3.2.3. RAV Data
4. Discussion
4.1. Multivariate Analysis of Morphometric Data
4.2. Conditional Inference Trees (CITs)
4.3. Caveats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eberhard, W.G. Evolution of genitalia: Theories, evidence, and new directions. Genetica 2010, 138, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.M.; Porter, A.H. The lock-and-key hypothesis: Evolutionary and biosystematic interpretation of insect genitalia. Annu. Rev. Entomol. 1989, 34, 231–245. [Google Scholar] [CrossRef]
- Tuxen, S.L. (Ed.) Taxonomist’s Glossary of Genitalia in Insects; Munksgaard: Copenhagen, Denmark, 1970; ISBN 978-87-16-00035-4. [Google Scholar]
- Hosken, D.J.; Stockley, P. Sexual selection and genital evolution. Trends Ecol. Evol. 2004, 19, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Simmons, L.W.; Fitzpatrick, J.L. Female genitalia can evolve more rapidly and divergently than male genitalia. Nat. Commun. 2019, 10, 1312. [Google Scholar] [CrossRef] [PubMed]
- House, C.; Simmons, L. Genital morphology and fertilization success in the dung beetle Onthophagus taurus: An example of sexually selected male genitalia. Proc. Biol. Sci. R. Soc. 2003, 270, 447–455. [Google Scholar] [CrossRef]
- Song, H.; Bucheli, S.R. Comparison of phylogenetic signal between male genitalia and non-genital characters in insect systematics. Cladistics 2010, 26, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, F.; Matsumura, Y.; Pohl, H.; Bai, M.; Hörnschemeyer, T.; Beutel, R.G. Insect morphology in the age of phylogenomics: Innovative techniques and its future role in systematics. Entomol. Sci. 2014, 17, 1–24. [Google Scholar] [CrossRef]
- Cantone, S.; Di Giulio, A. A new Neotropical ant species of genus Linepithema Mayr (Hymenoptera, Formicidae, Dolichoderinae) with partial revision of the L. fuscum group based on males. ZooKeys 2023, 1160, 125–144. [Google Scholar] [CrossRef]
- Hita Garcia, F.; Fischer, G.; Liu, C.; Audisio, T.L.; Alpert, G.D.; Fisher, B.L.; Economo, E.P. X-ray Microtomography for ant taxonomy: An exploration and case study with two new Terataner (Hymenoptera, Formicidae, Myrmicinae) species from Madagascar. PLoS ONE 2017, 12, e0172641. [Google Scholar] [CrossRef]
- Sharaf, M.R.; Gotzek, D.; Guénard, B.; Fisher, B.L.; Aldawood, A.S.; Al Dhafer, H.M.; Mohamed, A.A. Molecular phylogenetic analysis and morphological reassessments of thief ants identify a new potential case of biological invasions. Sci. Rep. 2020, 10, 12040. [Google Scholar] [CrossRef]
- Tóth, J. Morphometric study on the genitalia of sibling species Melitaea phoebe and M. telona (Lepidoptera: Nymphalidae). Acta Zool. Acad. Sci. Hung. 2010, 56, 273–282. [Google Scholar]
- Tschinkel, W.R. The morphometry of Solenopsis fire ants. PLoS ONE 2013, 8, e79559. [Google Scholar] [CrossRef]
- Dapporto, L. Geometric morphometrics reveal male genitalia differences in the Lasiommata megera/paramegaera complex (Lepidoptera, Nymphalidae) and the lack of a predicted hybridization area in the Tuscan Archipelago. J. Zool. Syst. Evol. Res. 2008, 46, 224–230. [Google Scholar] [CrossRef]
- Tatsuta, H.; Takahashi, K.H.; Sakamaki, Y. Geometric morphometrics in Entomology: Basics and applications. Entomol. Sci. 2018, 21, 164–184. [Google Scholar] [CrossRef]
- Tozetto, L.; Lattke, J.E. Revealing male genital morphology in the giant ant genus Dinoponera with geometric morphometrics. Arthropod Struct. Dev. 2020, 57, 100943. [Google Scholar] [CrossRef] [PubMed]
- Seifert, B. Removal of Allometric Variance improves species separation in multi-character discriminant functions when species are strongly allometric and exposes diagnostic characters. Myrmecol. News 2008, 11, 91–105. [Google Scholar]
- Baur, H.; Leuenberger, C. Analysis of ratios in multivariate morphometry. Syst. Biol. 2011, 60, 813–825. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 978-0-444-53868-0. [Google Scholar]
- Jauset, A.; Edo-Tena, E.; Parés-Casanova, P.; Castañé, C.; Agustí, N.; Alomar, O. Elliptic Fourier Analysis in the study of the male genitalia to discriminate three Macrolophus species (Hemiptera: Miridae). Insects 2017, 8, 120. [Google Scholar] [CrossRef]
- Derkarabetian, S.; Castillo, S.; Koo, P.K.; Ovchinnikov, S.; Hedin, M. A demonstration of unsupervised machine learning in species delimitation. Mol. Phylogenetics Evol. 2019, 139, 106562. [Google Scholar] [CrossRef]
- Yeates, D.K.; Seago, A.; Nelson, L.; Cameron, S.L.; Joseph, L.; Trueman, J.W.H. Integrative taxonomy, or iterative taxonomy? Syst. Entomol. 2011, 36, 209–217. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative taxonomy: A multisource approach to exploring biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.C.; Arthofer, W.; Seifert, B.; Muster, C.; Steiner, F.M.; Schlick-Steiner, B.C. Light at the end of the tunnel: Integrative taxonomy delimits cryptic species in the Tetramorium caespitum complex (Hymenoptera: Formicidae). Myrmecol. News 2017, 25, 95. [Google Scholar]
- Tarasov, S.I.; Solodovnikov, A.Y. Phylogenetic analyses reveal reliable morphological markers to classify mega-diversity in Onthophagini dung beetles (Coleoptera: Scarabaeidae: Scarabaeinae). Cladistics 2011, 27, 490–528. [Google Scholar] [CrossRef] [PubMed]
- Noboa, M.; Viera, W.; Díaz, A.; Vásquez, W.; Ron, L. Genitalic differentiations in Neoleucinodes elegantalis (Gueneé) (Lepidoptera: Crambidae) associated with Solanaceae crops in Ecuador. Insects 2017, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- Mutanen, M.; Kaitala, A. Genital variation in a dimorphic moth Selenia tetralunaria (Lepidoptera, Geometridae): Genital variation in a dimorphic moth. Biol. J. Linn. Soc. 2006, 87, 297–307. [Google Scholar] [CrossRef]
- Prieto, C.G.; Munguira, M.L.; Romo, H. Morphometric analysis of genitalia and wing pattern elements in the genus Cupido (Lepidoptera, Lycaenidae): Are Cupido minimus and C. carswelli different species? Dtsch. Entomol. Z. 2009, 56, 137–147. [Google Scholar] [CrossRef]
- Simonsen, T.J. Boloria phylogeny (Lepidoptera: Nymphalidae): Tentatively reconstructed on the basis of male and female genitalic morphology: Bolaria phylogeny. Syst. Entomol. 2005, 30, 653–665. [Google Scholar] [CrossRef]
- Belintani, T.; de Paiva, V.F.; de Oliveira, J.; da Rosa, J.A. New in morphometry: Geometric morphometry of the external female genitalia of Triatominae (Hemiptera: Reduviidae). Acta Trop. 2022, 229, 106383. [Google Scholar] [CrossRef] [PubMed]
- Knyshov, A.; Hoey-Chamberlain, R.; Weirauch, C. Comparative morphology of male genitalic structures in the minute litter bugs Dipsocoromorpha (Insecta: Hemiptera: Heteroptera). J. Morphol. 2018, 279, 1480–1517. [Google Scholar] [CrossRef]
- Rajonhson, D.M.; Raksakoon, C.; Payakkapol, A.; Dujardin, S.; Dujardin, J.-P.; Potiwat, R. Comparison of two different morphological methods to study the pronotum of Cimicidae: Bed bugs, bat bugs, and swallow bugs. Insects 2022, 13, 1155. [Google Scholar] [CrossRef]
- Wieczorek, K.; Płachno, B.J.; Świątek, P. Comparative morphology of the male genitalia of Aphididae (Insecta, Hemiptera): Part 1. Zoomorphology 2011, 130, 289–303. [Google Scholar] [CrossRef]
- Wieczorek, K.; Płachno, B.J.; Świątek, P. A comparative morphology of the male genitalia of Aphididae (Insecta, Hemiptera): Part 2. Zoomorphology 2012, 131, 303–324. [Google Scholar] [CrossRef]
- Singh, N.S.; Phillips-S, D.; Lal, D. Role of morphometric description of female genitalia to discriminate Phlebotomine sand flies species (Diptera; Psychodidae; Phlebotominae) from Northern part of India. J. Entomol. 2012, 9, 389–395. [Google Scholar] [CrossRef]
- Laciny, A.; Zettel, H.; Kopchinskiy, A.; Pretzer, C.; Pal, A.; Abu Salim, K.; Javad Rahimi, M.; Hoenigsberger, M.; Lim, L.; Jaitrong, W.; et al. Colobopsis explodens sp. n., model species for studies on “exploding ants” (Hymenoptera, Formicidae), with biological notes and first illustrations of males of the Colobopsis cylindrica group. ZooKeys 2018, 751, 1–40. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Moder, K.; Seifert, B.; Sanetra, M.; Dyreson, E.; Stauffer, C.; Christian, E. A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol. Phylogenetics Evol. 2006, 40, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, M.; Fisher, B.L. A revision of male ants of the Malagasy region (Hymenoptera: Formicidae): Key to subfamilies and treatment of the genera of Ponerinae. Zootaxa 2007, 1654, 21–40. [Google Scholar] [CrossRef]
- Yoshimura, M.; Fisher, B.L. A revision of male ants of the Malagasy region (Hymenoptera: Formicidae): Key to genera of the subfamily Proceratiinae. Zootaxa 2009, 2216, 1–21. [Google Scholar] [CrossRef]
- Yoshimura, M.; Fisher, B.L. A revision of male ants of the Malagasy region (Hymenoptera: Formicidae): Key to genera of the subfamily Dolichoderinae. Zootaxa 2011, 2794, 1–34. [Google Scholar] [CrossRef]
- Yoshimura, M.; Fisher, B.L. A revision of the Malagasy endemic genus Adetomyrma (Hymenoptera: Formicidae: Amblyoponinae). Zootaxa 2012, 3341, 1–31. [Google Scholar] [CrossRef]
- Wilson, C.M.; Smith-Herron, A.; Cook, J.L. Morphology of the male genitalia of Brachymyrmex and their implications in the Formicinae phylogeny. J. Hymenopt. Res. 2016, 50, 81–95. [Google Scholar] [CrossRef]
- Boudinot, B.E. The male genitalia of ants: Musculature, homology, and functional morphology (Hymenoptera, Aculeata, Formicidae). J. Hymenopt. Res. 2013, 30, 29. [Google Scholar] [CrossRef]
- Boudinot, B.E.; Fisher, B.L. A taxonomic revision of the Meranoplus F. Smith of Madagascar (Hymenoptera: Formicidae: Myrmicinae) with keys to species and diagnosis of the males. Zootaxa 2013, 3635, 301–339. [Google Scholar] [CrossRef] [PubMed]
- Seifert, B.; Kaufmann, B.; Fraysse, L. A taxonomic revision of the Palaearctic species of the ant genus Tapinoma Mayr 1861 (Hymenoptera: Formicidae). Zootaxa 2024, 5435, 1–74. [Google Scholar] [CrossRef]
- Csősz, S.; Loss, A.C.; Fisher, B.L. Taxonomic revision of the Malagasy Aphaenogaster swammerdami group (Hymenoptera: Formicidae). PeerJ 2021, 9, e10900. [Google Scholar] [CrossRef] [PubMed]
- Csősz, S.; Fisher, B. Taxonomic revision of the Malagasy Nesomyrmex madecassus species-group using a quantitative morphometric approach. ZooKeys 2016, 603, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Rakotonirina, J.C.; Csősz, S.; Fisher, B. Revision of the Malagasy Camponotus edmondi species group (Hymenoptera, Formicidae, Formicinae): Integrating qualitative morphology and multivariate morphometric analysis. ZooKeys 2016, 572, 81–154. [Google Scholar] [CrossRef] [PubMed]
- Rasoamanana, N.; Csősz, S.; Fisher, B.L. Taxonomic revision of imitating carpenter ants, Camponotus subgenus Myrmopytia (Hymenoptera, Formicidae) of Madagascar, using morphometry and qualitative traits. ZooKeys 2017, 681, 119–152. [Google Scholar] [CrossRef] [PubMed]
- Seifert, B. Revision of the Plagiolepis schmitzii group with description of Pl. invadens sp. nov.—A new invasive supercolonial species (Hymenoptera: Formicidae). Dtsch. Entomol. Z. 2020, 67, 183–196. [Google Scholar] [CrossRef]
- Seifert, B. Hypoponera ergatandria (Forel, 1893)—A cosmopolitan tramp species different from H. punctatissima (Roger, 1859) (Hymenoptera: Formicidae). Soil Org. 2013, 85, 189–201. [Google Scholar]
- Seifert, B. How to distinguish most similar insect species—Improving the stereomicroscopic and mathematical evaluation of external characters by example of ants. J. Appl. Entomol. 2002, 126, 445–454. [Google Scholar] [CrossRef]
- Ward, P.S.; Brady, S.G.; Fisher, B.L.; Schultz, T.R. The evolution of myrmicine ants: Phylogeny and biogeography of a hyperdiverse ant clade (Hymenoptera: Formicidae): Phylogeny and evolution of myrmicine ants. Syst. Entomol. 2015, 40, 61–81. [Google Scholar] [CrossRef]
- Bolton, B. A review of the Solenopsis genus-group and revision of Afrotropical Monomorium Mayr (Hymenoptera: Formicidae). Bull. Br. Mus. (Nat. Hist.) Entomol. 1987, 54, 263–452. [Google Scholar]
- Heterick, B.E. A revision of the Malagasy ants belonging to the genus Monomorium Mayr, 1855 (Hymenoptera: Formicidae). Proc. Calif. Acad. Sci. (Fourth Ser.) 2006, 59, 719–722. [Google Scholar]
- Ramamonjisoa, M.M.; Rasoamanana, N.; Fisher, B.L. Description of the male of Erromyrma Bolton & Fisher, 2016 (Hymenoptera, Formicidae). ZooKeys 2023, 1163, 61–77. [Google Scholar] [CrossRef]
- Ramamonjisoa, M.; Rasoamanana, N.; Fisher, B.L. Male-based key to the subfamilies and genera of Malagasy ants (Hymenoptera, Formicidae). ZooKeys, 2024; accepted. [Google Scholar]
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased recursive partitioning: A conditional inference framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Wolak, M. ICC: Facilitating estimation of the Intraclass Correlation Coefficient. 2022. Available online: https://cran.r-project.org/web/packages/ICC/index.html (accessed on 1 May 2023).
- Csősz, S.; Seifert, B.; Mikó, I.; Boudinot, B.E.; Borowiec, M.L.; Fisher, B.L.; Prebus, M.; Puniamoorthy, J.; Rakotonirina, J.; Rasoamanana, N.; et al. Insect morphometry is reproducible under average investigation standards. Ecol. Evol. 2021, 11, 547–559. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Jolliffe, I.T. (Ed.) Principal Component Analysis and Factor Analysis. In Principal Component Analysis; Springer: New York, NY, USA, 2002; pp. 150–166. ISBN 978-0-387-22440-4. [Google Scholar]
- Kruskal, J.B. Non-Metric Multidimensional Scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Mitteroecker, P.; Bookstein, F. Linear discrimination, ordination, and the visualization of selection gradients in modern morphometrics. Evol. Biol. 2011, 38, 100–114. [Google Scholar] [CrossRef]
- Hothorn, T.; Seibold, H.; Zeileis, A. Partykit: A Toolkit for Recursive Partitioning. 2023. Available online: https://cran.r-project.org/web/packages/partykit/index.html (accessed on 15 May 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; v.4.2.3; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 15 April 2023).
- Baer, B.; Boomsma, J.J. Mating biology of the leaf-cutting ants Atta colombica and A. cephalotes. J. Morphol. 2006, 267, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Rousseeuw, P.J. Silhouettes: A graphical aid to the interpretation and validation of cluster analysis. J. Comput. Appl. Math. 1987, 20, 53–65. [Google Scholar] [CrossRef]
- Seifert, B.; Ritz, M.; Csősz, S. Application of exploratory data analyses opens a new perspective in morphology-based alpha-taxonomy of eusocial organisms. Myrmecol. News 2014, 19, 1–15. [Google Scholar]
- Jolliffe, I.T. (Ed.) Outlier detection, influential observations, stability, sensitivity, and robust estimation of principal components. In Principal Component Analysis; Jolliffe, I.T. (Ed.) Springer: New York, NY, USA, 2002; pp. 232–268. ISBN 978-0-387-22440-4. [Google Scholar]
- Song, H. Species-specificity of male genitalia is characterized by shape, size, and complexity. Insect Syst. Evol. 2009, 40, 159–170. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation: Description | Character Definition | Figure |
|---|---|---|
| PaH: paramere height | Maximum height of the paramere in lateral view | Figure 1a |
| PaL: paramere length | Maximum length of the paramere in lateral view | Figure 1a |
| TeH: telomere height | Maximum height of the telomere in lateral view | Figure 1a |
| VoL: volsella length | Distance from the border of the basivolsella to the apex of the digitus in lateral view | Figure 1b |
| AeH: valviceps height | Maximum height of the valviceps in lateral view | Figure 1c |
| AeL: penisvalvae length | Maximum length from the apex of the valviceps to the valvura in lateral view | Figure 1c |
| SeL: valviceps denticles length | Distance between the first basal denticle to the first apical denticle of valviceps in lateral view | Figure 1c |
| VcL: valviceps length | Distance between the apex of the valviceps, not including the valvura in lateral view | Figure 1c |
| VcaH: apical height of the valviceps | Assessed vertically to the first apical denticle of the valviceps in lateral view | Figure 1c |
| VuL: valvura length | Maximum length of the valvura in lateral view | Figure 1c |
| Species | PaH | PaL | TeH | VoL | AeH |
| M. MG01 (n = 3) | 0.342 ± 0.015 [0.333, 0.359] | 0.365 ± 0.016 [0.347, 0.376] | 0.103 ± 0.014 [0.087, 0.111] | 0.204, 0.016 [0.194, 0.222] | 0.157 ± 0.004 [0.154, 0.162] |
| M. MG02 (n = 7) | 0.344 ± 0.021 [0.311, 0.374] | 0.357 ± 0.021 [0.335, 0.387] | 0.104 ± 0.012 [0.09, 0.118] | 0.188, 0.013 [0.167, 0.207] | 0.149 ± 0.007 [0.138, 0.158] |
| M. MG03 (n = 2) | 0.352 ± 0.009 [0.346, 0.358] | 0.426 ± 0.021 [0.4, 0.451] | 0.126 ± 0.002 [0.124, 0.127] | 0.162, 0.001 [0.162, 0.163] | 0.194 ± 0.014 [0.183, 0.204] |
| M. drm01 (n = 7) | 0.455 ± 0.008 [0.442, 0.468] | 0.561 ± 0.021 [0.519, 0.611] | 0.113, 0.016 [0.099, 0.146] | 0.277, 0.034 [0.236, 0.320] | 0.213 ± 0.009 [0.197, 0.224] |
| M. hanneli (n = 4) | 0.248 ± 0.008 [0.232, 0.267] | 0.342 ± 0.009 [0.335, 0.355] | 0.166 ± 0.008 [0.16, 0.176] | 0.248, 0.009 [0.454, 0.492] | 0.115 ± 0.001 [0.114, 0.115] |
| M. madecassum (n = 8) | 0.94 ± 0.017 [0.913, 0.959] | 1.032 ± 0.022 [1.009, 1.062] | 0.201 ± 0.018 [0.188, 0.232] | 0.477, 0.016 [0.454, 0.492] | 0.413 ± 0.037 [0.36, 0.482] |
| M. pharaonis (n = 4) | 0.789 ± 0.069 [0.728, 0.851] | 0.786 ± 0.03 [0.756, 0.816] | 0.174 ± 0.015 [0.159, 0.188] | 0.376, 0.018 [0.36, 0.392] | 0.385 ± 0.027 [0.363, 0.419] |
| M. termitobium_nr02 (n = 12) | 0.383 ± 0.033 [0.313, 0.434] | 0.446 ± 0.07 [0.339, 0.574] | 0.158 ± 0.02 [0.105, 0.181] | 0.253, 0.034 [0.201, 0.308] | 0.199 ± 0.029 [0.14, 0.238] |
| M. termitobium_nr03 (n = 4) | 0.323 ± 0.048 [0.311, 0.355] | 0.358 ± 0.003 [0.355, 0.36] | 0.103 ± 0.004 [0.1, 0.107] | 0.207, 0.002 [0.205, 0.21] | 0.152 ± 0.011 [0.145, 0.168] |
| M. termitobium_nr03b (n = 3) | 0.33 ± 0.022 [0.311, 0.355] | 0.348 ± 0.013 [0.335, 0.36] | 0.11 ± 0.013 [0.098, 0.125] | 0.203, 0.006 [0.199, 0.211] | 0.144 ± 0.005 [0.141, 0.15] |
| S. MG01 (n = 2) | 0.319 ± 0.022 [0.315, 0.323] | 0.364 ± 0.013 [0.36, 0.368] | 0.097 ± 0.007 [0.092, 0.103] | 0.191, 0.001 [0.19, 0.192] | 0.222 ± 0 [0.222, 0.223] |
| S. fisheri (n = 6) | 0.509 ± 0.04 [0.458, 0.555] | 0.564 ± 0.041 [0.53, 0.622] | 0.17 ± 0.022 [0.138, 0.193] | 0.35, 0.019 [0.328, 0.376] | 0.328 ± 0.029 [0.289, 0.376] |
| S. hildebrandti_nr01 (n = 6) | 0.274 ± 0.013 [0.252, 0.291] | 0.37 ± 0.012 [0.352, 0.382] | 0.113 ± 0.014 [0.101, 0.142] | 0.23, 0.009 [0.214, 0.24] | 0.235 ± 0.018 [0.216, 0.258] |
| S. hildebrandti_nr02 (n = 6) | 0.318 ± 0.011 [0.322, 0.378] | 0.329 ± 0.014 [0.312, 0.348] | 0.11 ± 0.006 [0.1, 0.117] | 0.212, 0.008 [0.236, 0.320] | 0.222 ± 0.016 [0.199, 0.248] |
| S. modesta (n = 6) | 0.347 ± 0.02 [0.322, 0.378] | 0.383 ± 0.015 [0.367, 0.41] | 0.098 ± 0.009 [0.1, 0.117] | 0.216, 0.014 [0.193, 0.23] | 0.227 ± 0.018 [0.201, 0.252] |
| Species | AeL | SeL | VcL | VcaH | VuL |
| M. MG01 (n = 3) | 0.187 ± 0.009 [0.182, 0.197] | 0.154 ± 0.015 [0.139, 0.169] | 0.166 ± 0.007 [0.159, 0.172] | 0.088 ± 0.007 [0.082, 0.095] | 0.113 ± 0.003 [0.109, 0.116] |
| M. MG02 (n = 7) | 0.188 ± 0.013 [0.174 ± 0.208] | 0.157 ± 0.003 [0.154, 0.16] | 0.167 ± 0.004 [0.161, 0.171] | 0.089 ± 0.008 [0.077, 0.102] | 0.115 ± 0.005 [0.106, 0.123] |
| M. MG03 (n = 2) | 0.229 ± 0.037 [0.203, 0.255] | 0.17 ± 0 [0.17, 0.17] | 0.177 ± 0.024 [0.16, 0.195] | 0.107 ± 0.007 [0.103, 0.112] | 0.14 ± 0.01 [0.133, 0.147] |
| M. drm01 (n = 7) | 0.235 ± 0.009 [0.221, 0.246] | 0.233 ± 0.013 [0.216, 0.255] | 0.232 ± 0.007 [[0.222, 0.246] | 0.133 ± 0.016 [0.118, 0.158] | 0.15, 0.004 [0.145, 0.156] |
| M. hanneli (n = 4) | 0.187 ± 0.002 [0.185, 0.188] | 0.106 ± 0.004 [0.102, 0.11] | 0.131 ± 0.003 [0.126, 0.134] | 0.091 ± 0.005 [0.085, 0.097] | 0.092 ± 0.002 [0.091, 0.095] |
| M. madecassum (n = 8) | 0.5 ± 0.034 [0.447, 0.554] | 0.35 ± 0.012 [0.329, 0.369] | 0.388 ± 0.029 [0.343, 0.422 | 0.204 ± 0.024 [0.174, 0.24] | 0.267 ± 0.026 [0.217, 0.306] |
| M. pharaonis (n = 4) | 0.527 ± 0.047 [0.488, 0.593] | 0.335 ± 0.022 [0.314, 0.365] | 0.366 ± 0.028 [0.337, 0.401] | 0.178 ± 0.01 [0.165, 0.187] | 0.255 ± 0.011 [0.241, 0.266] |
| M. termitobium_nr02 (n = 12) | 0.207 ± 0.031 [0.148, 0.242] | 0.191 ± 0.021 [0.15, 0.21] | 0.195 ± 0.03 [0.135, 0.229] | 0.106 ± 0.015 [0.084, 0.132] | 0.135 ± 0.017 [0.109, 0.165] |
| M. termitobium_nr03 (n = 4) | 0.178 ± 0.01 [0.167, 0.188] | 0.151 ± 0.01 [0.138, 0.159] | 0.151 ± 0.006 [0.144, 0.157] | 0.076 ± 0.006 [0.071, 0.084] | 0.111 ± 0.006 [0.106, 0.117] |
| M. termitobium_nr03b (n = 3) | 0.164 ± 0.008 [0.155, 0.17] | 0.152 ± 0.003 [0.149, 0.154] | 0.151, 0.002 [0.149, 0.152] | 0.097 ± 0.005 [0.093, 0.103] | 0.112 ± 0.005 [0.107, 0.116] |
| S. MG01 (n = 2) | 0.368 ± 0.012 [0.36, 0.377] | 0.382 ± 0.02 [0.368, 0.396] | 0.286, 0.019 [0.272, 0.3] | 0.154, 0.007 0.149, 0.158] | 0.087 ± 0.01 [0.08, 0.094] |
| S. fisheri (n = 6) | 0.49 ± 0.045 [0.432, 0.566] | 0.495 ± 0.033 [0.468, 0.555] | 0.391 ± 0.021 [0.369, 0.426] | 0.134 ± 0.015 [0.113, 0.15] | 0.142 ± 0.018 [0.121, 0.173] |
| S. hildebrandti_nr01 (n = 6) | 0.305 ± 0.023 [0.285, 0.342] | 0.313 ± 0.012 [0.296, 0.333] | 0.263 ± 0.009 [0.255, 0.278] | 0.091 ± 0.01 [0.074, 0.101] | 0.093 ± 0.019 [0.078, 0.124] |
| S. hildebrandti_nr02 (n = 6) | 0.281 ± 0.018 [0.254, 0.298] | 0.293 ± 0.01 [0.283, 0.31] | 0.249 ± 0.012 [0.239, 0.265] | 0.087 ± 0.005 [0.082, 0.097] | 0.088 ± 0.007 [0.075, 0.093] |
| S. modesta (n = 6) | 0.37 ± 0.013 [0.351, 0.389] | 0.375 ± 0.011 [0.366, 0.395] | 0.292 ± 0.014 [0.282, 0.317] | 0.129 ± 0.011 [0.112, 0.144] | 0.089 ± 0.007 [0.075, 0.095] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasoarimalala, N.F.; Ramiadantsoa, T.; Rakotonirina, J.C.; Fisher, B.L. Linear Morphometry of Male Genitalia Distinguishes the Ant Genera Monomorium and Syllophopsis (Hymenoptera: Formicidae) in Madagascar. Insects 2024, 15, 605. https://doi.org/10.3390/insects15080605
Rasoarimalala NF, Ramiadantsoa T, Rakotonirina JC, Fisher BL. Linear Morphometry of Male Genitalia Distinguishes the Ant Genera Monomorium and Syllophopsis (Hymenoptera: Formicidae) in Madagascar. Insects. 2024; 15(8):605. https://doi.org/10.3390/insects15080605
Chicago/Turabian StyleRasoarimalala, Nomena F., Tanjona Ramiadantsoa, Jean Claude Rakotonirina, and Brian L. Fisher. 2024. "Linear Morphometry of Male Genitalia Distinguishes the Ant Genera Monomorium and Syllophopsis (Hymenoptera: Formicidae) in Madagascar" Insects 15, no. 8: 605. https://doi.org/10.3390/insects15080605