Investigating the Influence of Anthropogenic Activities on Behavioral Changes of an Orb Web Spider (Neoscona vigilans)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Spider Identification
2.2. Data Analysis
2.3. Web Anomalies
2.4. Statistical Analysis
3. Results
3.1. Anomalies
3.2. Relationship between Spider Body Measurements and Web Properties
3.3. Relationship between Web Characteristics and Distance from the Road
3.4. Relationship between Web Anomalies and Spider Body Measures
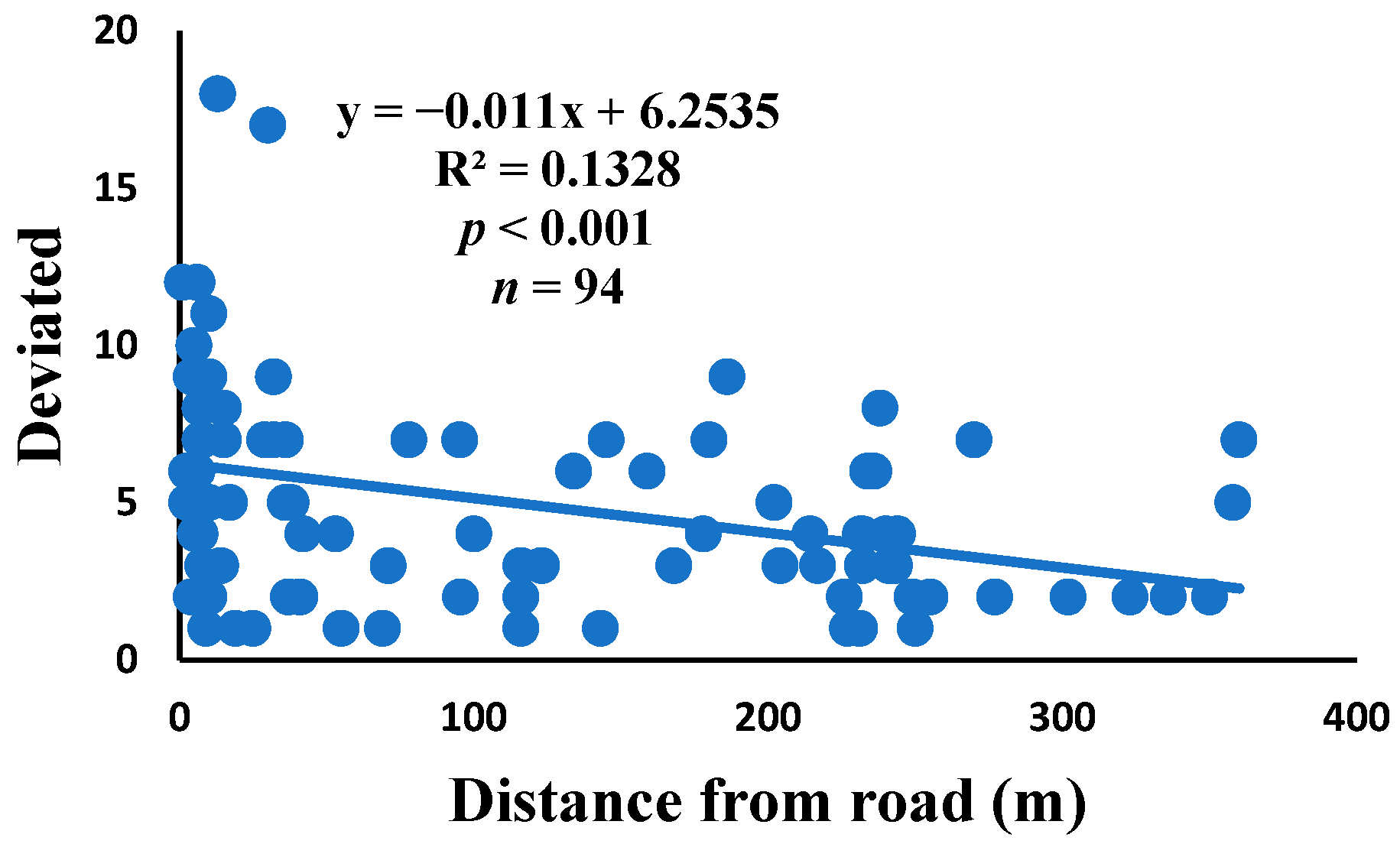
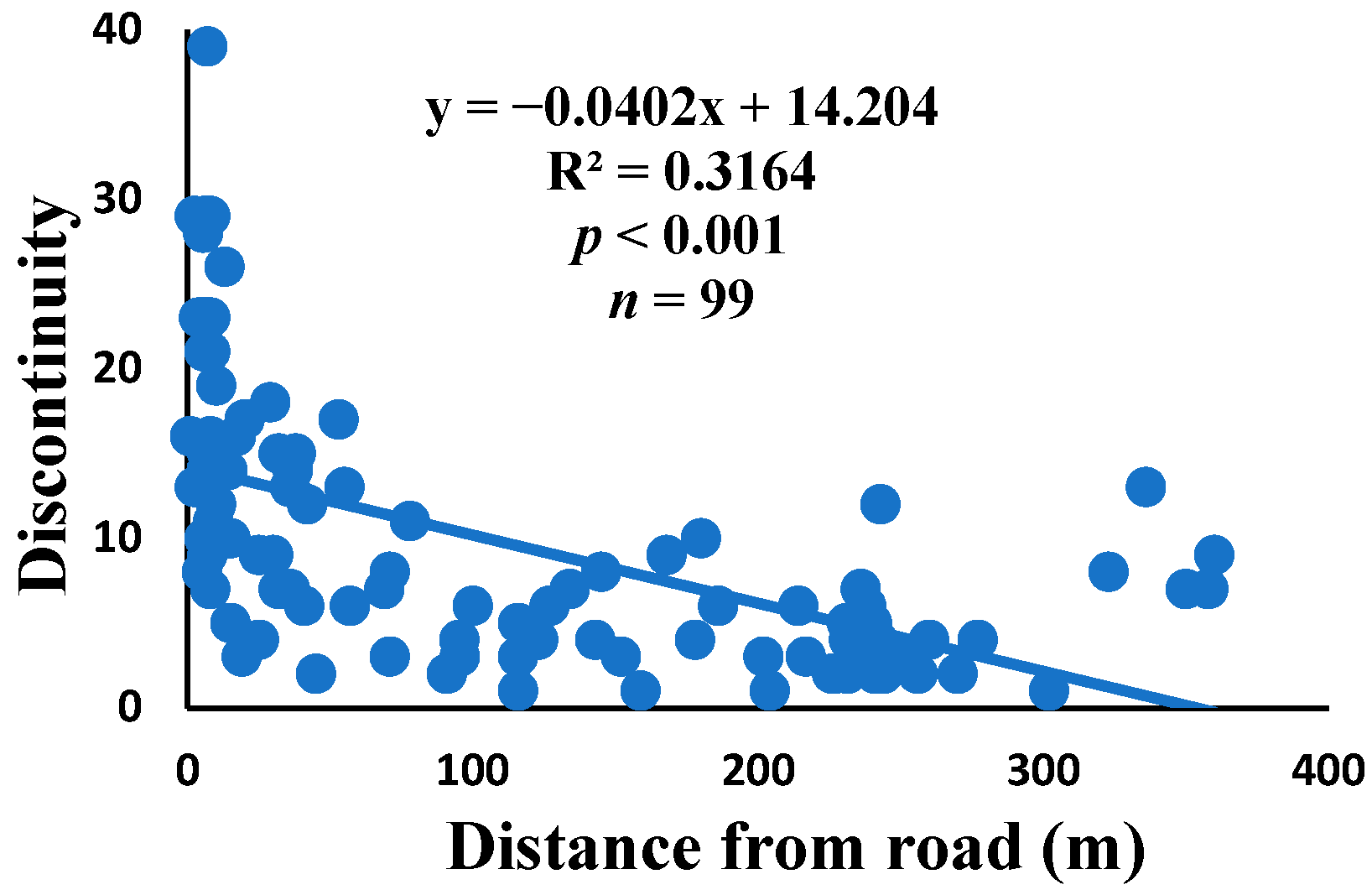
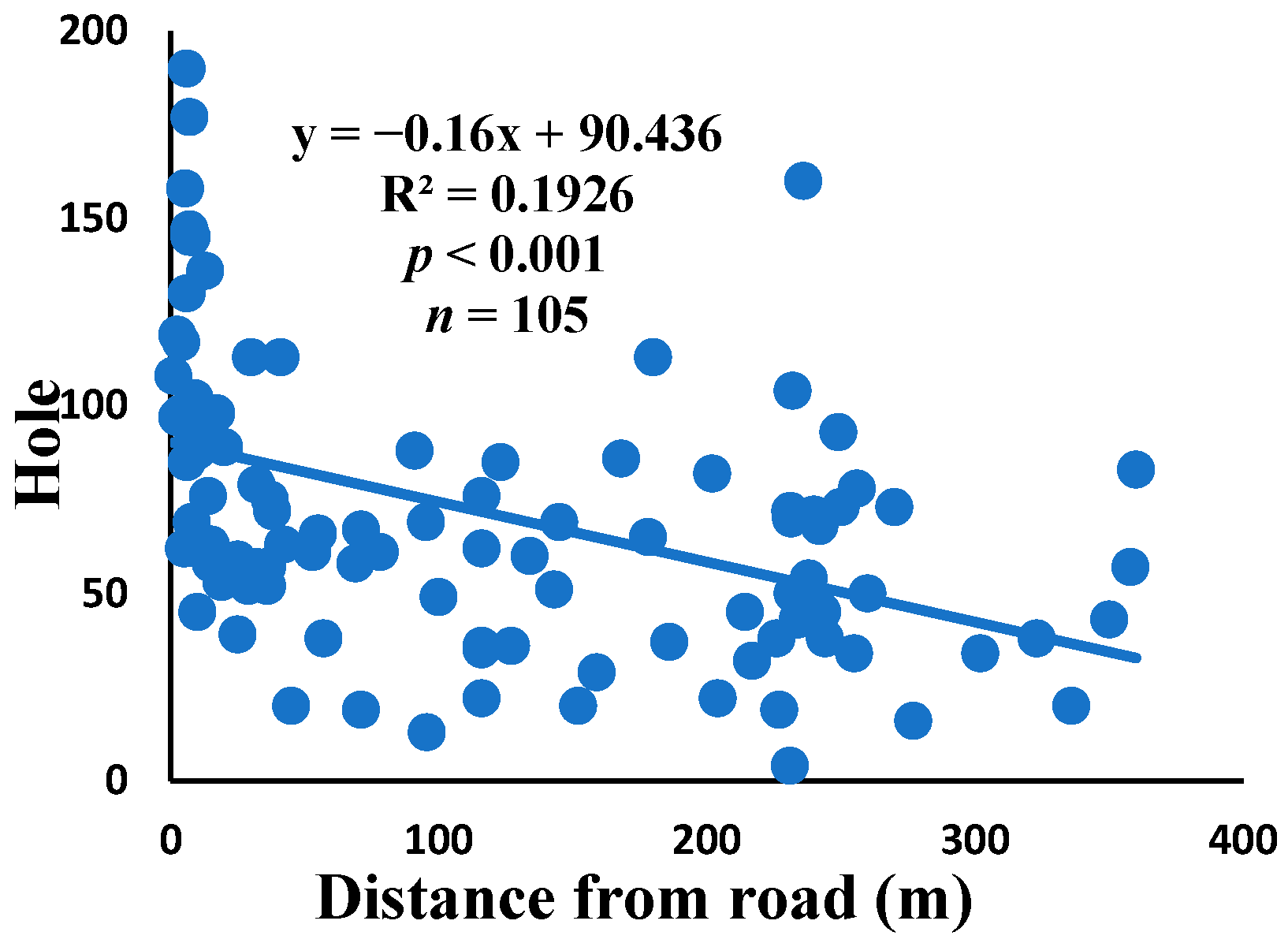
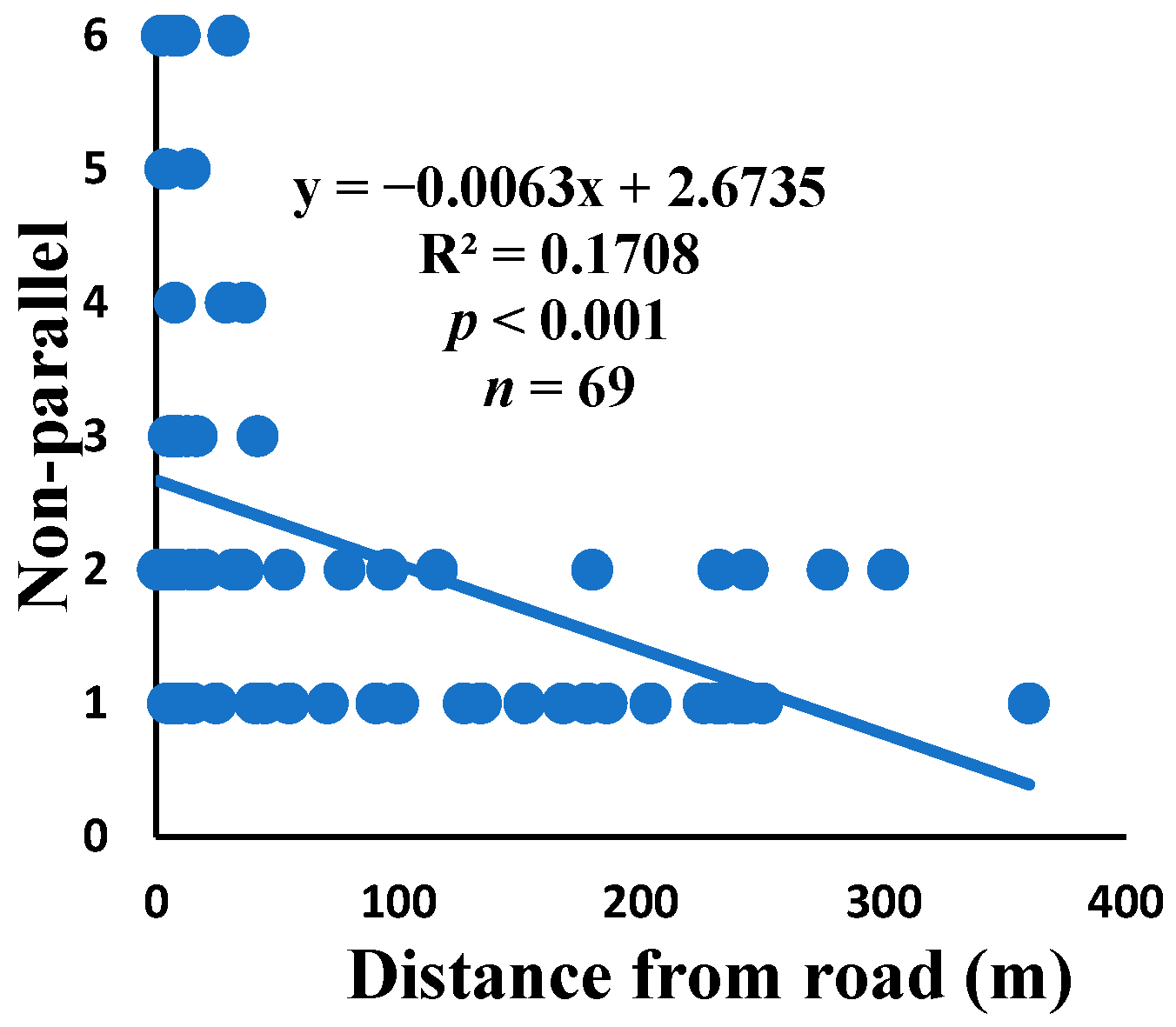
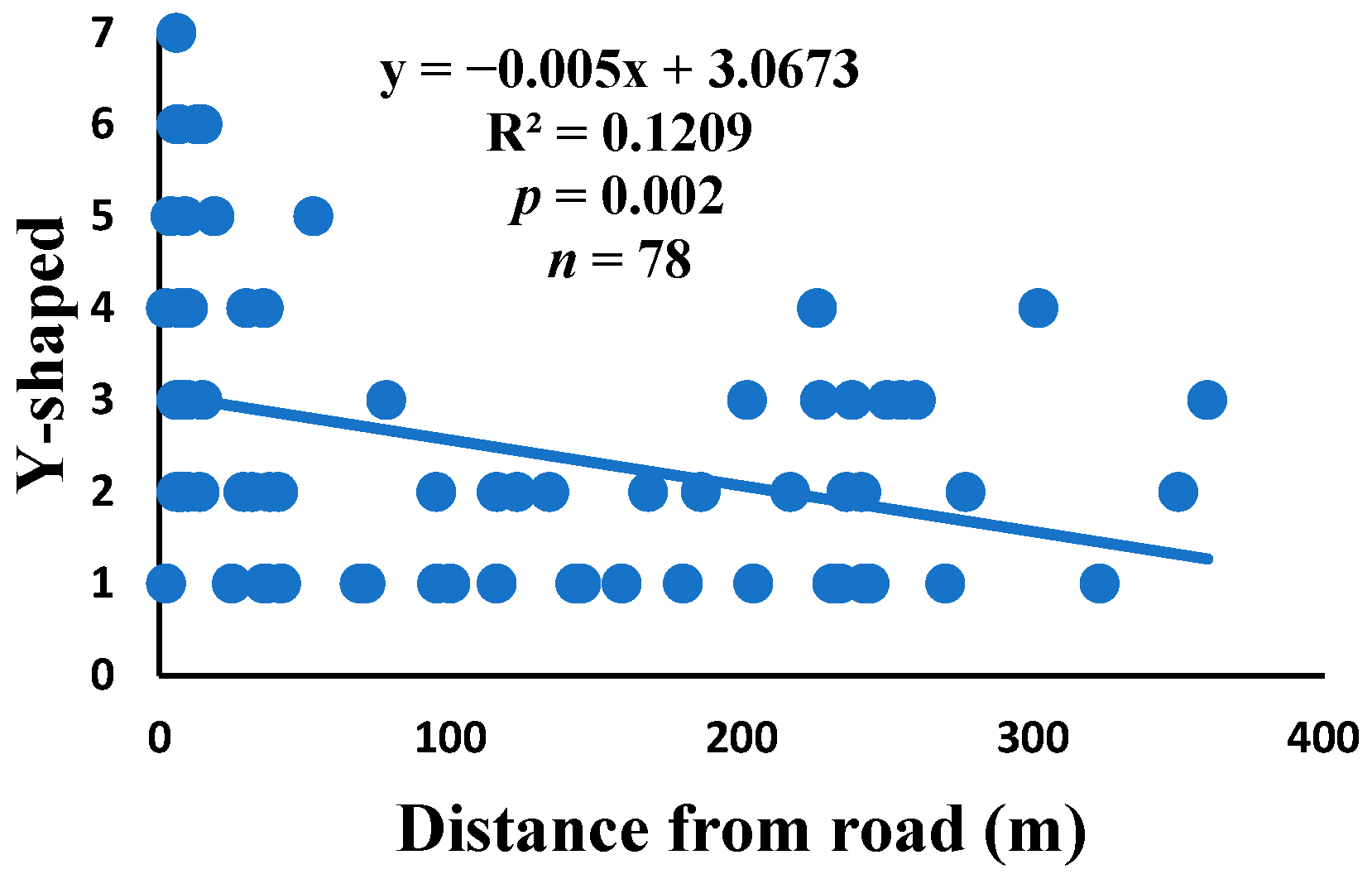
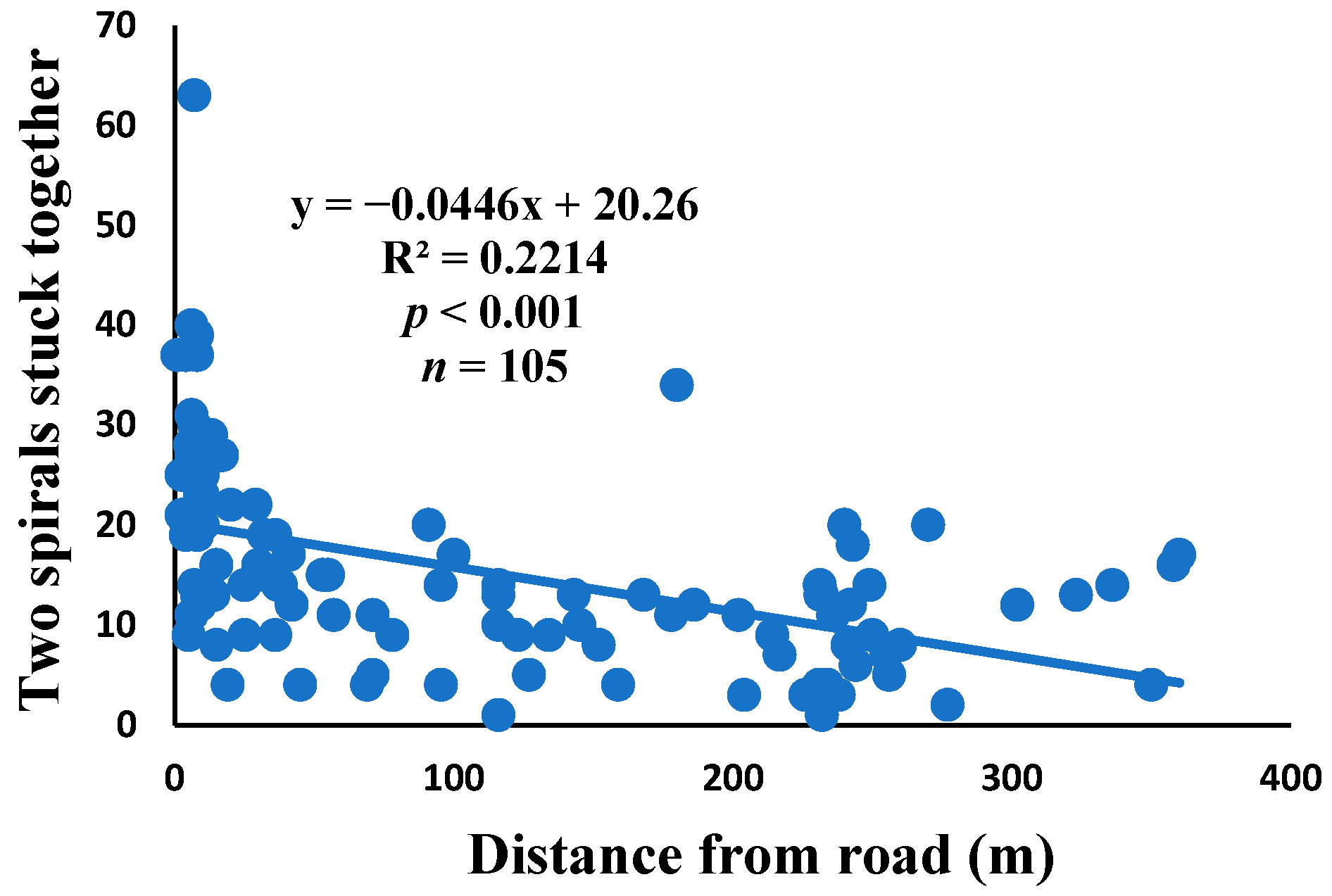
3.5. Relationship between Web Anomalies and Distance from the Road
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rybak, J.; Olejniczak, T. Accumulation of polycyclic aromatic hydrocarbons (PAHs) on the spider webs in the vicinity of road traffic emissions. Environ. Sci. Pollut. Res. 2014, 21, 2313–2324. [Google Scholar] [CrossRef]
- Ambily, C.B.; Antony, A. Diversity and distribution of spiders in agro ecosystem of Ernakulum, District, Kerala. J. Zool. Stud. 2016, 3, 73–77. [Google Scholar]
- Chaubey, S.N. Studies on habit and habitat, external morphology, feeding capacity and prey preference of garden jumping spider, Opisthoncus SPECIES. Indian J. Sci. Res. 2019, 10, 29–34. [Google Scholar] [CrossRef]
- Mishra, A.; Rastogi, N. Unraveling the roles of solitary and social web-making spiders in perennial ecosystems: Influence on pests and beneficials. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 567–576. [Google Scholar] [CrossRef]
- Khan, A.A.; Kundoo, A.A.; Khan, Z.H.; Hussain, K. Identification of potential and suitable natural enemies of arthropod pests for conservation biological control in vegetable ecosystem of Kashmir. J. Entomol. Zool. Stud. 2020, 8, 2251–2255. [Google Scholar]
- Ghazanfar, M.; Hussain, M.; Hashim, M.; Fahid, A.M. Checklist of spider (Araneae) fauna of Pakistan: A review. J. Entomol. Zool. Stud. 2016, 4, 245–256. [Google Scholar]
- Lawania, K.K.; Mathur, P. Study on the pattern and archetecture of spider’s web with special reference to seasonal abundance in eastern region of Rajasthan. India J. Environ. Sci. Toxicol. Food Tech. 2015, 9, 1–9. [Google Scholar]
- Nyffeler, M.; Bonte, D. Where have all the spiders gone? Observations of a dramatic population density decline in the once very abundant garden spider, Araneus diadematus (Araneae: Araneidae), in the Swiss Midland. Insects 2020, 11, 248. [Google Scholar] [CrossRef]
- Solanki, R.; Kumar, D. Spiders (Araneae) from five major agro-ecosystems of Jambughoda village, Panchmahal district, Gujarat, India. Int. J. Sci. Res. 2015, 4, 958–961. [Google Scholar]
- Lv, H.; Li, H.; Qiu, Z.; Zhang, F.; Song, J. Assessment of pedestrian exposure and deposition of PM10, PM2. 5 and ultrafine particles at an urban roadside: A case study of Xi’an, China. Atmos. Pollut. 2021, 12, 112–121. [Google Scholar] [CrossRef]
- Hagler, G.S.W.; Baldauf, R.W.; Thoma, E.D.; Long, T.R.; Snow, R.F.; Kinsey, J.S.; Oudejans, L.; Gullett, B.K. Ultrafine particles near a major roadway in Raleigh, North Carolina: Downwind attenuation and correlation with traffic-related pollutants. Atmos. Environ. 2009, 43, 1229–1234. [Google Scholar] [CrossRef]
- Jinsart, W.; Kaewmanee, C.; Inoue, M.; Hara, K.; Hasegawa, S.; Karita, K.; Tamura, K.; Yano, E. Driver exposure to particulate matter in Bangkok. J. Air Waste Manag. Assoc. 2012, 62, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Su, I.; Buehler, M.J. Mesomechanics of a three-dimensional spider web. J. Mech. Phys. Solids 2020, 144, 104096. [Google Scholar] [CrossRef]
- Jiang, Y.; Nayeb-Hashemi, H. Dynamic response of spider orb webs subject to prey impact. Int. J. Mech. Sci. 2020, 186, 105899. [Google Scholar] [CrossRef]
- Rutkowski, R.; Justyna, R.; Wioletta, R.; Maciej, B.; Katarzyna, P.; Izabela, J. Mutagenicity of indoor air pollutants adsorbed on spider webs. Ecotoxicol. Environ. Saf. 2019, 171, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Rybak, J. Possible use of spider webs for the indication of organic road pollutants. J. Ecol. Eng. 2014, 15, 39–45. [Google Scholar]
- Zhou, T.; Luo, X.; Hou, Y.; Xiang, Y.; Peng, S. Quantifying the effects of road width on roadside vegetation and soil conditions in forests. Landsc. Ecol. 2020, 35, 69–81. [Google Scholar] [CrossRef]
- Stojanowska, A.; Mach, T.; Olszowski, T.; Bihałowicz, J.S.; Górka, M.; Rybak, J.; Świsłowski, P. Air pollution research based on spider web and parallel continuous particulate monitoring—A comparison study coupled with identification of sources. Minerals 2021, 11, 812. [Google Scholar] [CrossRef]
- Ramírez, O.; de la Campa AM, S.; Sánchez-Rodas, D.; de la Rosa, J.D. Hazardous trace elements in thoracic fraction of airborne particulate matter: Assessment of temporal variations, sources, and health risks in a megacity. Sci. Total Environ. 2020, 710, 136344. [Google Scholar] [CrossRef]
- Cranford, S.W.; Tarakanova, A.; Pugno, N.M.; Buehler, M.J. Nonlinear material behaviour of spider silk yields robust webs. Nature 2012, 482, 72–76. [Google Scholar] [CrossRef]
- Turner, J.; Vollrath, F.; Hesselberg, T. Wind speed affects prey-catching behaviour in an orb web spider. Naturwissenschaften 2011, 98, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Blamires, S.J.; Chao, Y.C.; Liao, C.P.; Tso, I.M. Multiple prey cues induce foraging flexibility in a trap-building predator. Anim. Behav. 2011, 81, 955–961. [Google Scholar] [CrossRef]
- Bosia, F.; Dal Poggetto, V.F.; Gliozzi, A.S.; Greco, G.; Lott, M.; Miniaci, M.; Pugno, N.M. Optimized structures for vibration attenuation and sound control in nature: A review. Matter 2022, 5, 3311–3340. [Google Scholar] [CrossRef]
- Agnarsson, I.; Coddington, J.A.; Kuntner, M. Systematics: Progress in the study of spider diversity and evolution. In Spider Research in the 21st Century: Trends and Perspectives; Siri Scientific Press: Manchester, UK, 2013. [Google Scholar]
- Toscani, C.; Leborgne, R.; Pasquet, A. Behavioural analysis of web building anomalies in the orb-weaving spider Zygiella x-notata (Araneae, Araneidae). Arachnol. Mitteilungen 2012, 43, 79–83. [Google Scholar] [CrossRef]
- Harmer, A.M.; Blackledge, T.A.; Madin, J.S.; Herberstein, M.E. High-performance spider webs: Integrating biomechanics, ecology and behaviour. J. R. Soc. Interface 2011, 8, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Pasquet, A.; Marchal, J.; Anotaux, M.; Leborgne, R. Imperfections in perfect architecture: The orb web of spiders. Eur. J. Entomol. 2013, 110, 493–500. [Google Scholar] [CrossRef]
- Anotaux, M.; Toscani, C.; Leborgne, R.; Châline, N.; Pasquet, A. Aging and foraging efficiency in an orb-web spider. J. Ethol. 2014, 32, 155–163. [Google Scholar] [CrossRef]
- Pasquet, A.; Marchal, J.; Anotaux, M.; Leborgne, R. Does building activity influence web construction and web characteristics in the orb-web spider Zygiella x-notata (Araneae, Araneidae)? Zool. Stud. 2014, 53, 11. [Google Scholar] [CrossRef]
- Pasquet, A.; Toscani, C.; Anotaux, M. Influence of aging on brain and web characteristics of an orb web spider. J. Ethol. 2018, 36, 85–91. [Google Scholar] [CrossRef]
- Mohammadi, A.; Fatemizadeh, F. Quantifying landscape degradation following construction of a highway using landscape metrics in southern Iran. Front. Ecol. Evol. 2021, 9, 721313. [Google Scholar] [CrossRef]
- Siemers, B.M.; Schaub, A. Hunting at the highway: Traffic noise reduces foraging efficiency in acoustic predators. Proc. R. Soc. B Biol. Sci. 2011, 278, 1646–1652. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Zhao, Z.; Li, S. Is phenotypic evolution affected by spiders’ construction behaviors? Syst. Biol. 2023, 72, 319–340. [Google Scholar] [CrossRef]
- Caleb, J.T. Spiders (Arachnida: Araneae) from the vicinity of Araabath Lake, Chennai, India. J. Threat. Taxa 2020, 12, 15186–15193. [Google Scholar] [CrossRef]
- Tikader, B.K. Family Araneidae (= Argiopidae), typical orbweavers. Fauna India 1982, 2, 1–293. [Google Scholar]
- Raychaudhuri, D.; Saha, S. Spiders (Araneae: Arachnida) of Reserve Forests of Dooars: Gorumara National Park, Chapramari Wildlife Sanctuary and Mahananda Wildlife Sanctuary. World Sci. News 2015, 20, 1–336. [Google Scholar]
- Herberstein, M.E.; Tso, I.M. Evaluation of formulae to estimate the capture area and mesh height of orb webs (Araneoidea, Araneae). J. Arachnol. 2000, 28, 180–184. [Google Scholar] [CrossRef]
- Prokop, P.; Grygláková, D. Factors affecting the foraging success of the wasp-like spider Argiope bruennichi (Araneae): Role of web design. Biologia 2005, 60, 165–169. [Google Scholar]
- Blackledge, T.A.; Gillespie, R.G. Estimation of capture areas of spider orb webs in relation to asymmetry. J. Arachnol. 2002, 30, 70–77. [Google Scholar] [CrossRef]
- Hesselberg, T. Ontogenetic changes in web design in two orb-web spiders. Ethology 2010, 116, 535–545. [Google Scholar] [CrossRef]
- Rybak, J. Spider webs as pollution indicators. In Advances in Environmental Research; Nova Science Publishers Inc.: New York, NY, USA, 2016; Volume 49. [Google Scholar]
- Davis, A.K.; Stewart, K.; Phelan, C.; Schultz, A. How Urban-Tolerant Are They? Testing Prey–Capture Behavior of Introduced Jorō Spiders (Trichonephila clavata) Next to Busy Roads. Arthropoda 2024, 2, 55–65. [Google Scholar] [CrossRef]
- Michalko, R.; Košulič, O.; Martinek, P.; Birkhofer, K. Disturbance by invasive pathogenic fungus alters arthropod predator–prey food-webs in ash plantations. J. Anim. Ecol. 2021, 90, 2213–2226. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, J.; Wang, Z.; Zhibin, W.; Tian, J.; Chen, J.; Lan, J. Transcriptome changes reveal the toxic mechanism of cadmium and lead combined exposure on silk production and web-weaving behavior of spider A. ventricosus. Environ. Sci. Technol. 2023, 57, 14917–14928. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.A.; Dyakonova, V.; Rillich, J.; Schildberger, K. Octopamine and experience-dependent modulation of aggression in crickets. J. Neurosci. 2005, 25, 1431–1441. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.W. The life cycle, habitat and variation in selected web parameters in the spider, Nephila clavipes Koch (Araneidae). Am. Midl. Nat. 1977, 98, 95–108. [Google Scholar] [CrossRef]
- Zschokke, S.; Vollrath, F. Web construction patterns in a range of orb-weaving spiders (Araneae). Eur. J. Entomol. 1995, 92, 523–541. [Google Scholar]
- Johnson, J.C.; Garver, E.; Martin, T. Black widows on an urban heat island: Extreme heat affects spider development and behaviour from egg to adulthood. Anim. Behav. 2020, 167, 77–84. [Google Scholar] [CrossRef]
- Dahirel, M.; De Cock, M.; Vantieghem, P.; Bonte, D. Urbanization-driven changes in web building and body size in an orb web spider. J. Anim. Ecol. 2019, 88, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Marko, V.; Bogya, S.; Kondorosy, E.; Blommers, L.H. Side effects of kaolin particle films on apple orchard bug, beetle and spider communities. Int. J. Pest Manag. 2010, 56, 189–199. [Google Scholar] [CrossRef]
- Martin, A.E.; Pervin, E.; Graham, S.L.; Henry, M.; Fahrig, L. Abundance of aerially-dispersing spiders declines with increasing road traffic. Ecoscience 2019, 26, 383–388. [Google Scholar] [CrossRef]
- Witt, P.N.; Baum, R. Changes in orb webs of spiders during growth (Araneus Diadematus Clerck and Neoscona Vertebrata Mc Cook). Behaviour 1960, 16, 309–318. [Google Scholar]
- Sensenig, A.; Agnarsson, I.; Blackledge, T.A. Behavioural and biomaterial coevolution in spider orb webs. J. Evol. Biol. 2010, 23, 1839–1856. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.G.; Kusahara, M.; Maezono, Y.; Miyashita, T. Adjustment of web-building initiation to high humidity: A constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. Sci. Nat. 2014, 101, 587–593. [Google Scholar] [CrossRef]
- Blamires, S.J.; Sellers, W.I. Modelling temperature and humidity effects on web performance: Implications for predicting orb-web spider (Argiope spp.) foraging under Australian climate change scenarios. Conserv. Physiol. 2019, 7, coz083. [Google Scholar] [CrossRef] [PubMed]
- Tew, E.R.; Adamson, A.; Hesselberg, T. The web repair behaviour of an orb spider. Anim. Behav. 2015, 103, 137–146. [Google Scholar] [CrossRef]
- Rhisiart, A.A.; Vollrath, F. Design features of the orb web of the spider, Araneus diadematus. Behav. Ecol. 1994, 5, 280–287. [Google Scholar] [CrossRef]
- Sensenig, A.T.; Agnarsson, I.; Blackledge, T.A. Adult spiders use tougher silk: Ontogenetic changes in web architecture and silk biomechanics in the orb-weaver spider. J. Zoo. 2011, 285, 28–38. [Google Scholar] [CrossRef]
- Sanphui, P.; Dey, S.; Mukherjee, D.; Saha, S. Affirming length of body or part/s and weight there of as a determinant of capture area of spider web. World Sci. News 2017, 71, 105–117. [Google Scholar]
- Butt, A.; Alam, I.; Naz, R. Variations in web architecture of Argiope trifasciata (Araneae, Araneidae) and its relationship with body parameters and entangled prey. Pak. J. Zool. 2017, 49, 855–860. [Google Scholar] [CrossRef]
- Anotaux, M.; Marchal, J.; Châline, N.; Desquilbet, L.; Leborgne, R.; Gilbert, C.; Pasquet, A. Ageing alters spider orb-web construction. Anim. Behav. 2012, 84, 1113–1121. [Google Scholar] [CrossRef]
- Opell, B.D. Redesigning spider webs: Stickiness, capture area and the evolution of modern orb-webs. Evol. Ecol. Res. 1999, 1, 503–516. [Google Scholar]
- Sensenig, A.T.; Kelly, S.P.; Lorentz, K.A.; Lesher, B.; Blackledge, T.A. Mechanical performance of spider orb webs is tuned for high-speed prey. J. Exp. Biol. 2013, 216, 3388–3394. [Google Scholar] [CrossRef] [PubMed]
- Rao, D.; Fernandez, O.C.; Castañeda-Barbosa, E.; Díaz-Fleischer, F. Reverse positional orientation in a neotropical orb-web spider, Verrucosa arenata. Sci. Nat. 2011, 98, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Kuntner, M.; Gregorič, M.; Li, D. Mass predicts web asymmetry in Nephila spiders. Sci. Nat. 2010, 97, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S.; Countryman, S.; Cushing, P.E. Spiders in space—Orb-web-related behaviour in zero gravity. Sci. Nat. 2021, 108, 1. [Google Scholar] [CrossRef]
- Tahir, H.M.; Butt, A.; Alam, I. Relationship of web characteristics and body measures of Leucauge decorata (Araneae: Tetragnathidae). Pak. J. Zool. 2010, 42, 261–265. [Google Scholar]
- Herberstein, M.E.; Heiling, A.M. Does mesh height influence prey length in orb-web spiders (Araneae)? EJE 2013, 95, 367–371. [Google Scholar]
- Watanabe, T. Effects of web design on the prey capture efficiency of the uloborid spider Octonoba sybotides under abundant and limited prey conditions. Zool. Sci. 2001, 18, 585–590. [Google Scholar] [CrossRef]
- Aoyanagi, Y.; Okumura, K. Simple model for the mechanics of spider webs. Phys. Rev. Lett. 2010, 104, 038102. [Google Scholar] [CrossRef]
- Krafft, B.; Cookson, L.J. The role of silk in the behaviour and sociality of spiders. Psyche J. Entomol. 2012, 529564. [Google Scholar] [CrossRef]
- Foellmer, M.W.; Marson, M.; Moya-Laraño, J. Running performance as a function of body size, leg length, and angle of incline in male orb-web spiders, Argiope aurantia. Evol. Ecol. Res. 2011, 13, 513–526. [Google Scholar]
- Foelix, R.; Hebets, E. Sensory Biology of Whip Spiders (Arachnida, Amblypygi). 2001. Available online: https://digitalcommons.unl.edu/bioscihebets/32/ (accessed on 1 April 2024).
- Papadopoulos, N.T.; Carey, J.R.; Katsoyannos, B.I.; Kouloussis, N.A.; Müller, H.G.; Liu, X. Supine behaviour predicts the time to death in male Mediterranean fruitflies (Ceratitis capitata). Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 1633–1637. [Google Scholar] [CrossRef]
- Heiling, A.M.; Herberstein, M.E. Interpretations of orb-web variability: A review of past and current ideas. Ekol.(Bratisl.)/Ecol.(Bratisl.) 2000, 19, 97–106. [Google Scholar]
- Samu, F.; Vollrath, F. Spider orb web as bioassay for pesticide side effects. Entomol. Exp. Appl. 1992, 62, 117–124. [Google Scholar] [CrossRef]
- Osaki, S.; Osaki, M. Evolution of spiders from nocturnal to diurnal gave spider silks mechanical resistance against UV irradiation. Polym. J. 2011, 43, 200–204. [Google Scholar] [CrossRef]
- Wilder, S.M. Spider nutrition: An integrative perspective. In Advances in Insect Physiology; Academic Press: Cambridge, MA, USA, 2011; Volume 40, pp. 87–136. [Google Scholar]
- Tietjen, W.J.; Cady, A.B. Sublethal exposure to a neurotoxic pesticide affects activity rhythms and patterns of four spider species. J. Arachnol. 2007, 35, 396–406. [Google Scholar] [CrossRef]
- Yeoman, M.S.; Faragher, R.G.A. Ageing and the nervous system: Insights from studies on invertebrates. Biogerontology 2001, 2, 85–97. [Google Scholar] [CrossRef]
- Landolfa, M.A.; Barth, F.G. Vibrations in the orb web of the spider Nephila clavipes: Cues for discrimination and orientation. J. Comp. Physiol. A 1996, 179, 493–508. [Google Scholar] [CrossRef]
- Mulder, T.; Wilkins, L.; Mortimer, B.; Vollrath, F. Dynamic environments do not appear to constrain spider web building behaviour. Sci. Nat. 2021, 108, 20. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Evol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Hesselberg, T. Exploration behaviour and behavioural flexibility in orb-web spiders: A review. Curr. Zool. 2015, 61, 313–327. [Google Scholar] [CrossRef]
- Tew, N.; Hesselberg, T. The effect of wind exposure on the web characteristics of a tetragnathid orb spider. J. Insect Behav. 2017, 30, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.B.; Candolin, U. Behavioral responses to changing environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef]
- Johnson, J.C.; Urcuyo, J.; Moen, C.; Stevens, D.R. Urban heat island conditions experienced by the Western black widow spider (Latrodectus hesperus): Extreme heat slows development but results in behavioral accommodations. PLoS ONE 2019, 14, e0220153. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S. Nomenclature of the orb-web. J. Arachnol. 1999, 27, 542–546. [Google Scholar]
- Jyoti, J.; Kumar, A.; Lakhani, P.; Kumar, N.; Bhushan, B. Structural properties and their influence on the prey retention in the spider web. Philos. Trans. R. Soc. A 2019, 377, 20180271. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S. Form and function of the orb-web. Eur. Arachnol. 2000, 19, 99. [Google Scholar]
- Heard, T.A.; Hendrikz, J.K. Factors influencing flight activity of colonies of the stingless bee Trigona-carbonaria (Hymenoptera, Apidae). Aust. J. Zool. 1993, 41, 343–353. [Google Scholar] [CrossRef]
- Namazi, H.R. The complexity based analysis of the correlation between spider’s brain signal and web. ARC J. Neurosci. 2017, 2, 38–44. [Google Scholar]




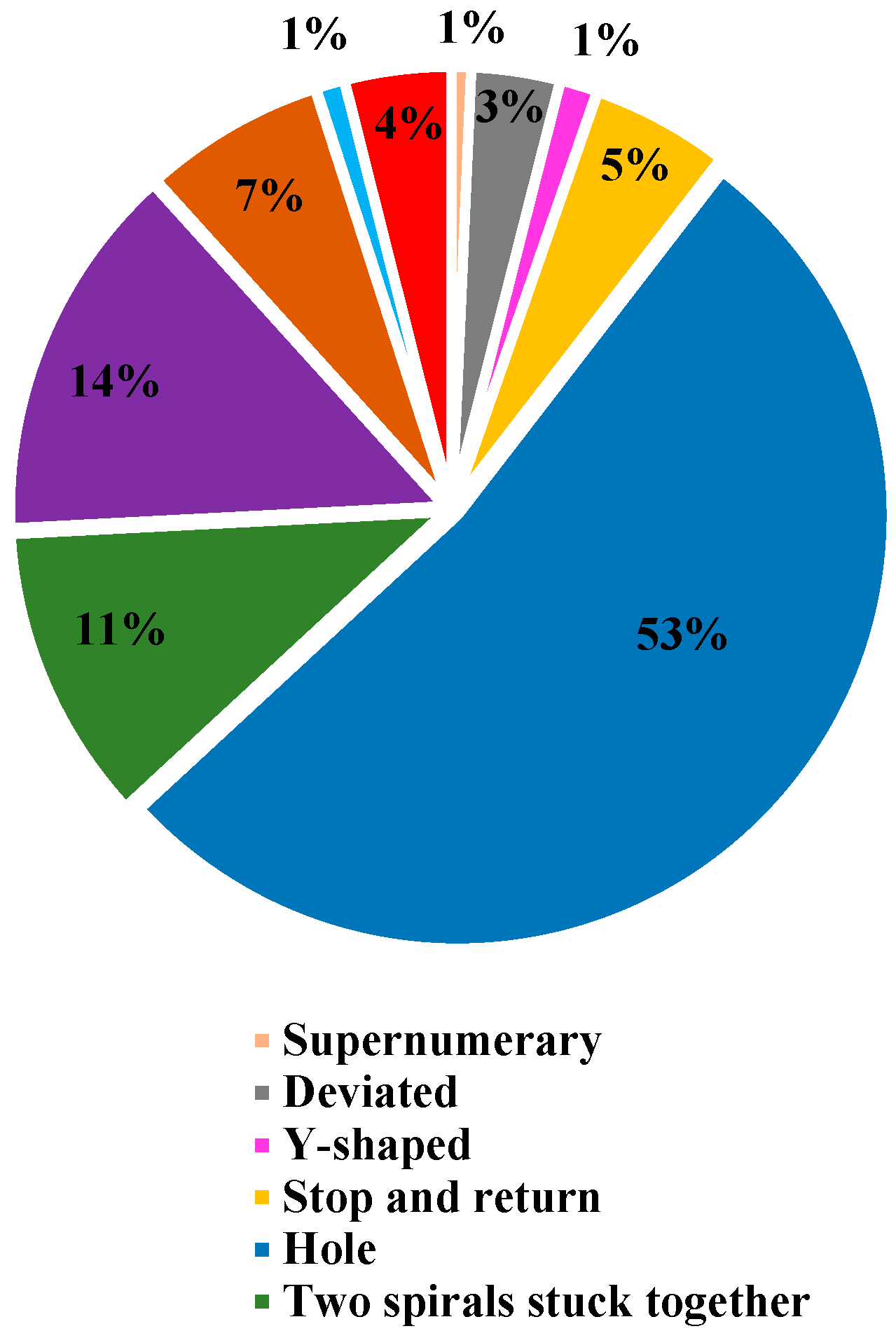


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anomalies | Percentage of Webs in Which an Anomaly Was Present | Percentage of Anomalies | The Average Number of Anomalies per Web | Total Anomalies | n |
|---|---|---|---|---|---|
| Hole | 100% | 52.7% | 72.1 | 7567 | 105 |
| Discontinuity | 94.3% | 6.8% | 9.8 | 970 | 99 |
| Supernumerary | 52.4% | 0.7% | 1.8 | 100 | 55 |
| Two spirals stuck together | 100% | 11.1% | 15.1 | 1590 | 105 |
| Nonparallel | 65.7% | 1% | 2.2 | 149 | 69 |
| Deviated | 89.5% | 3.3% | 5.0 | 470 | 94 |
| Zigzag | 85.7% | 3.9% | 6.3 | 564 | 90 |
| Y-shaped | 74.3% | 1.4% | 2.5 | 196 | 78 |
| More than two spirals stuck together | 99% | 14.1% | 19.5 | 2026 | 104 |
| Stop and return | 99% | 5.1% | 7.1 | 737 | 104 |
| Total | 100% | 14,369 |
| Web Characteristic | Fourth Leg Length (mm) | Carapace Width (mm) | Body Length (mm) | |||
|---|---|---|---|---|---|---|
| R2 | p | R2 | p | R2 | p | |
| Horizontal web diameter (cm) | 0.145 | <0.001 * | 0.086 | 0.002 | 0.120 | <0.001 * |
| Mesh size (mm) | 0.279 | <0.001 * | 0.237 | <0.001 * | 0.181 | <0.001 * |
| Upper radii | 0.052 | 0.019 | 0.049 | 0.024 | 0.044 | 0.032 |
| Anchor points | 0.022 | 0.136 | 0.048 | 0.026 | 0.044 | 0.032 |
| Capture area (cm2) | 0.347 | <0.001 * | 0.241 | <0.001 * | 0.249 | <0.001 * |
| Lower radii | 0.030 | 0.077 | 0.030 | 0.081 | 0.026 | 0.104 |
| Web height from ground (cm) | 0.002 | 0.620 | 0.010 | 0.319 | 0.007 | 0.388 |
| CTL (cm) | 0.096 | 0.001 * | 0.061 | 0.012 | 0.084 | 0.003 |
| Upper spirals | 0.035 | 0.059 | 0.045 | 0.031 | 0.019 | 0.158 |
| No. of prey | 0.038 | 0.048 | 0.036 | 0.055 | 0.039 | 0.046 |
| Asymmetry | 0.029 | 0.083 | 0.034 | 0.062 | 0.026 | 0.103 |
| Lower spirals | 0.003 | 0.562 | 0.007 | 0.404 | 0.001 | 0.784 |
| Plant height (cm) | 0.000 | 0.931 | 0.011 | 0.287 | 0.011 | 0.294 |
| Web size (cm) | 0.345 | <0.001 * | 0.247 | <0.001 * | 0.268 | <0.001 * |
| Vertical web diameter (cm) | 0.382 | <0.001 * | 0.295 | <0.001 * | 0.288 | <0.001 * |
| Foliage radius (cm) | 0.001 | 0.792 | 0.024 | 0.120 | 0.011 | 0.281 |
| Web Characteristic | R2 | p | n |
|---|---|---|---|
| Horizontal web diameter (cm) | 0.006 | 0.181 | 105 |
| Mesh size (mm) | 0.025 | 0.632 | 105 |
| Upper radii | 0.021 | 0.268 | 105 |
| Anchor points | 0.066 | 0.001 * | 105 |
| Capture area (cm2) | 0.027 | 0.165 | 105 |
| Lower radii | 0.006 | 0.865 | 105 |
| Web height from ground (cm) | 0.041 | 0.004 * | 105 |
| CTL (cm) | 0.005 | 0.331 | 105 |
| Upper spirals | 0.000 | 0.771 | 105 |
| No. of prey | 0.093 | 0.020 * | 94 |
| Asymmetry | 0.001 | 0.303 | 105 |
| Lower spirals | 0.001 | 0.520 | 105 |
| Plant height (cm) | 0.058 | <0.001 * | 105 |
| Web size (cm) | 0.028 | 0.149 | 105 |
| Vertical web diameter (cm) | 0.040 | 0.254 | 105 |
| Foliage radius (cm) | 0.060 | 0.001 * | 105 |
| Web Anomaly | Fourth Leg Length (mm) | Carapace Width (mm) | Body Length (mm) | n | |||
|---|---|---|---|---|---|---|---|
| R2 | p | R2 | P | R2 | p | ||
| Two spirals stuck together | 0.001 | 0.793 | 0.012 | 0.269 | 0.007 | 0.382 | 104 |
| Supernumerary | 0.003 | 0.670 | 0.003 | 0.696 | 0.001 | 0.788 | 55 |
| More than two spirals stuck together | 0.057 | 0.014 | 0.028 | 0.091 | 0.031 | 0.076 | 104 |
| Deviated | 0.185 | 0.001 * | 0.011 | 0.136 | 0.184 | 0.001 * | 94 |
| Discontinuity | 0.020 | 0.159 | 0.001 | 0.794 | 0.010 | 0.316 | 99 |
| Y-shaped | 0.023 | 0.190 | 0.018 | 0.244 | 0.016 | 0.263 | 78 |
| Non-parallel | 0.000 | 0.906 | 0.007 | 0.493 | 0.001 | 0.761 | 69 |
| Stop and return | 0.000 | 0.936 | 0.000 | 0.907 | 0.000 | 0.971 | 104 |
| Zigzag | 0.000 | 0.994 | 0.003 | 0.632 | 0.007 | 0.443 | 90 |
| Hole | 0.015 | 0.209 | 0.001 | 0.819 | 0.003 | 0.570 | 104 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bilal, A.; Butt, A.; Kazam, A.; Ali, S.; Isha; Chang, Y.-C. Investigating the Influence of Anthropogenic Activities on Behavioral Changes of an Orb Web Spider (Neoscona vigilans). Insects 2024, 15, 609. https://doi.org/10.3390/insects15080609
Bilal A, Butt A, Kazam A, Ali S, Isha, Chang Y-C. Investigating the Influence of Anthropogenic Activities on Behavioral Changes of an Orb Web Spider (Neoscona vigilans). Insects. 2024; 15(8):609. https://doi.org/10.3390/insects15080609
Chicago/Turabian StyleBilal, Ahmad, Abida Butt, Adeel Kazam, Shakir Ali, Isha, and Young-Cheol Chang. 2024. "Investigating the Influence of Anthropogenic Activities on Behavioral Changes of an Orb Web Spider (Neoscona vigilans)" Insects 15, no. 8: 609. https://doi.org/10.3390/insects15080609
APA StyleBilal, A., Butt, A., Kazam, A., Ali, S., Isha, & Chang, Y.-C. (2024). Investigating the Influence of Anthropogenic Activities on Behavioral Changes of an Orb Web Spider (Neoscona vigilans). Insects, 15(8), 609. https://doi.org/10.3390/insects15080609