
Meta-Analysis and MaxEnt Model Prediction of the Distribution of Phenacoccus solenopsis Tinsley in China under the Context of Climate Change


Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Meta-Analysis
2.1.1. Data Collection and Extraction
2.1.2. Data Analysis
2.2. MaxEnt Prediction and Analysis
2.2.1. Species Occurrence Data
2.2.2. Environmental Variables
2.2.3. Optimization of Model Parameters
2.2.4. Shift of Suitable Habitat Distribution Center
2.2.5. Model Evaluation
3. Results
3.1. Survival Response to Temperature
3.1.1. Cumulative Effect Size
3.1.2. Introduction of Explanatory Variables
3.1.3. Model Diagnostics
3.2. Predicting the Distribution of P. solenopsis in China
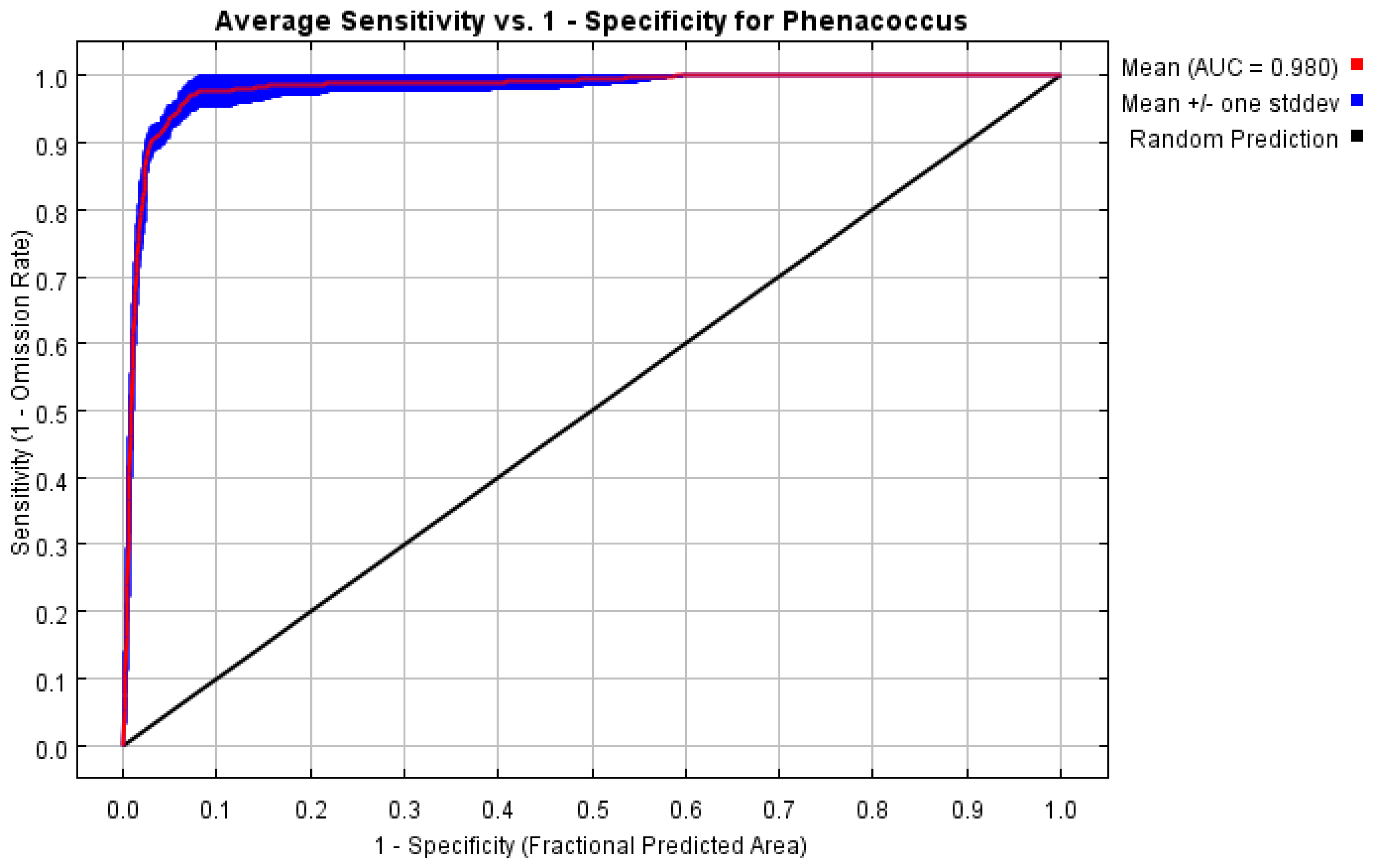
3.2.1. Prediction Accuracy Evaluation of MaxEnt Model
3.2.2. Main Environmental Factors Affecting Distribution
3.2.3. Current Potential Distribution
3.2.4. Potentially Suitable Climatic Distributions in the Future
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fuchs, T.W.; Stewart, J.W.; Minzenmayer, R.; Rose, M. First record of Phenacoccus solenopsis Tinsley in cultivated cotton in the United States. Southwest. Entomol. 1991, 16, 215–221. [Google Scholar]
- Tong, H.-J.; Ao, Y.; Li, Z.-H.; Wang, Y.; Jiang, M.-X. Invasion biology of the cotton mealybug, Phenacoccus solenopsis Tinsley: Current knowledge and future directions. J. Integr. Agric. 2019, 18, 758–770. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, T.; Tian, H.; Wan, F.; Zhang, G. Range expansion of the invasive cotton mealybug, Phenacoccus solenopsis Tinsley: An increasing threat to agricultural and horticultural crops in China. J. Integr. Agric. 2020, 19, 881–885. [Google Scholar] [CrossRef]
- Spodek, M.; Ben-Dov, Y.; Mondaca, L.; Protasov, A.; Erel, E.; Mendel, Z. The cotton mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Israel: Pest status, host plants and natural enemies. Phytoparasitica 2018, 46, 45–55. [Google Scholar] [CrossRef]
- Hodgson, C.; Abbas, G.; Arif, M.J.; Saeed, S.; Karar, H. Phenacoccus solenopsis Tinsley (Sternorrhyncha: Coccoidea: Pseudococcidae), an invasive mealybug damaging cotton in Pakistan and India, with a discussion on seasonal morphological variation. Zootaxa 2008, 1913, 1–35. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134–141. [Google Scholar] [CrossRef]
- Deng, X.; Xu, D.; Liao, W.; Wang, R.; Zhuo, Z. Predicting the distributions of Scleroderma guani (Hymenoptera: Bethylidae) under climate change in China. Ecol. Evol. 2022, 12, e9410. [Google Scholar] [CrossRef]
- Wei, X.; Xu, D.; Liu, Q.; Wu, Y.; Zhuo, Z. Predicting the potential distribution range of Batocera horsfieldi under CMIP6 climate change using the MaxEnt model. J. Econ. Entomol. 2023, 117, 187–198. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of MaxEnt. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. S2), 19729–19736. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with MaxEnt: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Xu, D.; Zhuo, Z.; Wang, R.; Ye, M.; Pu, B. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling. Glob. Ecol. Conserv. 2019, 19, e691. [Google Scholar] [CrossRef]
- Mamun, M.; Kim, S.; An, K. Distribution pattern prediction of an invasive alien species largemouth bass using a maximum entropy model (MaxEnt) in the Korean peninsula. J. Asia-Pac. Biodivers. 2018, 11, 516–524. [Google Scholar] [CrossRef]
- Kramer Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Merow, C.; Silander, J.A. A comparison of Maxlike and MaxEnt for modelling species distributions. Methods Ecol. Evol. 2014, 5, 215–225. [Google Scholar] [CrossRef]
- Hou, Z.; Sun, Z.; Du, G.; Shao, D.; Zhong, Q.; Yang, S. Assessment of suitable cultivation region for Pepino (Solanum muricatum) under different climatic conditions using the MaxEnt model and adaptability in the Qinghai–Tibet plateau. Heliyon 2023, 9, e18974. [Google Scholar] [CrossRef]
- Zheng, Y.; Yuan, C.; Matsushita, N.; Lian, C.; Geng, Q. Analysis of the distribution pattern of the ectomycorrhizal fungus Cenococcum geophilum under climate change using the optimized MaxEnt model. Ecol. Evol. 2023, 13, e10565. [Google Scholar] [CrossRef]
- Zhao, G.; Cui, X.; Sun, J.; Li, T.; Wang, Q.; Ye, X.; Fan, B. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, L.; Zhou, X.; Chen, R.; Zhao, G.; Zhang, F. Prediction of the potentially suitable areas of Leonurus japonicus in China based on future climate change using the optimized MaxEnt model. Ecol. Evol. 2023, 13, e10597. [Google Scholar] [CrossRef]
- Wiese, D.; Escalante, A.A.; Murphy, H.; Henry, K.A.; Gutierrez-Velez, V.H. Integrating environmental and neighborhood factors in MaxEnt modeling to predict species distributions: A case study of Aedes albopictus in southeastern Pennsylvania. PLoS ONE 2019, 14, e223821. [Google Scholar] [CrossRef]
- Wan, J.; Qi, G.-J.; Ma, J.; Ren, Y.; Wang, R.; McKirdy, S. Predicting the potential geographic distribution of Bactrocera bryoniae and Bactrocera neohumeralis (Diptera: Tephritidae) in China using MaxEnt ecological niche modeling. J. Integr. Agric. 2020, 19, 2072–2082. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Zhu, G.; Qiao, H. Effect of the MaxEnt model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using MaxEnt. PeerJ 2019, 7, 6281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.P.; Yuan, X.J.; Fan, J.Y.; Wang, M.L. Effects of model parameters in MaxEnt modeling of ecological niche and geographic distribution: Case study of the brown marmorated stink bug, Halyomorpha haly. J. Biosaf. 2018, 27, 46–51. [Google Scholar]
- Zhao, Z.; Xiao, N.; Shen, M.; Li, J. Comparison between optimized MaxEnt and random forest modeling in predicting potential distribution: A case study with Quasipaa boulengeri in China. Sci. Total Environ. 2022, 842, 156867. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better MaxEnt models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Muscarella, R.; Galante, P.J.; Soley Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENM eval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for MaxEnt ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Sharma, R.; Khan, S.; Kaul, V. Predicting the potential habitat suitability and distribution of “Weed-Onion” (Asphodelus tenuifolius Cavan.) in India under predicted climate change scenarios. J. Agric. Food Res. 2023, 14, 100697. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Osorio-Olvera, L.; Jiménez-García, D. An exhaustive analysis of heuristic methods for variable selection in ecological niche modeling and species distribution modeling. Ecol. Inform. 2019, 53, 100983. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. MaxEnt modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Peng, D.; Li, Y.; Chen, Z.; Zhai, Y.; Chen, L.; Bo, H. Potential global distribution of the guava root-knot nematode Meloidogyne enterolobii under different climate change scenarios using MaxEnt ecological niche modeling. J. Integr. Agric. 2023, 22, 2138–2150. [Google Scholar]
- Araujo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Change Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef]
- Kumar, R.; Nagrare, V.S.; Nitharwal, M.; Swami, D.; Prasad, Y.G. Within-plant distribution of an invasive mealybug, Phenacoccus solenopsis, and associated losses in cotton. Phytoparasitica 2014, 42, 311–316. [Google Scholar] [CrossRef]
- Marchioro, C.A.; Krechemer, F.S. Potential global distribution of Diabrotica species and the risks for agricultural production. Pest Manag. Sci. 2018, 74, 2100–2109. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Ramasamy, M.; Das, B.; Ramesh, R. Predicting climate change impacts on potential worldwide distribution of fall armyworm based on CMIP6 projections. J. Pest Sci. 2022, 95, 841–854. [Google Scholar] [CrossRef]
- Rhoden, C.M.; Peterman, W.E.; Taylor, C.A. MaxEnt-directed field surveys identify new populations of narrowly endemic habitat specialists. PeerJ 2017, 5, e3632. [Google Scholar] [CrossRef]
- Valencia-Rodríguez, D.; Jiménez-Segura, L.; Rogéliz, C.A.; Parra, J.L. Ecological niche modeling as an effective tool to predict the distribution of freshwater organisms: The case of the Sabaleta Brycon henni (Eigenmann, 1913). PLoS ONE 2021, 16, e247876. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, H.; Zhao, W.; Zhao, Q. Niche shifts and the potential distribution of Phenacoccus solenopsis (Hemiptera: Pseudococcidae) under climate change. PLoS ONE 2017, 12, e180913. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ge, X.; Chen, L.; Zhang, L.; Wang, T.; Zong, S. Climate change impacts on the potential distribution of Eogystia hippophaecolus in China. Pest Manag. Sci. 2019, 75, 215–223. [Google Scholar] [CrossRef]
- Wang, F.; Wang, D.; Guo, G.; Zhang, M.; Lang, J.; Wei, J. Potential distributions of the invasive barnacle scale Ceroplastes cirripediformis (Hemiptera: Coccidae) under climate change and implications for its management. J. Econ. Entomol. 2021, 114, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Biber-Freudenberger, L.; Ziemacki, J.; Tonnang, H.E.; Borgemeister, C. Future risks of pest species under changing climatic conditions. PLoS ONE 2016, 11, e153237. [Google Scholar] [CrossRef]
- Wei, J.; Peng, L.; He, Z.; Lu, Y.; Wang, F. Potential distribution of two invasive pineapple pests under climate change. Pest Manag. Sci. 2020, 76, 1652–1663. [Google Scholar] [CrossRef]
- Kou, J.; Wang, T.; Yu, F.; Sun, Y.; Feng, C.; Shao, X. The moss genus Didymodon as an indicator of climate change on the Tibetan Plateau. Ecol. Indic. 2020, 113, 106204. [Google Scholar] [CrossRef]
- Harrington, R.; Clark, S.J.; Welham, S.J.; Verrier, P.J.; Denholm, C.H.; Hulle, M.; Maurice, D.; Rounsevell, M.D.; Cocu, N.; Consortium, E.U.E. Environmental change and the phenology of European aphids. Glob. Chang. Biol. 2007, 13, 1550–1564. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Emission | Description |
|---|---|
| SSP1–2.6 | SSP1 (Low forcing scenario) upgrade to RCP2.6 scenario (Radiative forcing reaches 2.6 W/m2 in 2100) |
| SSP3–7.0 | SSP3 (Medium forcing scenario) new RCP7.0 emission path (Radiative forcing reaches 7.0 W/m2 in 2100) |
| SSP5–8.5 | SSP5 (High forcing scenario) upgrade to RCP8.5 scenario (SSP5 is the only SSP scenario that can achieve radiative forcing of 8.5 W/m2 in 2100) |
| Variable | Description | Whether to Retain after Filtering |
|---|---|---|
| Bio01 | Annual Mean Temperature (°C) | No |
| Bio02 | Mean Diurnal Range (Mean of Monthly (Max Temp–Min Temp)) (°C) | No |
| Bio03 | Isothermality (BIO2/BIO7) (×100) | Yes |
| Bio04 | Temperature Seasonality (Standard Deviation × 100) | No |
| Bio05 | Max Temperature of Warmest Month (°C) | No |
| Bio06 | Min Temperature of Coldest Month (°C) | No |
| Bio07 | Temperature Annual Range (BIO5–BIO6) (°C) | No |
| Bio08 | Mean Temperature of Wettest Quarter (°C) | Yes |
| Bio09 | Mean Temperature of Driest Quarter (°C) | Yes |
| Bio10 | Mean Temperature of Warmest Quarter (°C) | No |
| Bio11 | Mean Temperature of Coldest Quarter (°C) | No |
| Bio12 | Annual Precipitation (mm) | No |
| Bio13 | Precipitation of Wettest Month (mm) | Yes |
| Bio14 | Precipitation of Driest Month (mm) | Yes |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | Yes |
| Bio16 | Precipitation of Wettest Quarter (mm) | No |
| Bio17 | Precipitation of Driest Quarter (mm) | No |
| Bio18 | Precipitation of Warmest Quarter (mm) | Yes |
| Bio19 | Precipitation of Coldest Quarter (mm) | No |
| Altitude | Altitude (m) | Yes |
| Slope | Incline (°) | Yes |
| Variable | Percent Contribution (%) | Permutation Importance (%) |
|---|---|---|
| Bio18 | 40.6 | 5.3 |
| Bio8 | 21 | 47.5 |
| Bio3 | 17.9 | 29.5 |
| Bio13 | 5.5 | 1.6 |
| Altitude | 4.5 | 2.3 |
| Bio14 | 3.7 | 0.4 |
| Bio9 | 3.6 | 11.9 |
| Bio15 | 2.8 | 1.4 |
| Slope | 0.3 | 0.1 |
| Period | Highly Suitable | Moderately Suitable | Low Suitable | Total Suitable |
|---|---|---|---|---|
| Current | 16.77 | 158.41 | 250.49 | 425.67 |
| 2050s, SSPs1–2.6 | 23.33 (+39.07%) | 187.55 (+18.40%) | 288.47 (+15.16%) | 499.35 (+17.31%) |
| 2050s, SSPs3–7.0 | 23.57 (+40.52%) | 167.27 (+5.60%) | 300.39 (+19.92%) | 491.23 (+15.40%) |
| 2050s, SSPs5–8.5 | 23.35 (+39.18%) | 184.21 (+16.29%) | 370.58 (+47.94%) | 578.13 (+35.82%) |
| 2070s, SSPs1–2.6 | 21.57 (+28.59%) | 166.00 (+4.79%) | 292.06 (+16.59%) | 479.63 (+12.67%) |
| 2070s, SSPs3–7.0 | 16.32 (−2.68%) | 188.05 (+18.71%) | 387.32 (+54.63%) | 591.70 (+39.00%) |
| 2070s, SSPs5–8.5 | 29.26 (+74.40%) | 181.63 (+14.66%) | 328.49 (+31.14%) | 539.37 (+26.27%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Peng, Y.; Xu, D.; Zhuo, Z. Meta-Analysis and MaxEnt Model Prediction of the Distribution of Phenacoccus solenopsis Tinsley in China under the Context of Climate Change. Insects 2024, 15, 675. https://doi.org/10.3390/insects15090675
Liu Z, Peng Y, Xu D, Zhuo Z. Meta-Analysis and MaxEnt Model Prediction of the Distribution of Phenacoccus solenopsis Tinsley in China under the Context of Climate Change. Insects. 2024; 15(9):675. https://doi.org/10.3390/insects15090675
Chicago/Turabian StyleLiu, Zhiqian, Yaqin Peng, Danping Xu, and Zhihang Zhuo. 2024. "Meta-Analysis and MaxEnt Model Prediction of the Distribution of Phenacoccus solenopsis Tinsley in China under the Context of Climate Change" Insects 15, no. 9: 675. https://doi.org/10.3390/insects15090675
APA StyleLiu, Z., Peng, Y., Xu, D., & Zhuo, Z. (2024). Meta-Analysis and MaxEnt Model Prediction of the Distribution of Phenacoccus solenopsis Tinsley in China under the Context of Climate Change. Insects, 15(9), 675. https://doi.org/10.3390/insects15090675