


Temperature Variation Regulates the Trade-Off Between Pre- and Post-Hatching Investment in a Burying Beetle


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Beetle Husbandry
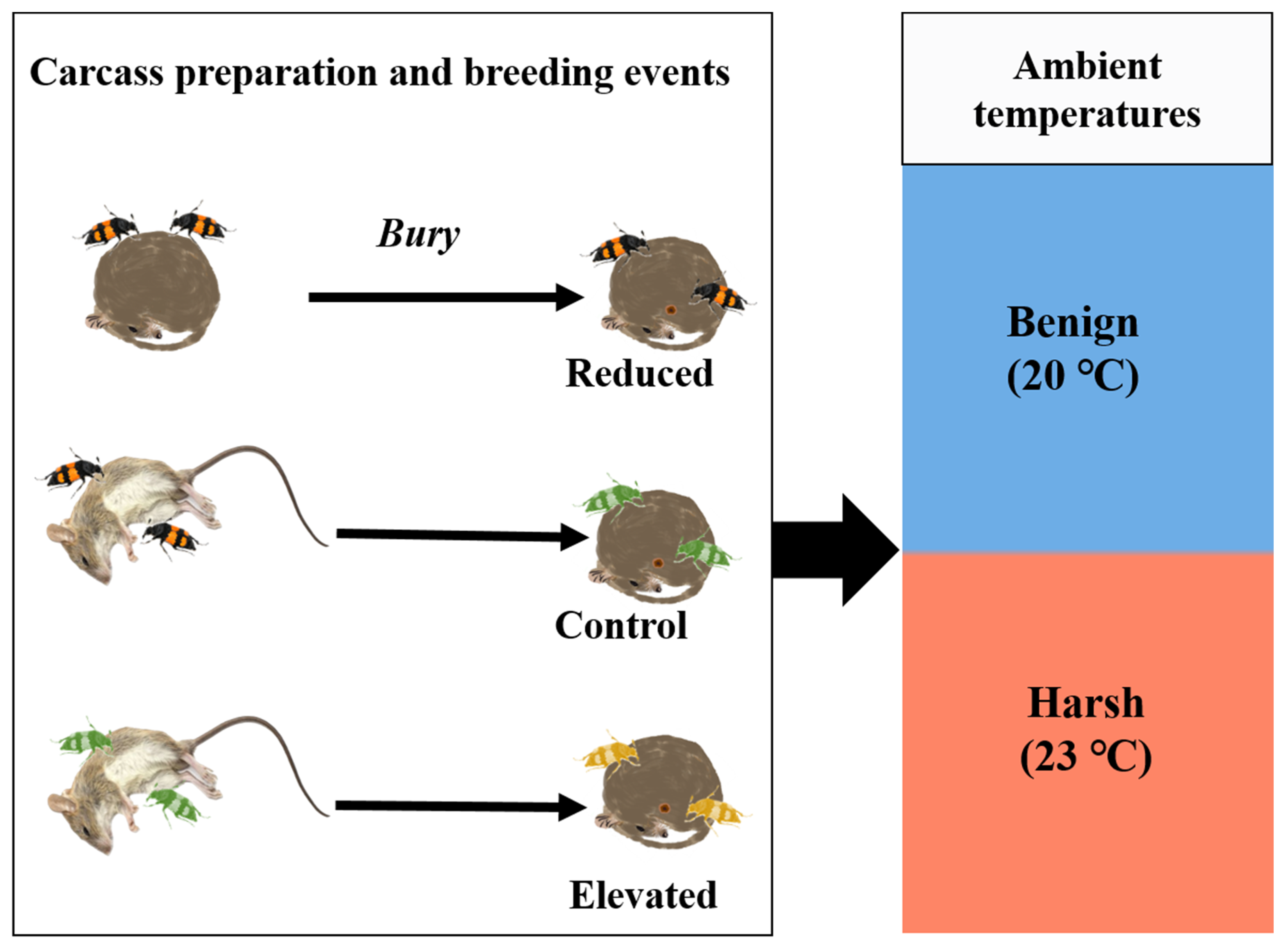
2.2. Experimental Protocol
2.3. Statistical Analyses
3. Results
3.1. Effects of Carcass-Preparation Investment and Ambient Temperature on Parental Care
3.1.1. Male Pre- and Post-Hatching Care
3.1.2. Female Pre- and Post-Hatching Care
3.2. Effects of Carcass-Preparation Investment and Ambient Temperature on Reproductive Success and Parental Body Mass Change
3.2.1. Reproductive Success
3.2.2. Offspring Performance
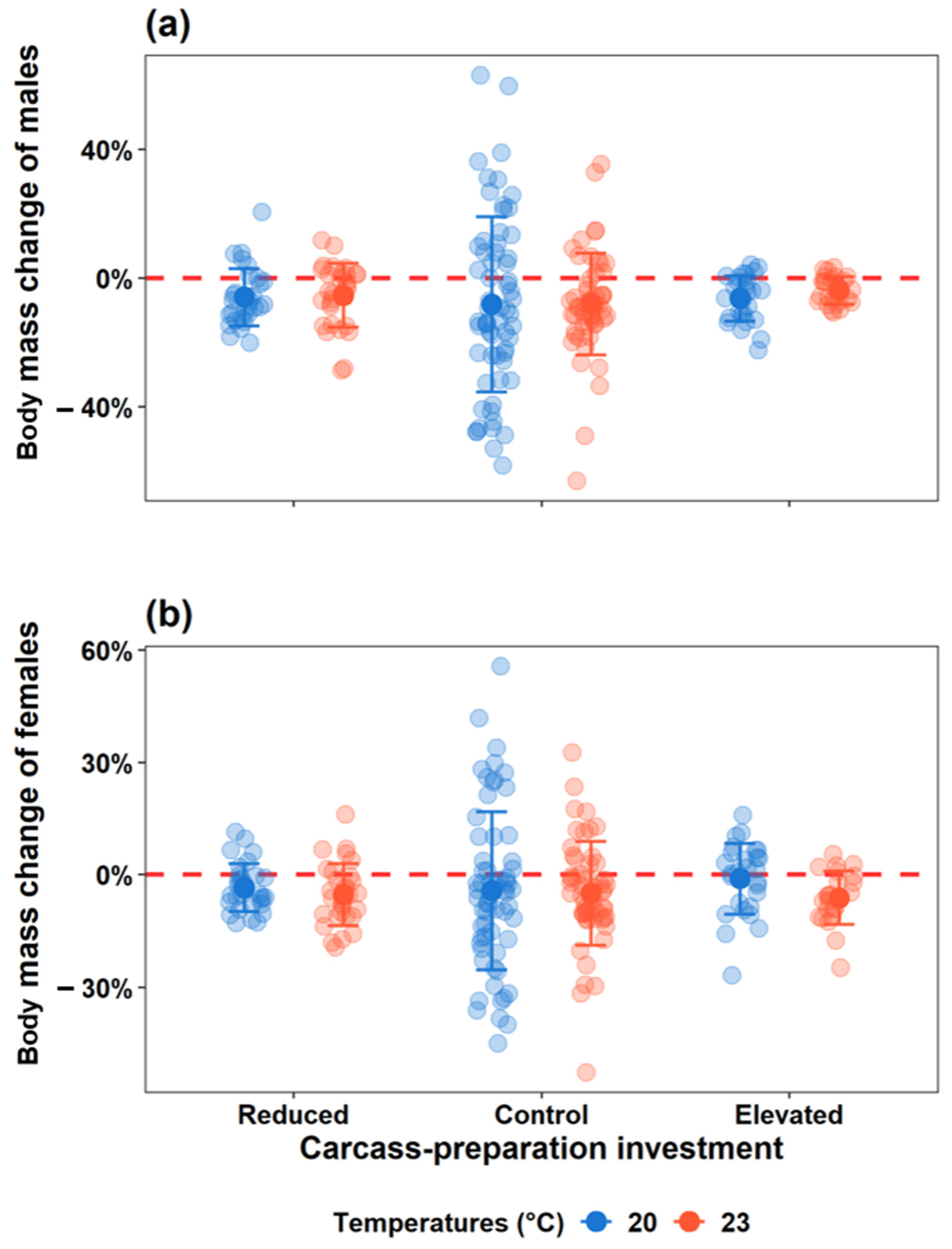
3.2.3. Parental Body Mass Change
4. Discussion
4.1. Reduced Carcass-Preparation Investment and Ambient Temperature Impact Parental Care and Reproductive Success
4.2. Elevated Carcass-Preparation Investment and Ambient Temperature Impact Parental Care and Reproductive Success
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2023: Synthesis Report; Contribution of Working Groups i, ii and iii to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Saunders, D.A.; Mawson, P.; Dawson, R. The impact of two extreme weather events and other causes of death on Carnaby’s black cockatoo: A promise of things to come for a threatened species? Pac. Conserv. Biol. 2011, 17, 141–148. [Google Scholar]
- Fragueira, R.; Helfenstein, F.; Fischer, K.; Beaulieu, M. Birds of different morphs use slightly different strategies to achieve similar reproductive performance following heatwave exposure. J. Anim. Ecol. 2021, 90, 2594–2608. [Google Scholar] [CrossRef] [PubMed]
- Hunt, J.; Simmons, L.W. Behavioural dynamics of biparental care in the dung beetle Onthophagus taurus. Anim. Behav. 2002, 64, 65–75. [Google Scholar]
- Nguyen, T.M.; Bressac, C.; Chevrier, C. Heat stress affects male reproduction in a parasitoid wasp. J. Insect Physiol. 2013, 59, 248–254. [Google Scholar]
- Pilakouta, N.; Allan, D.; Moore, E.; Russell, A.A. Chronic and acute thermal stressors have non-additive effects on fertility. Proc. R. Soc. B Biol. Sci. 2024, 291, rspb20241086. [Google Scholar]
- Sales, K.; Vasudeva, R.; Dickinson, M.E.; Godwin, J.L.; Lumley, A.J.; Michalczyk, Ł.; Hebberecht, L.; Thomas, P.; Franco, A.; Gage, M.J.G. Experimental heatwaves compromise sperm function and cause transgenerational damage in a model insect. Nat. Commun. 2018, 9, 4771. [Google Scholar]
- Chevrier, C.; Nguyen, T.M.; Bressac, C. Heat shock sensitivity of adult male fertility in the parasitoid wasp Anisopteromalus calandrae (Hymenoptera, Pteromalidae). J. Therm. Biol. 2019, 85, 102419. [Google Scholar]
- Porcelli, D.; Gaston, K.J.; Butlin, R.K.; Snook, R.R. Local adaptation of reproductive performance during thermal stress. J. Evol. Biol. 2017, 30, 422–429. [Google Scholar]
- Martinet, B.; Zambra, E.; Przybyla, K.; Lecocq, T.; Anselmo, A.; Nonclercq, D.; Rasmont, P.; Michez, D.; Hennebert, E. Mating under climate change: Impact of simulated heatwaves on the reproduction of model pollinators. Funct. Ecol. 2021, 35, 739–752. [Google Scholar]
- Rukke, B.A.; Sivasubramaniam, R.; Birkemoe, T.; Aak, A. Temperature stress deteriorates bed bug (Cimex lectularius) populations through decreased survival, fecundity and offspring success. PLoS ONE 2018, 13, e0193788. [Google Scholar]
- McCowan, L.S.C.; Griffith, S.C. Baked Eggs: Catastrophic heatwave-induced reproductive failure in the desert-adapted Zebra finch (Taeniopygia guttata). Int. J. Avian Sci. 2021, 163, 1207–1216. [Google Scholar] [CrossRef]
- Pilakouta, N.; Sellers, L.; Barratt, R.; Ligonniere, A. The consequences of heatwaves for animal reproduction are timing-dependent. Funct. Ecol. 2023, 37, 2425–2433. [Google Scholar] [CrossRef]
- Wiley, E.M.; Ridley, A.R. The effects of temperature on offspring provisioning in a cooperative breeder. Anim. Behav. 2016, 117, 187–195. [Google Scholar] [CrossRef]
- Moss, J.B.; Moore, A.J. Constrained flexibility of parental cooperation limits adaptive responses to harsh conditions. Evolution 2021, 75, 1835–1849. [Google Scholar] [CrossRef]
- Kumar, S.; Kler, T.K.; Sekhon, G.S.; Sahni, T. Impacts on avian migratory patterns due to climate change and hormonal disruption: A review. Mitig. Adapt. Strateg. Glob. Ch. 2024, 29, 69. [Google Scholar] [CrossRef]
- Both, C.; Bouwhuis, S.; Lessells, C.M.; Visser, M.E. Climate change and population declines in a long-distance migratory bird. Nature 2006, 441, 81–83. [Google Scholar] [CrossRef]
- Charmantier, A.; McCleery, R.H.; Cole, L.R.; Perrins, C.; Kruuk, L.E.B.; Sheldon, B.C. Adaptive phenotypic plasticity in response to climate change in a wild bird population. Science 2008, 320, 800–803. [Google Scholar] [CrossRef]
- Dawson, R.D.; Lawrie, C.C.; O’Brien, E.L. The importance of microclimate variation in determining size, growth and survival of avian offspring: Experimental evidence from a cavity nesting passerine. Oecologia 2005, 144, 499–507. [Google Scholar] [CrossRef]
- McKechnie, A.E. Physiological and morphological effects of climate change. In Effects of Climate Change on Birds, 2nd ed.; Dunn, P.O., Møller, A.P., Eds.; Oxford University Press: Oxford, UK, 2019; pp. 133–201. [Google Scholar]
- Sun, B.-J.; Wang, Y.; Wang, Y.; Lu, H.-L.; Du, W.-G. Anticipatory parental effects in a subtropical lizard in response to experimental warming. Front. Zool. 2018, 15, 51. [Google Scholar] [CrossRef]
- Sidhu, K.K.; Zafeiri, S.; Malcolm, C.; Caplat, P.; Lancaster, L.T.; Bocedi, G.; Pilakouta, N. Heatwaves during early development have long-term consequences for parental care in adulthood. Anim. Behav. 2024, 217, 65–72. [Google Scholar] [CrossRef]
- Walsh, B.S.; Parratt, S.R.; Hoffman, A.A.; Atkinson, D.; Snook, R.R.; Bretman, A.; Price, T.A.R. The impact of climate change on fertility. Trends Ecol. Evol. 2019, 34, 249–259. [Google Scholar]
- Stearns, S.C. Trade-offs in life-history evolution. Funct. Ecol. 1989, 3, 259–268. [Google Scholar] [CrossRef]
- Gonzalez, O.; Zedrosser, A.; Pelletier, F.; Swenson, J.E.; Festa-Bianchet, M. Litter reductions reveal a trade-off between offspring size and number in Brown bears. Behav. Ecol. Sociobiol. 2012, 66, 1025–1032. [Google Scholar]
- Richardson, J.; Stephens, J.; Smiseth, P.T. Increased allocation to reproduction reduces future competitive ability in a burying beetle. J. Anim. Ecol. 2020, 89, 1918–1926. [Google Scholar] [CrossRef]
- Santos, E.S.A.; Nakagawa, S. The costs of parental care: A meta-analysis of the trade-off between parental effort and survival in birds. J. Evol. Biol. 2012, 25, 1911–1917. [Google Scholar] [CrossRef]
- Trivers, R. Parental investment and sexual selection. In Sexual Selection and the Descent of Man; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 1871–1971. [Google Scholar]
- Bonnet, X.; Lourdais, O.; Shine, R.; Naulleau, G. Reproduction in a typical capital breeder: Costs, currencies, and complications in the Aspic viper. Ecology 2002, 83, 2124–2135. [Google Scholar] [CrossRef]
- van Noordwijk, A.J.; de Jong, G. Acquisition and allocation of resources: Their influence on variation in life history tactics. Am. Nat. 1986, 128, 137–142. [Google Scholar]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009; pp. 88–125. [Google Scholar]
- Scott, M.P. The ecology and behavior of burying beetles. Annu. Rev. Entomol. 1998, 43, 595–618. [Google Scholar]
- Bartlett, J. The Behavioural Ecology of the Burying Beetle. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 1987. [Google Scholar]
- Arce, A.N.; Johnston, P.R.; Smiseth, P.T.; Rozen, D.E. Mechanisms and fitness effects of antibacterial defences in a carrion beetle. J. Evol. Biol. 2012, 25, 930–937. [Google Scholar]
- Müller, J.K.; Eggert, A.-K.; Sakaluk, S.K. Carcass maintenance and biparental brood care in burying beetles: Are males redundant? Ecol. Entomol. 1998, 23, 195–200. [Google Scholar]
- Eggert, A.-K.; Reinking, M.; Müller, J.K. Parental care improves offspring survival and growth in burying beetles. Anim. Behav. 1998, 55, 97–107. [Google Scholar] [PubMed]
- Schrader, M.; Hughes, P.; Jenkins, S.; Kusher, I.; Lopez, J.; Oglesby, H.; McGhee, K.E. Can age-related changes in parental care modulate inbreeding depression? A test using the burying beetle, Nicrophorus orbicollis. Ecol. Evol. 2022, 12, e9391. [Google Scholar] [PubMed]
- Pilakouta, N.; Jamieson, S.; Moorad, J.A.; Smiseth, P.T. Parental care buffers against inbreeding depression in burying beetles. Proc. Natl. Acad. Sci. USA 2015, 112, 8031–8035. [Google Scholar] [PubMed]
- Bartlett, J.; Ashworth, C.M. Brood size and fitness in Nicrophorus vespilloides (Coleoptera: Silphidae). Behav. Ecol. Sociobiol. 1988, 22, 429–434. [Google Scholar]
- Keppner, E.M.; Laubenthal, M.; Prang, M.A.; Conrad, T.; Steiger, S. Harsh nutritional environment has positive and negative consequences for family living in a burying beetle. Ecol. Evol. 2023, 13, e9699. [Google Scholar]
- Richardson, J.; Smiseth, P.T. Nutrition during sexual maturation and at the time of mating affects mating behaviour in both sexes of a burying beetle. Anim. Behav. 2019, 151, 77–85. [Google Scholar]
- Richardson, J.; Smiseth, P.T. Effects of variation in resource acquisition during different stages of the life cycle on life-history traits and trade-offs in a burying beetle. J. Evol. Biol. 2019, 32, 19–30. [Google Scholar]
- Eggert, A.-K.; Müller, J.K. Timing of oviposition and reproductive skew in cobreeding female burying beetles (Nicrophorus vespilloides). Behav. Ecol. 2000, 11, 357–366. [Google Scholar]
- Komdeur, J.; Schrama, M.J.J.; Meijer, K.; Moore, A.J.; Beukeboom, L.W. Cobreeding in the burying beetle, Nicrophorus vespilloides: Tolerance rather than cooperation. Ethology 2013, 119, 1138–1148. [Google Scholar]
- Ma, L.; Versteegh, M.A.; Hammers, M.; Komdeur, J. Sex-specific influence of communal breeding experience on parenting performance and fitness in a burying beetle. R. Soc. Open Sci. 2022, 9, 211179. [Google Scholar]
- Richardson, J.; Smiseth, P.T. Maternity uncertainty in cobreeding beetles: Females lay more and larger eggs and provide less care. Behav. Ecol. 2020, 31, 641–650. [Google Scholar]
- Smiseth, P.T.; Dawson, C.; Varley, E.; Moore, A.J. How do caring parents respond to mate loss? differential response by males and females. Anim. Behav. 2005, 69, 551–559. [Google Scholar]
- Wang, W.; Ma, L.; Versteegh, M.A.; Wu, H.; Komdeur, J. Parental care system and brood size drive sex difference in reproductive allocation: An experimental study on burying beetles. Front. Ecol. Evol. 2021, 9, 739396. [Google Scholar]
- Grew, R.; Ratz, T.; Richardson, J.; Smiseth, P. Parental care buffers against effects of ambient temperature on offspring performance in an insect. Behav. Ecol. 2019, 30, 1443–1450. [Google Scholar]
- De Gasperin, O.; Kilner, R.M. Interspecific interactions change the outcome of sexual conflict over prehatching parental investment in the burying beetle Nicrophorus vespilloides. Ecol. Evol. 2015, 5, 5552–5560. [Google Scholar]
- De Gasperin, O.; Duarte, A.; Troscianko, J.; Kilner, R.M. Fitness costs associated with building and maintaining the burying beetle’s carrion nest. Sci. Rep. 2016, 6, 35293. [Google Scholar]
- Carter, D.O.; Yellowlees, D.; Tibbett, M. Temperature affects microbial decomposition of cadavers (Rattus rattus) in contrasting soils. Appl. Soil Ecol. 2008, 40, 129–137. [Google Scholar]
- Mattey, S.N.; Strutt, L.; Smiseth, P.T. Intergenerational effects of inbreeding in Nicrophorus vespilloides: Offspring suffer fitness costs when either they or their parents are inbred. J. Evol. Biol. 2013, 26, 843–853. [Google Scholar]
- Wang, W.; Ma, L.; Versteegh, M.A.; Wu, H.; Komdeur, J. Detection of reproductive trade-offs is influenced by resource availability and maintenance: An experimental study in the burying beetle (Nicrophorus vespilloides). Behav. Ecol. Sociobiol. 2022, 76, 76. [Google Scholar]
- Bladon, E.K.; English, S.; Pascoal, S.; Kilner, R.M. Early-life effects on body size in each sex interact to determine reproductive success in the burying beetle Nicrophorus vespilloides. J. Evol. Biol. 2020, 33, 1725–1734. [Google Scholar]
- Jarrett, B.J.M.; Schrader, M.; Rebar, D.; Houslay, T.M.; Kilner, R.M. Cooperative interactions within the family enhance the capacity for evolutionary change in body size. Nat. Ecol. Evol. 2017, 1, 0178. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-Plus, 3rd ed.; Springer: New York, NY, USA, 2001. [Google Scholar]
- Hartig, F. R Package DHARMa, version 0.4.6; Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Fox, J.; Weisberg, S.; Price, B. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Lenth, R.V. R Package Emmeans, version 1.7.1.; Estimated Marginal Means, Aka Least-Squares Means; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Briscoe, N.J.; Morris, S.D.; Mathewson, P.D.; Buckley, L.B.; Jusup, M.; Levy, O.; Maclean, I.M.D.; Pincebourde, S.; Riddell, E.A.; Roberts, J.A.; et al. Mechanistic forecasts of species responses to climate change: The promise of biophysical ecology. Glob. Change Biol. 2023, 29, 1451–1470. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, J.; Kokko, H.; Parker, G.A. What do isogamous organisms teach us about sex and the two sexes? Philos. Trans. R. Soc. B 2016, 371, 20150532. [Google Scholar]
- Cotter, S.C.; Kilner, R.M. Sexual division of antibacterial resource defence in breeding burying beetles, Nicrophorus vespilloides. J. Anim. Ecol. 2010, 79, 35–43. [Google Scholar] [CrossRef]
- Abram, P.K.; Boivin, G.; Moiroux, J.; Brodeur, J. Behavioural effects of temperature on ectothermic animals: Unifying thermal physiology and behavioural plasticity. Biol. Rev. 2017, 92, 1859–1876. [Google Scholar]
- Evans, R.K.; Toews, M.D.; Sial, A.A. Impact of short- and long-term heat stress on reproductive potential of drosophila Suzukii matsumura (Diptera: Drosophilidae). J. Therm. Biol. 2018, 78, 92–99. [Google Scholar]
- Dowd, W.W.; King, F.A.; Denny, M.W. Thermal Variation, Thermal extremes and the physiological performance of individuals. J. Therm. Biol. 2015, 218, 1956–1967. [Google Scholar]
- Malik, T.G.; Jarrett, B.J.M.; Sun, S.-J. The effect of experimental warming on reproductive performance and parental care in the burying beetle Nicrophorus nepalensis. R. Soc. Open Sci. 2024, 11, 240653. [Google Scholar] [CrossRef]
- Keller, M.L.; Howard, D.R.; Hall, C.L. The thermal ecology of burying beetles: Temperature influences reproduction and daily activity in Nicrophorus marginatus. Ecol. Entomol. 2021, 46, 1266–1272. [Google Scholar]
- Pellissier Scott, M.; Traniello, J.F.A. Behavioural Cues Trigger ovarian development in the burying beetle, Nicrophorus tomentosus. J. Insect Physiol. 1987, 33, 693–696. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Male Pre-Hatching Care | Male Post-Hatching Care | Female Pre-Hatching Care | Female Post-Hatching Care | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Est. ± SE | LRχ²/z | p | Est. ± SE | LRχ²/z | p | Est. ± SE | LRχ²/z | p | LRχ² | p | |
| 1. Anova test for GLMs | |||||||||||
| Carcass-preparation investment | 7.38 | 0.02 | 18.80 | <0.001 | 0.78 | 0.68 | 5.65 | 0.06 | |||
| Ambient temperature | 0.19 | 0.67 | 131.39 | <0.001 | 0.02 | 0.88 | 263.86 | <0.001 | |||
| Interaction | 10.76 | 0.005 | 6.91 | 0.03 | |||||||
| 2. post-hoc test | |||||||||||
| Control-Reduced | 0.28 ± 0.12 | 2.30 | 0.04 | ||||||||
| Control-Elevated | 0.26 ± 0.12 | 2.15 | 0.06 | ||||||||
| Control: 20–23 °C | 1.13 ± 0.11 | 9.84 | <0.001 | 0.14 ± 0.15 | 0.97 | 0.33 | |||||
| Reduced: 20–23 °C | 0.48 ± 0.16 | 2.95 | 0.003 | 0.23 ± 0.20 | 1.15 | 0.25 | |||||
| Elevated: 20–23 °C | 0.91 ± 0.17 | 5.52 | <0.001 | −0.43 ± 0.20 | −2.15 | 0.03 | |||||
| 20 °C: Control-Reduced | 0.71 ± 0.13 | 5.33 | <0.001 | −0.05 ± 0.17 | −0.31 | 0.94 | |||||
| 20 °C: Control-Elevated | 0.21 ± 0.13 | 1.69 | 0.17 | 0.36 ± 0.17 | 2.17 | 0.06 | |||||
| 23 °C: Control-Reduced | 0.06 ± 0.15 | 0.41 | 0.90 | 0.03 ± 0.18 | 0.19 | 0.98 | |||||
| 23 °C: Control-Elevated | −0.002 ± 0.16 | −0.01 | 1.00 | −0.21 ± 0.19 | −1.15 | 0.44 | |||||
| Variables | Clutch Size | Brood Size | Brood Mass | Mean Larval Mass | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Est. ± SE | z | p | Est. ± SE | z | p | Est. ± SE | t | p | Est. ± SE | t | p | |
| Control: 20–23 °C | 0.22 ± 0.07 | 2.91 | 0.004 | 0.23 ± 0.08 | 2.87 | 0.004 | 1.41 ± 0.24 | 5.89 | <0.001 | 0.02 ± 0.003 | 4.50 | <0.001 |
| Reduced: 20–23 °C | −0.29 ± 0.11 | −2.53 | 0.01 | 0.26 ± 0.10 | 2.52 | 0.01 | 1.00 ± 0.30 | 3.37 | 0.001 | 0.01 ± 0.005 | 3.15 | 0.002 |
| Elevated: 20–23 °C | 0.08 ± 0.11 | 0.71 | 0.48 | −0.14 ± 0.11 | −1.34 | 0.19 | 0.20 ± 0.31 | 0.63 | 0.53 | 0.04 ± 0.005 | 7.32 | <0.001 |
| 20 °C: Control-Reduced | 0.76 ± 0.10 | 7.80 | <0.001 | 0.08 ± 0.08 | 0.93 | 0.58 | 0.61 ± 0.25 | 2.41 | 0.03 | 0.008 ± 0.004 | 1.94 | 0.11 |
| 20 °C: Control-Elevated | 0.15 ± 0.09 | 1.59 | 0.21 | 0.32 ± 0.08 | 3.90 | <0.001 | 1.03 ± 0.25 | 4.12 | <0.001 | −0.019 ± 0.004 | −4.71 | <0.001 |
| 23 °C: Control-Reduced | 0.25 ± 0.10 | 2.63 | 0.02 | 0.11± 0.09 | 1.16 | 0.43 | 0.20 ± 0.26 | 0.79 | 0.68 | 0.006 ± 0.004 | 1.43 | 0.28 |
| 23 °C: Control-Elevated | 0.009 ± 0.10 | 0.09 | 0.99 | −0.05 ± 0.09 | −0.54 | 0.83 | −0.19 ± 0.26 | −0.72 | 0.72 | 0.0005 ± 0.004 | 0.11 | 0.99 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, D.; Ma, L.; Komdeur, J. Temperature Variation Regulates the Trade-Off Between Pre- and Post-Hatching Investment in a Burying Beetle. Insects 2025, 16, 378. https://doi.org/10.3390/insects16040378
Ma D, Ma L, Komdeur J. Temperature Variation Regulates the Trade-Off Between Pre- and Post-Hatching Investment in a Burying Beetle. Insects. 2025; 16(4):378. https://doi.org/10.3390/insects16040378
Chicago/Turabian StyleMa, Donghui, Long Ma, and Jan Komdeur. 2025. "Temperature Variation Regulates the Trade-Off Between Pre- and Post-Hatching Investment in a Burying Beetle" Insects 16, no. 4: 378. https://doi.org/10.3390/insects16040378
APA StyleMa, D., Ma, L., & Komdeur, J. (2025). Temperature Variation Regulates the Trade-Off Between Pre- and Post-Hatching Investment in a Burying Beetle. Insects, 16(4), 378. https://doi.org/10.3390/insects16040378