
Isogroup Selection to Optimize Biocontrol Increases Cannibalism in Omnivorous (Zoophytophagous) Bugs


Abstract
1. Introduction
2. Methods
2.1. Populations and Rearing Conditions
2.2. Cannibalism Tests
2.3. Statistical Analysis
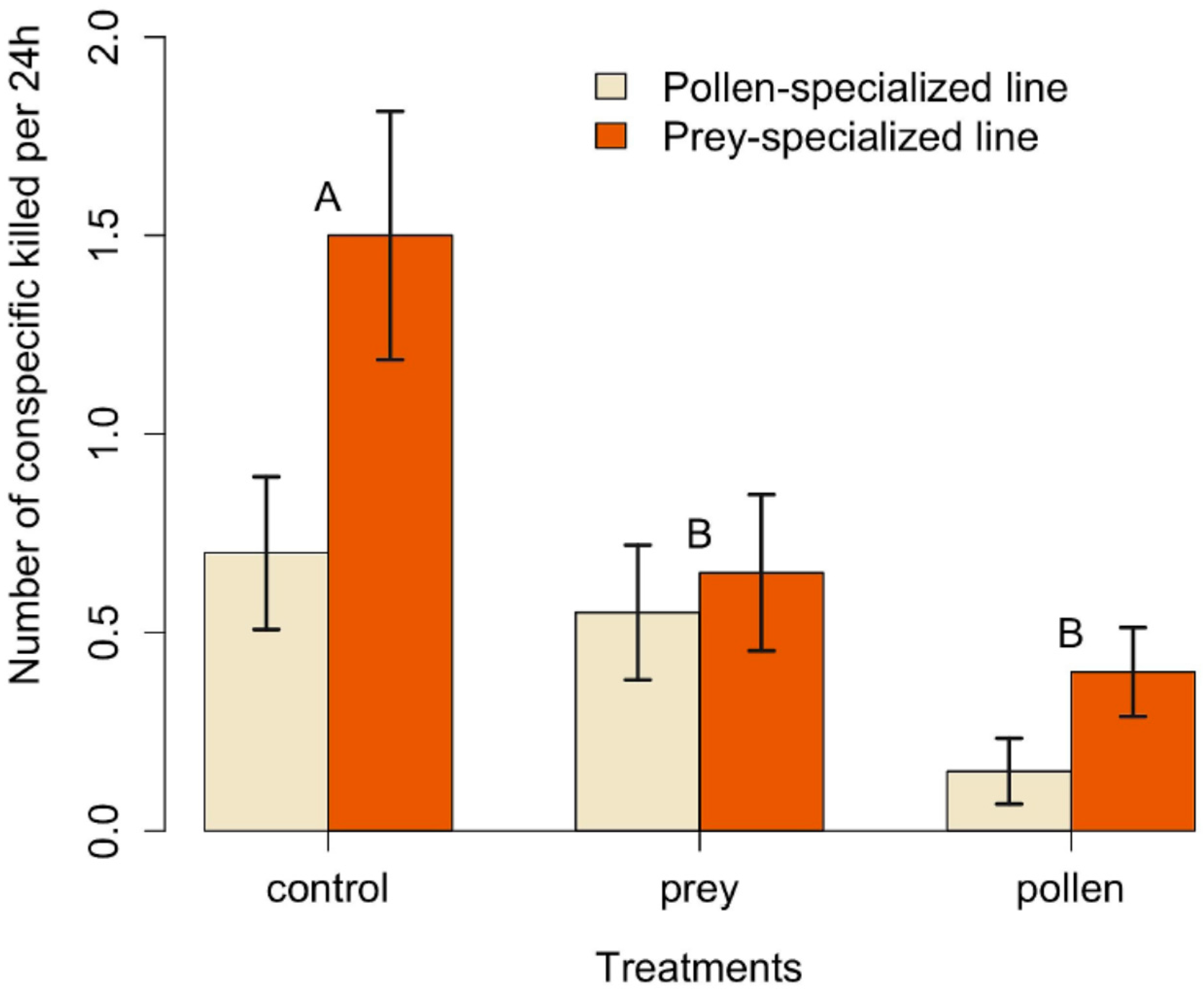
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fox, L.R. Cannibalism in natural populations. Ann. Rev. Ecol. Syst. 1975, 6, 87–106. [Google Scholar] [CrossRef]
- Polis, G.A. The evolution and dynamics of intraspecific predation. Ann. Rev. Ecol. Syst. 1981. [Google Scholar] [CrossRef]
- Ullyett, G.C. Competition for food and allied phenomena in sheep blowfly population. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1950, 234, 77–174. [Google Scholar] [CrossRef]
- Snyder, W.E.; Joseph, S.B.; Preziosi, R.F.; Moore, A.J. Nutritional benefits of cannibalism for the lady beetle Harmonia axyridis (Coleoptera: Coccinellidae) when prey quality is poor. Environ. Entomol. 2000, 29, 1173–1179. [Google Scholar] [CrossRef]
- Mayntz, D.; Toft, S. Nutritional value of cannibalism and the role of starvation and nutrient imbalance for cannibalistic tendencies in a generalist predator. J. Anim. Ecol. 2006, 75, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Leon-Beck, M.; Coll, M. Plant and prey consumption cause a similar reductions in cannibalism by an omnivorous bug. J. Insect Behav. 2007, 20, 67–76. [Google Scholar] [CrossRef]
- Wagner, J.D.; Wise, D.H. Influence of prey availability and conspecifics on patch quality for a cannibalistic forager: laboratory experiments with the wolf spider Schizocosa. Oecologia 1997, 109, 474–482. [Google Scholar] [CrossRef]
- Hironori, Y.; Katsuhiro, S. Cannibalism and interspecific predation in two predatory ladybirds in relation to prey abundance in the field. Entomophaga 1997, 42, 153–163. [Google Scholar] [CrossRef]
- Cottrell, T.E.; Yeargan, K.V. Effect of pollen on Coleomegilla maculata (Coleoptera: Coccinellidae) population density, predation, and cannibalism in sweet corn. Environ. Entomol. 1998, 27, 1402–1410. [Google Scholar] [CrossRef]
- Dumont, F.; Lucas, E.; Réale, D. Evidence of genetic basis of zoophagy and nymphal developmental time in isogroup lines of the zoophytophagous mullein bug, Campylomma verbasci (Hemiptera: Miridae). BioControl 2016. [Google Scholar] [CrossRef]
- Dumont, F.; Lucas, E.; Réale, D. Coexistence of phytozoophagous and zoophytophagous strategies linked to genotypic diet specialization in mullein bug Campylomma verbasci. PLoS ONE 2017, 12, e0176369. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science and Business Media: New York, NY, USA, 2009. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Crawley, M.J. The R Book; John Wiley & Sons: New York, NY, USA, 2012. [Google Scholar]
- Herberich, E.; Sikorski, J.; Hothorn, T. A robust procedure for comparing multiple means under heteroscedasticity in unbalanced designs. PLoS ONE 2010, 5, e9788. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 21 July 2017).
- Wagner, J.D.; Wise, D.H. Cannibalism regulates densities of young wolf spiders: Evidence from field and laboratory experiments. Ecology 1996, 77, 639–652. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Svanbäck, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The ecology of individuals: incidence and implications of individual specialization. Am. Nat. 2003, 161, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Svanbäck, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Pruitt, J.N.; Riechert, S.E. The ecological consequences of temperament in spiders. Curr. Zool. 2012, 58, 588–595. [Google Scholar] [CrossRef]
- Claessen, D.; de Roos, A.M.; Persson, L. Population dynamic theory of size-dependent cannibalism. Proc. R. Soc. B. 2004, 271, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Wise, D.H. Cannibalism, food limitation, intraspecific competition, and the regulation of spider populations. Annu. Rev. Entomol. 2006, 51, 441–465. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, V.H. The interaction of cannibalism and omnivory: Consequences for community dynamics. Ecology 2007, 88, 2697–2705. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.B.; Barros, E.M.; Coelho, R.R.; Pimentel, R.M.M. Zoophytophagous pentatomids feeding on plants and implications for biological control. Arth. Plant Int. 2010, 4, 219–227. [Google Scholar] [CrossRef]
- Castañé, C.; Arno, J.; Gabarra, R.; Alomar, O. Plant damage to vegetable crops by zoophytophagous mirid predators. Biolog. Control 2011, 59, 22–29. [Google Scholar] [CrossRef]
- Arnoldi, D.; Stewart, R.K.; Boivin, G. Predatory mirids of the green apple aphid Aphid pomi, the two-spotted spider mite Tetranychus urticae and the European red spider mite Panonychus ulmi in apple orchards in Québec. Entomophaga 1992, 37, 283–292. [Google Scholar] [CrossRef]
- Boivin, G.; Stewart, R.K. Identification and evaluations of damage to McIntosh apples by phytophagous mirids (Hemiptera: Miridae) in southwestern Québec. Can. Entomol. 1982, 114, 1037–1045. [Google Scholar] [CrossRef]
- Kain, D.P.; Agnello, A.M. Relationship between plant phenology and Campylomma verbasci (Hemiptera: Miridae) damage to apple fruit. Environ. Entomol. 2013, 42, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.A. Zoophytophagy in the plantbug Nesidiocoris tenuis. Agric. Forest Entomol. 2008, 10, 75–80. [Google Scholar] [CrossRef]
- Aubry, O.; Cormier, D.; Chouinard, G.; Lucas, E. Influence of extraguild prey and intraguild predators on the phytophagy of the zoophytophagous bug Campylomma verbasci. J. Pest Sci. 2017, 90, 287–297. [Google Scholar] [CrossRef]
- Bartlett, D. Feeding and egg laying behaviour in Campylomma verbasci Meyer (Hemiptera: Miridae). Master’s Thesis, Simon Fraser University, Burnaby, Canada, 1996. [Google Scholar]
- Aubry, O.; Cormier, D.; Chouinard, G.; Lucas, E. Influence of plant, animal and mixed resources on development of the zoophytophagous plant bug Campylomma verbasci (Hemiptera: Miridae). Biocontrol Sci. Technol. 2015, 25, 1426–1442. [Google Scholar] [CrossRef]
- Pels, B.; Sabelis, M.W. Local dynamics, overexploitation and predator dispersal in an acarine predator-prey system. Oikos 1999, 86, 573–583. [Google Scholar] [CrossRef]
- Hoy, M.A. The potential for genetic improvement of predators for pest management programs. In Genetics in Relation to Insect Management; Rockefeller Foundation Press: New York, NY, USA, 1979; pp. 106–115. [Google Scholar]
- Segal, D.; Glazer, I. Genetics for improving biological control agents: The case of entomopathogenic nematodes. Crop Prot. 2000, 19, 685–689. [Google Scholar] [CrossRef]


{kind=link}
| Comparisons | Estimates ± s.e. | z-Value | p-Value |
|---|---|---|---|
| Prey–Control | −0.61 ± 0.25 | −2.39 | 0.04 |
| Pollen–Control | −1.39 ± 0.34 | −4.11 | <0.001 |
| Prey–Pollen | 0.78 ± 0.36 | 2.14 | 0.08 |
| Treatment | Estimates (± SD) | z-Value | p-Value |
|---|---|---|---|
| Prey-Specialized Line | |||
| Pollen | −1.32 ± 0.40 | −3.32 | 0.0009 |
| Prey | −0.84 ± 0.33 | −2.52 | 0.01 |
| Pollen-Specialized Line | |||
| Pollen | −1.54 ± 0.64 | −2.42 | 0.02 |
| Prey | −0.24 ± 0.40 | −0.60 | 0.55 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumont, F.; Réale, D.; Lucas, E. Isogroup Selection to Optimize Biocontrol Increases Cannibalism in Omnivorous (Zoophytophagous) Bugs. Insects 2017, 8, 74. https://doi.org/10.3390/insects8030074
Dumont F, Réale D, Lucas E. Isogroup Selection to Optimize Biocontrol Increases Cannibalism in Omnivorous (Zoophytophagous) Bugs. Insects. 2017; 8(3):74. https://doi.org/10.3390/insects8030074
Chicago/Turabian StyleDumont, François, Denis Réale, and Eric Lucas. 2017. "Isogroup Selection to Optimize Biocontrol Increases Cannibalism in Omnivorous (Zoophytophagous) Bugs" Insects 8, no. 3: 74. https://doi.org/10.3390/insects8030074
APA StyleDumont, F., Réale, D., & Lucas, E. (2017). Isogroup Selection to Optimize Biocontrol Increases Cannibalism in Omnivorous (Zoophytophagous) Bugs. Insects, 8(3), 74. https://doi.org/10.3390/insects8030074