An Advanced View on Baculovirus per Os Infectivity Factors



Abstract
1. Introduction
2. Definition and General Features of PIFs
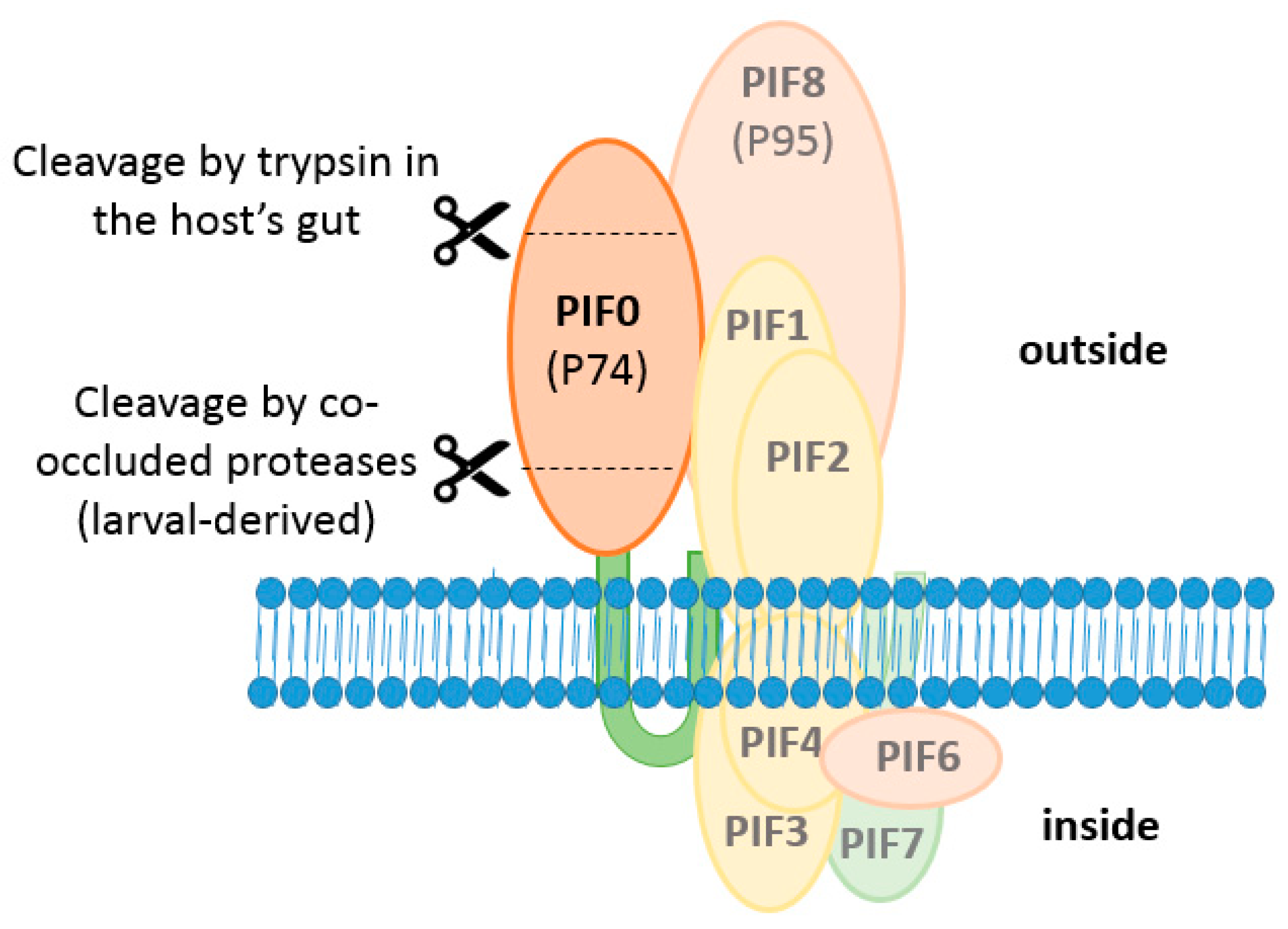
3. The AcMNPV PIF-Complex
4. PIFs Mediate Viral Entry under Alkaline Conditions in the Midgut
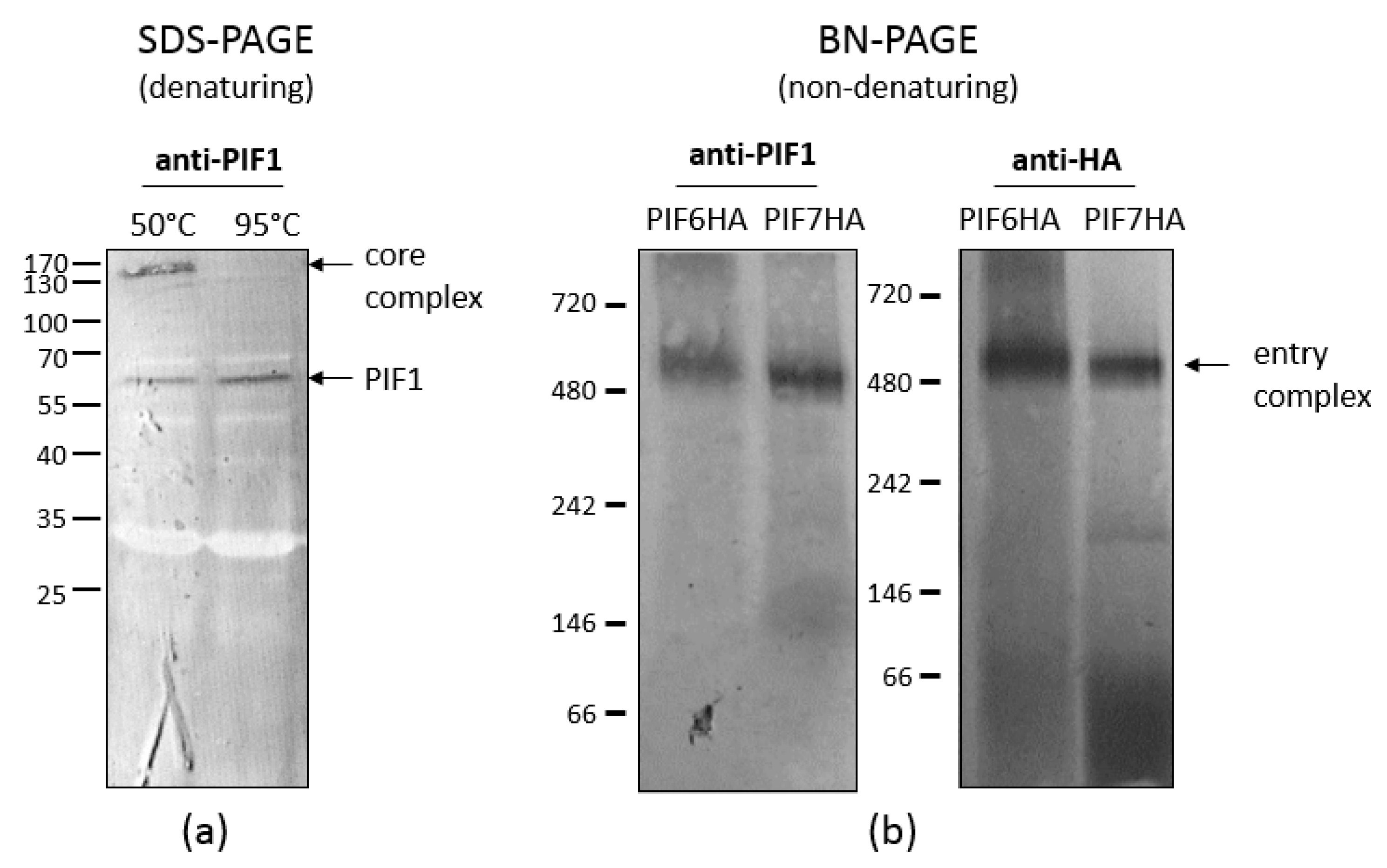
5. PIFs of the Stable Core of the ODV Entry Complex
5.1. PIF1 and 2 Mediate Binding to the Columnar Cell Microvilli
5.2. PIF3 Mediate Oral Infectivity after Binding and Fusion
5.3. The Role of PIF1-3 in Host-Range Determination
5.4. PIF4 Provides Proteolytic Resistance to the Stable Core
6. PIFs That Associate with Lower Affinity to the Core Complex
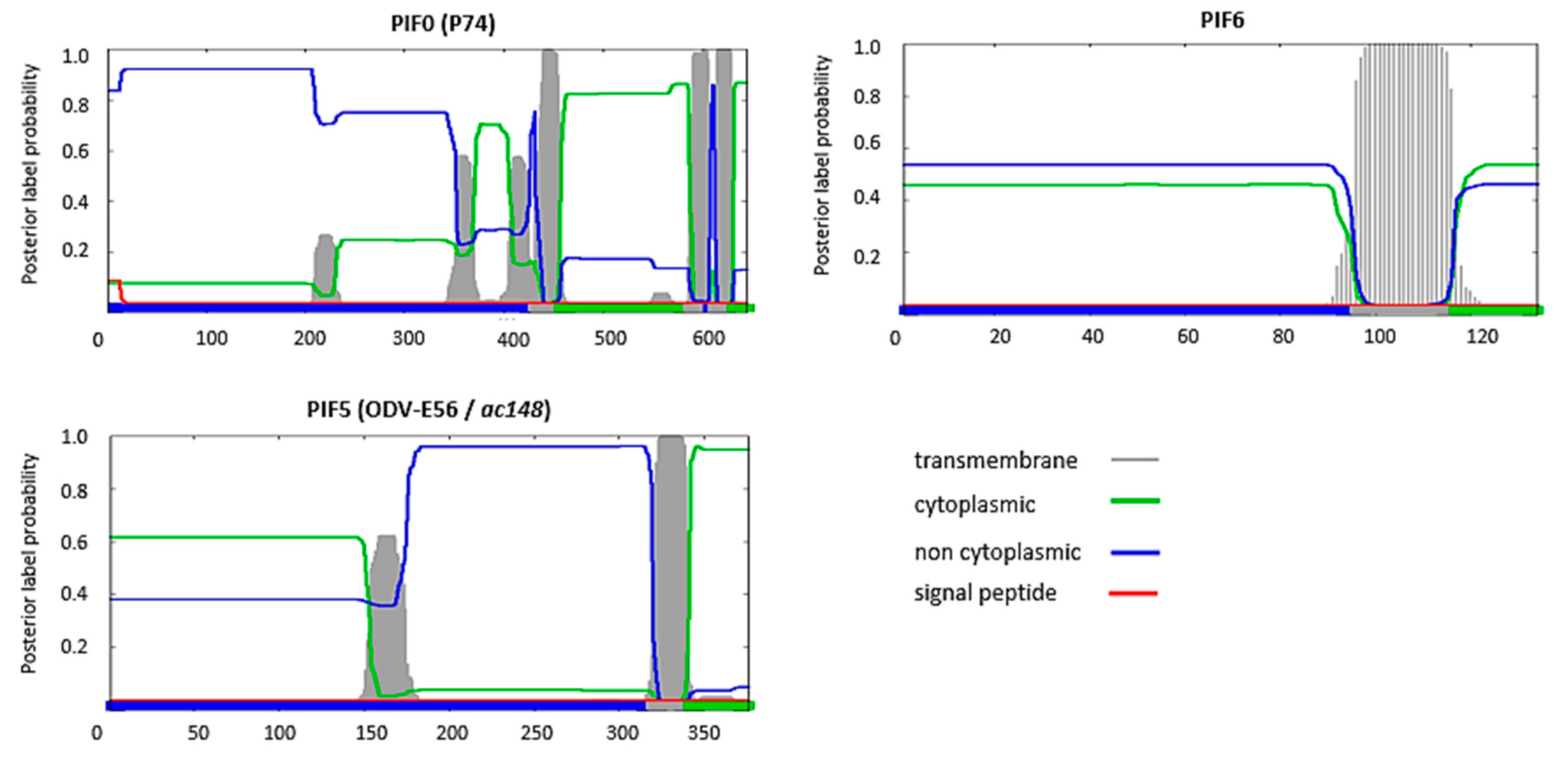
6.1. PIF0 Mediates ODV Binding with the Midgut Epithelium
6.2. PIF0 Is Cleaved by Co-Occluded Proteases and Trypsins
6.3. PIF6 Is Also a Component of the ODV Entry Complex
6.4. The Zinc-Finger Domain of PIF8 Is Important for Oral Infectivity
7. PIF5 Might Operate Independent from the ODV Entry Complex
8. ODV-E66 Might Retrospectively Be Assigned as PIF9
9. Conclusions and Perspectives
Funding
Conflicts of Interest
References
- Williams, T.; Bergoin, M.; van Oers, M.M. Diversity of large DNA viruses of invertebrates. J. Invertebr. Pathol. 2017, 147, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, B.; Rabalski, L.; Krol, E.; Sihler, W.; Souza, M. Baculovirus biopesticides–a safe alternative to chemical protection of plants. J. Biopest. 2009, 2, 209–216. [Google Scholar]
- Moscardi, F. A nucleopolyhedrovirus for control of the velvet bean caterpillar in brazilian soybeans. In Biological Control: A Global Perspective; CABI International: Wallingford, UK, 2007; pp. 344–352. ISBN 184-593-266-8. [Google Scholar]
- Peng, K.; van Lent, J.W.; Boeren, S.; Fang, M.; Theilmann, D.A.; Erlandson, M.A.; Vlak, J.M.; van Oers, M.M. Characterization of novel components of the baculovirus per os infectivity factor complex. J. Virol. 2012, 86, 4981–4988. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; van Oers, M.M.; Hu, Z.; van Lent, J.W.; Vlak, J.M. Baculovirus per os infectivity factors form a complex on the surface of occlusion-derived virus. J. Virol. 2010, 84, 9497–9504. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, P.; Kuzio, J.; Williams, G.V.; Wilson, J.A. Analysis of p74, a pdv envelope protein of autographa californica nucleopolyhedrovirus required for occlusion body infectivity in vivo. J. Gen. Virol. 1997, 78, 3091–3100. [Google Scholar] [CrossRef] [PubMed]
- Kikhno, I.; Gutierrez, S.; Croizier, L.; Croizier, G.; Ferber, M.L. Characterization of pif, a gene required for the per os infectivity of spodoptera littoralis nucleopolyhedrovirus. J. Gen. Virol. 2002, 83, 3013–3022. [Google Scholar] [CrossRef] [PubMed]
- Pijlman, G.P.; Pruijssers, A.J.; Vlak, J.M. Identification of pif-2, a third conserved baculovirus gene required for per os infection of insects. J. Gen. Virol. 2003, 84, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; van Oers, M.M.; Donly, B.C.; Erlandson, M.A.; Hegedus, D.D.; Theilmann, D.A. Acmnpv ac83 is a pif protein required for odv and bv nucleocapsid assembly as well as assembly of the pif complex in odv envelopes. J. Virol. 2016. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.; Makalliwa, G.A.; Li, J.; Wang, H.; Hu, Z.; Wang, M. Per os infectivity factors: A complicated and evolutionarily conserved entry machinery of baculovirus. Sci. China Life Sci. 2017, 60, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, T.; Washburn, J.O.; Sitapara, R.; Sid, E.; Volkman, L.E. Specific binding of autographa californica m nucleopolyhedrovirus occlusion-derived virus to midgut cells of heliothis virescens larvae is mediated by products of pif genes ac119 and ac022 but not by ac115. J. Virol. 2005, 79, 15258–15264. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.; Nie, Y.; Harris, S.; Erlandson, M.A.; Theilmann, D.A. Autographa californica multiple nucleopolyhedrovirus core gene ac96 encodes a per os infectivity factor (pif-4). J. Virol. 2009, 83, 12569–12578. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Sparks, W.O.; Bonning, B.C. Autographa californica multiple nucleopolyhedrovirus odv-e56 envelope protein is required for oral infectivity and can be substituted functionally by rachiplusia ou multiple nucleopolyhedrovirus odv-e56. J. Gen. Virol. 2010, 91, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Fang, M.; Erlandson, M.A.; Theilmann, D.A. Analysis of the autographa californica multiple nucleopolyhedrovirus overlapping gene pair lef3 and ac68 reveals that ac68 is a per os infectivity factor and that lef3 is critical but not essential, for virus replication. J. Virol. 2012, 86, 3985–3994. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, L.; Zhang, S.; Deng, Z.; Huang, Z.; Yuan, M.; Wu, W.; Yang, K. The autographa californica multiple nucleopolyhedrovirus ac110 gene encodes a new per os infectivity factor. Virus Res. 2016, 221, 30–37. [Google Scholar]
- Zhu, S.; Wang, W.; Wang, Y.; Yuan, M.; Yang, K. The baculovirus core gene ac83 is required for nucleocapsid assembly and per os infectivity of autographa californica nucleopolyhedrovirus. J. Virol. 2013, 87, 10573–10586. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L. A combined transmembrane topology and signal peptide prediction method. J. Mol. Biol. 2004, 338, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Braunagel, S.C.; Williamson, S.T.; Saksena, S.; Zhong, Z.; Russell, W.K.; Russell, D.H.; Summers, M.D. Trafficking of odv-e66 is mediated via a sorting motif and other viral proteins: Facilitated trafficking to the inner nuclear membrane. Proc. Natl. Acad. Sci. USA 2004, 101, 8372–8377. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Song, J.; Jiang, T.; Liang, C.; Chen, X. The n-terminal hydrophobic sequence of autographa californica nucleopolyhedrovirus pif-3 is essential for oral infection. Arch. Virol. 2007, 152, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Boogaard, B.; van Lent, J.W.M.; Theilmann, D.A.; Erlandson, M.A.; van Oers, M.M. Baculoviruses require an intact odv entry-complex to resist proteolytic degradation of per os infectivity factors by co-occluded proteases from the larval host. J. Gen. Virol. 2017, 98, 3101–3110. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, C.; Zhang, N.; Li, J.; Deng, F.; Wang, H.; Vlak, J.M.; Hu, Z.; Wang, M. The group i alphabaculovirus-specific protein, ac5, is a novel component of the occlusion body but is not associated with odvs or the pif complex. J. Gen. Virol. 2018, 99, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Terra, W.R.; Ferreira, C. Insect digestive enzymes: Properties, compartmentalization and function. Comp. Biochem. Physiol. B Comp. Biochem. 1994, 109, 1–62. [Google Scholar] [CrossRef]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Slavicek, J.M. Baculovirus enhancins and their role in viral pathogenicity. In Molecular Virology; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Toprak, U.; Harris, S.; Baldwin, D.; Theilmann, D.; Gillott, C.; Hegedus, D.D.; Erlandson, M.A. Role of enhancin in mamestra configurata nucleopolyhedrovirus virulence: Selective degradation of host peritrophic matrix proteins. J. Gen. Virol. 2012, 93, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Granados, R.R. An intestinal mucin is the target substrate for a baculovirus enhancin. Proc. Natl. Acad. Sci. USA 1997, 94, 6977–6982. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G.F. Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. [Google Scholar]
- Granados, R.R.; Lawler, K.A. In vivo pathway of autographa californica baculovirus invasion and infection. Virology 1981, 108, 297–308. [Google Scholar] [CrossRef]
- Kawanishi, C.; Summers, M.; Stoltz, D.; Arnott, H. Entry of an insect virus in vivo by fusion of viral envelope and microvillus membrane. J. Invertebr. Pathol. 1972, 20, 104–108. [Google Scholar] [CrossRef]
- Horton, H.M.; Burand, J.P. Saturable attachment sites for polyhedron-derived baculovirus on insect cells and evidence for entry via direct membrane fusion. J. Virol. 1993, 67, 1860–1868. [Google Scholar] [PubMed]
- Hoekstra, D.; de Boer, T.; Klappe, K.; Wilschut, J. Fluorescence method for measuring the kinetics of fusion between biological membranes. Biochemistry 1984, 23, 5675–5681. [Google Scholar] [CrossRef] [PubMed]
- Blissard, G.W.; Wenz, J.R. Baculovirus gp64 envelope glycoprotein is sufficient to mediate ph-dependent membrane fusion. J. Virol. 1992, 66, 6829–6835. [Google Scholar] [PubMed]
- Monsma, S.A.; Blissard, G.W. Identification of a membrane fusion domain and an oligomerization domain in the baculovirus gp64 envelope fusion protein. J. Virol. 1995, 69, 2583–2595. [Google Scholar] [PubMed]
- Wilson, N.F.; Snell, W.J. Microvilli and cell-cell fusion during fertilization. Trends Cell Biol. 1998, 8, 93–96. [Google Scholar] [CrossRef]
- Mu, J.; van Lent, J.W.; Smagghe, G.; Wang, Y.; Chen, X.; Vlak, J.M.; van Oers, M.M. Live imaging of baculovirus infection of midgut epithelium cells: A functional assay of per os infectivity factors. J. Gen. Virol. 2014, 95, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Li, K.; Tang, P.; Li, Y.; Zhou, Q.; Yang, K.; Zhang, Q. Three-dimensional visualization of the autographa californica multiple nucleopolyhedrovirus occlusion-derived virion envelopment process gives new clues as to its mechanism. Virology 2015, 476, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, M.; Huang, H.; Luo, X.; Deng, F.; Wang, H.; Hu, Z. Functional studies of per os infectivity factor 3 of helicoverpa armigera nucleopolyhedrovirus. J. Gen. Virol. 2012, 93, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Haas-Stapleton, E.J.; Washburn, J.O.; Volkman, L.E. Spodoptera frugiperda resistance to oral infection by autographa californica multiple nucleopolyhedrovirus linked to aberrant occlusion-derived virus binding in the midgut. J. Gen. Virol. 2005, 86, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, X.; Hou, D.; Huang, H.; Liu, X.; Deng, F.; Wang, H.; Arif, B.M.; Hu, Z.; Wang, M. The host specificities of baculovirus per os infectivity factors. PLoS ONE 2016, 11, e0159862. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.-Q.; Zhang, J.; Chen, X.-M.; He, Q.; Cao, M.-Y.; Wang, L.; Li, H.-Q.; Xiao, W.-F.; Pan, C.-X.; Lu, C. Bombyx mori nucleopolyhedrovirus orf79 is a per os infectivity factor associated with the pif complex. Virus Res. 2014, 184, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, M.; Deng, F.; Wang, H.; Hu, Z. Orf85 of hearnpv encodes the per os infectivity factor 4 (pif4) and is essential for the formation of the pif complex. Virology 2012, 427, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Braunagel, S.; Russell, W.; Rosas-Acosta, G.; Russell, D.; Summers, M. Determination of the protein composition of the occlusion-derived virus of autographa californica nucleopolyhedrovirus. Proc. Natl. Acad. Sci. USA 2003, 100, 9797–9802. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, K.; Cai, K.; Yao, Q. Determination of protein composition and host-derived proteins of bombyx mori nucleopolyhedrovirus by 2-dimensional electrophoresis and mass spectrometry. Intervirology 2008, 51, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Deng, F.; Hou, D.; Zhao, Y.; Guo, L.; Wang, H.; Hu, Z. Proteomics of the autographa californica nucleopolyhedrovirus budded virions. J. Virol. 2010, 84, 7233–7242. [Google Scholar] [CrossRef] [PubMed]
- Kuzio, J.; Jaques, R.T.; Faulkner, P. Identification of p74, a gene essential for virulence of baculovirus occlusion bodies. Virology 1989, 173, 759–763. [Google Scholar] [CrossRef]
- Slack, J.M.; Dougherty, E.M.; Lawrence, S.D. A study of the autographa californica multiple nucleopolyhedrovirus odv envelope protein p74 using a gfp tag. J. Gen. Virol. 2001, 82, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.M.; Lawrence, S.D.; Krell, P.J.; Arif, B.M. A soluble form of p74 can act as a per os infectivity factor to the autographa californica multiple nucleopolyhedrovirus. J. Gen. Virol. 2010, 91, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Zhou, W.; Xu, H.; Zheng, Y.; Qi, Y. The heliothis armigera single nucleocapsid nucleopolyhedrovirus envelope protein p74 is required for infection of the host midgut. Virus Res. 2004, 104, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Haas-Stapleton, E.J.; Washburn, J.O.; Volkman, L.E. P74 mediates specific binding of autographa californica m nucleopolyhedrovirus occlusion-derived virus to primary cellular targets in the midgut epithelia of heliothis virescens larvae. J. Virol. 2004, 78, 6786–6791. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; van Lent, J.W.; Vlak, J.M.; Hu, Z.; van Oers, M.M. In situ cleavage of baculovirus occlusion-derived virus receptor binding protein p74 in the peroral infectivity complex. J. Virol. 2011, 85, 10710–10718. [Google Scholar] [CrossRef] [PubMed]
- Bonning, B.C.; Hoover, K.; Duffey, S.; Hammock, B.D. Production of polyhedra of the autographa-californica nuclear polyhedrosis-virus using the sf21 and tn5b1-4 cell-lines and comparison with host-derived polyhedra by bioassay. J. Invertebr. Pathol. 1995, 66, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.M.; Lawrence, S.D. Evidence for proteolytic cleavage of the baculovirus occlusion-derived virion envelope protein p74. J. Gen. Virol. 2005, 86, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Slack, J.M.; Lawrence, S.D.; Krell, P.J.; Arif, B.M. Trypsin cleavage of the baculovirus occlusion-derived virus attachment protein p74 is prerequisite in per os infection. J. Gen. Virol. 2008, 89, 2388–2397. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, R.; Deng, F.; Wang, H.; Hu, Z. Functional studies of per os infectivity factors of helicoverpa armigera single nucleocapsid nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 2331–2338. [Google Scholar] [CrossRef] [PubMed]
- Rohrmann, G. Baculovirus Molecular Biology; National Center for Biotechnology Information: Bethesda, MD, USA, 2011. [Google Scholar]
- Sparks, W.O.; Harrison, R.L.; Bonning, B.C. Autographa californica multiple nucleopolyhedrovirus odv-e56 is a per os infectivity factor but is not essential for binding and fusion of occlusion-derived virus to the host midgut. Virology 2011, 409, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Braunagel, S.C.; Elton, D.M.; Ma, H.; Summers, M.D. Identification and analysis of an autographa californica nuclear polyhedrosis virus structural protein of the occlusion-derived virus envelope: Odv-e56. Virology 1996, 217, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Wu, M.; Deng, F.; Song, J.; Dong, C.; Wang, H.; Hu, Z. Identification of protein-protein interactions of the occlusion-derived virus-associated proteins of helicoverpa armigera nucleopolyhedrovirus. J. Gen. Virol. 2010, 91, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Sparks, W.O.; Rohlfing, A.; Bonning, B.C. A peptide with similarity to baculovirus odv-e66 binds the gut epithelium of heliothis virescens and impedes infection with autographa californica multiple nucleopolyhedrovirus. J. Gen. Virol. 2011, 92, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Chen, L.; Hu, X.; Yu, S.; Yang, R.; Wu, X. Autographa californica multiple nucleopolyhedrovirus odv-e66 is an essential gene required for oral infectivity. Virus Res. 2011, 158, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Thézé, J.; Bézier, A.; Periquet, G.; Drezen, J.-M.; Herniou, E.A. Paleozoic origin of insect large dsdna viruses. Proc. Natl. Acad. Sci. USA 2011, 108, 15931–15935. [Google Scholar] [CrossRef] [PubMed]
- Casartelli, M.; Corti, P.; Leonardi, M.G.; Fiandra, L.; Burlini, N.; Pennacchio, F.; Giordana, B. Absorption of albumin by the midgut of a lepidopteran larva. J. Insect Physiol. 2005, 51, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Grenet, A.G.; Castelli, I.; Cermenati, G.; Ravallec, M.; Fiandra, L.; Debaisieux, S.; Multeau, C.; Lautredou, N.; Dupressoir, T. Densovirus crosses the insect midgut by transcytosis and disturbs the epithelial barrier function. J. Virol. 2013, 87, 12380–12391. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PIF | ORF | Size (kDa) | Nr. Of Cys. | Reference |
|---|---|---|---|---|
| PIF0 (P74) | ac138 | 74 | 6 | Faulkner et al. [6] |
| PIF1 | ac119 | 60 | 24 | Kikhno et al. [7] |
| PIF2 | ac22 | 44 | 14 | Pijlman et al. [8] |
| PIF3 | ac115 | 23 | 12 | Ohkawa et al. [11] |
| PIF4 | ac96 | 20 | 2 | Fang et al. [12] |
| PIF5 (ODV-E56) | ac148 | 56 | 6 | Harrison et al. [13] |
| PIF6 | ac68 | 16 | 1 | Nie et al. [14] |
| PIF7 | ac110 | 7 | 1 | Liu et al. [15] |
| PIF8 (VP91/P95) | ac83 | 96 | 13 | Zhu et al. [16] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boogaard, B.; Van Oers, M.M.; Van Lent, J.W.M. An Advanced View on Baculovirus per Os Infectivity Factors. Insects 2018, 9, 84. https://doi.org/10.3390/insects9030084
Boogaard B, Van Oers MM, Van Lent JWM. An Advanced View on Baculovirus per Os Infectivity Factors. Insects. 2018; 9(3):84. https://doi.org/10.3390/insects9030084
Chicago/Turabian StyleBoogaard, Bob, Monique M. Van Oers, and Jan W. M. Van Lent. 2018. "An Advanced View on Baculovirus per Os Infectivity Factors" Insects 9, no. 3: 84. https://doi.org/10.3390/insects9030084
APA StyleBoogaard, B., Van Oers, M. M., & Van Lent, J. W. M. (2018). An Advanced View on Baculovirus per Os Infectivity Factors. Insects, 9(3), 84. https://doi.org/10.3390/insects9030084