A Biocontrol Strain of Pseudomonas aeruginosa CQ-40 Promote Growth and Control Botrytis cinerea in Tomato


Abstract
:1. Introduction
2. Results
2.1. Isolation, Purification and Screening of Antagonistic Strains
2.2. Identification of Strain CQ-4
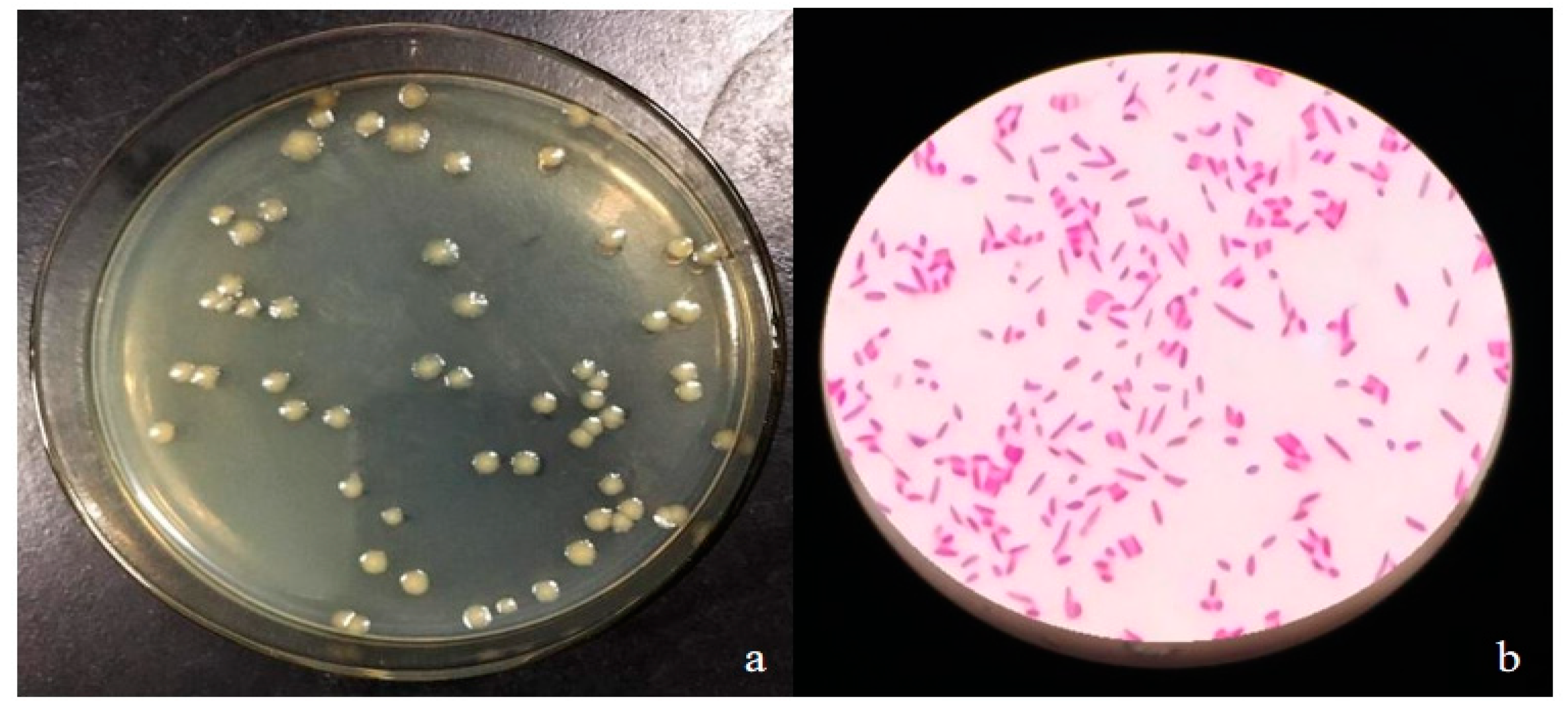
2.2.1. StrainCQ-4 Morphology Detection
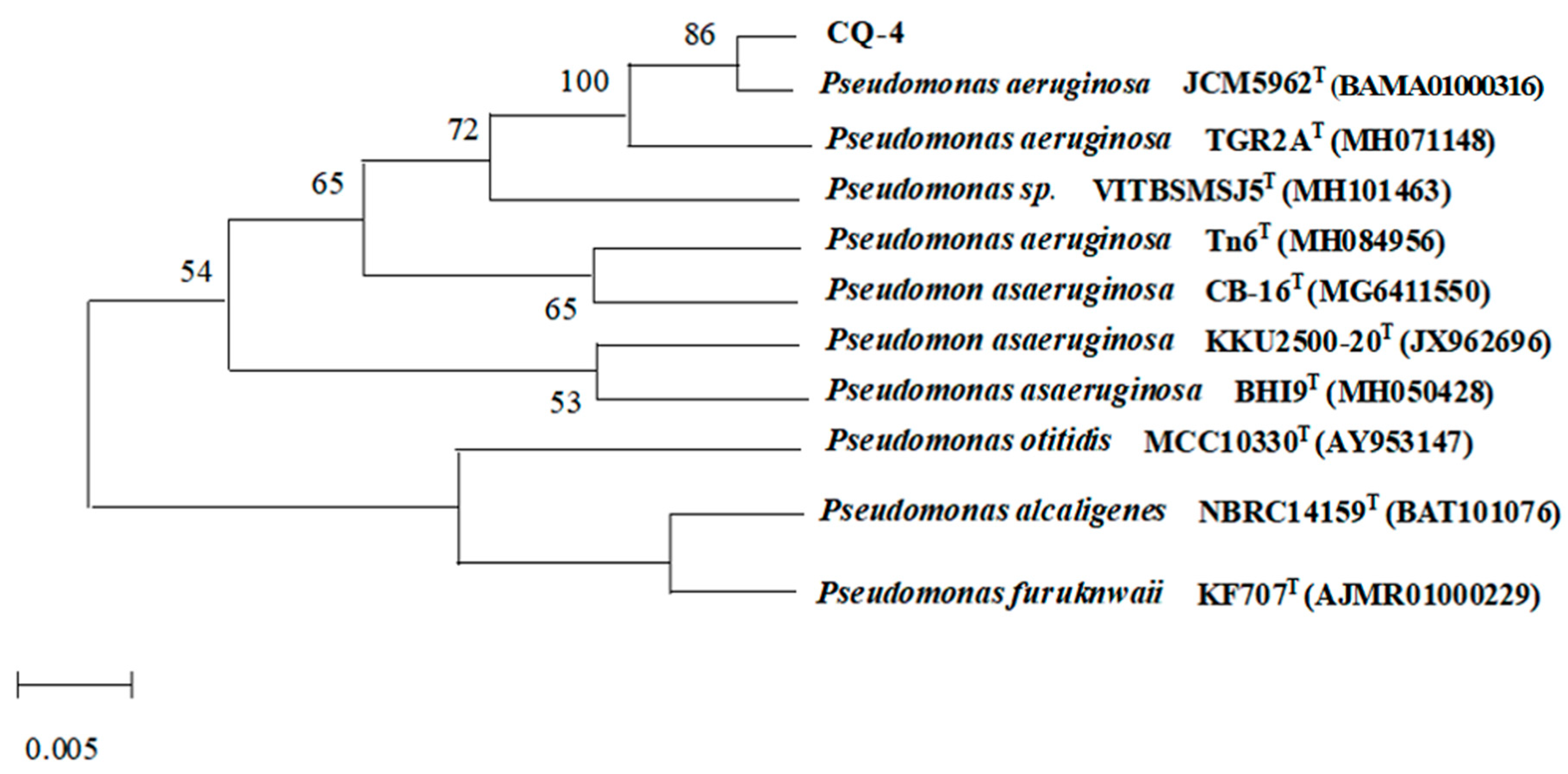
2.2.2. Molecular Identifification of Bacterial Strain CQ-4
2.2.3. Physicochemical Identifification of Bacterial Strain CQ-4
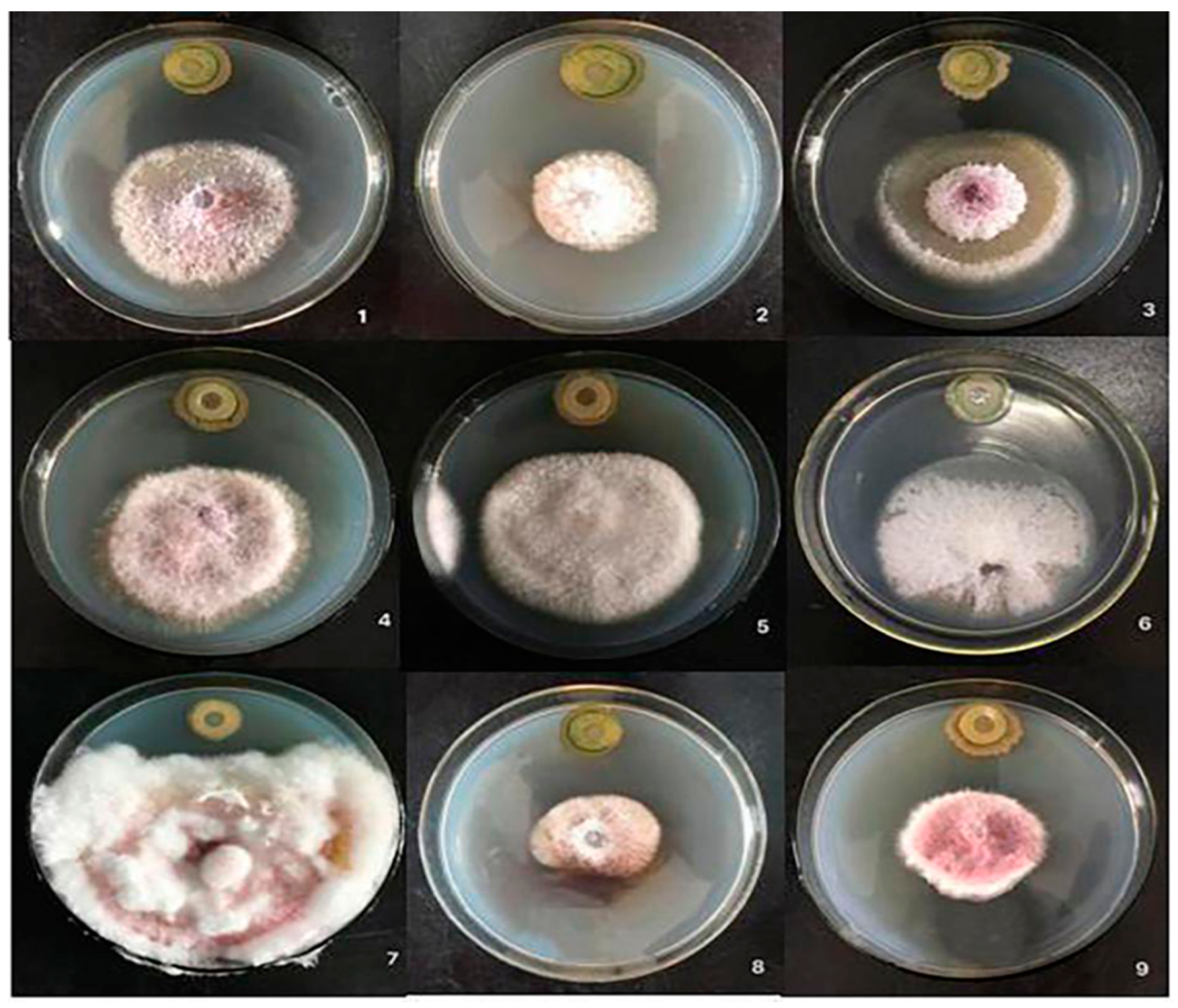
2.3. Determination of Antibacterial Spectrum of Biocontrol Bacteria CQ-4
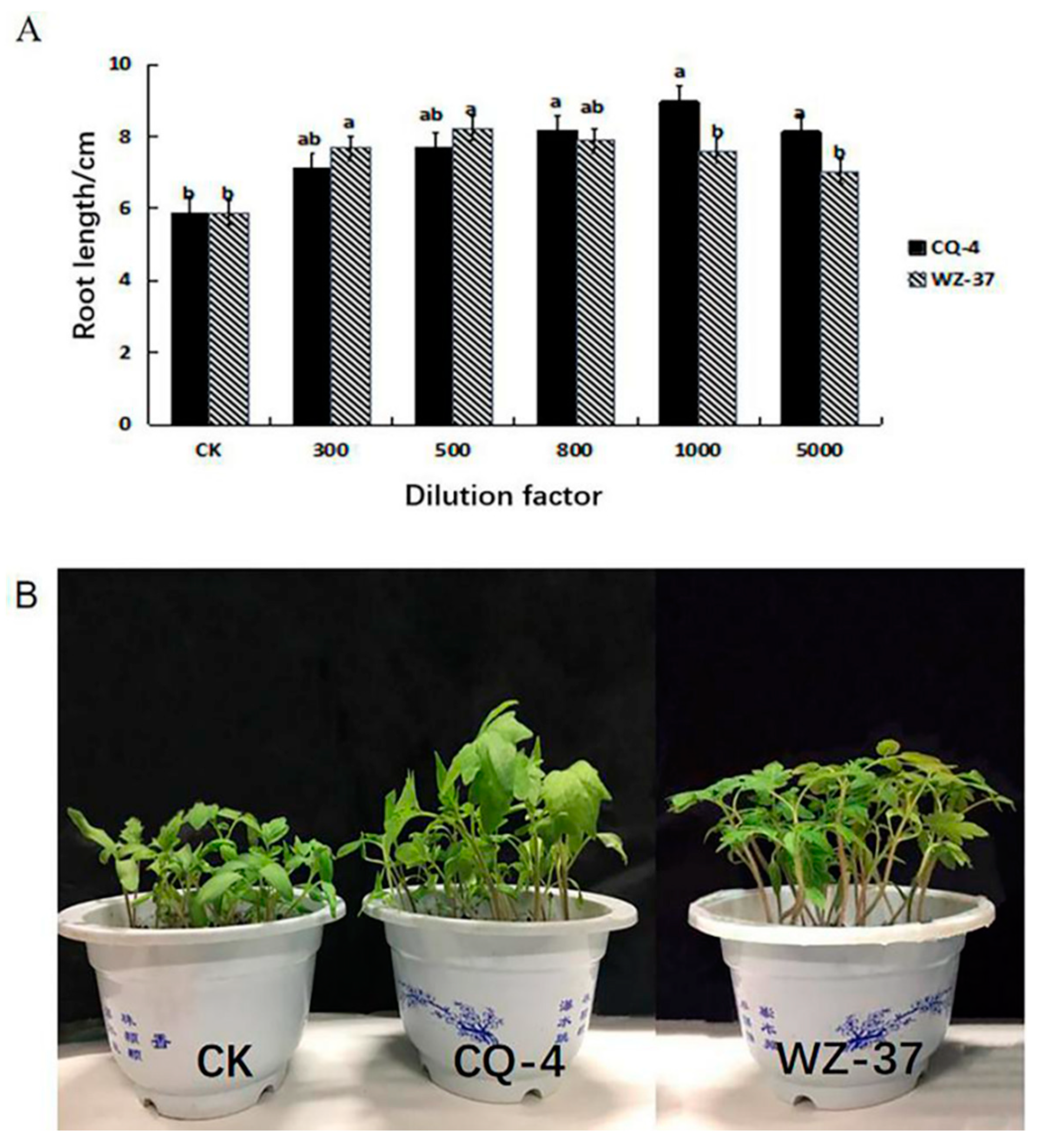
2.4. Determination of the Effect of Biocontrol Bacteria CQ-4 on Tomato Growth Promotion
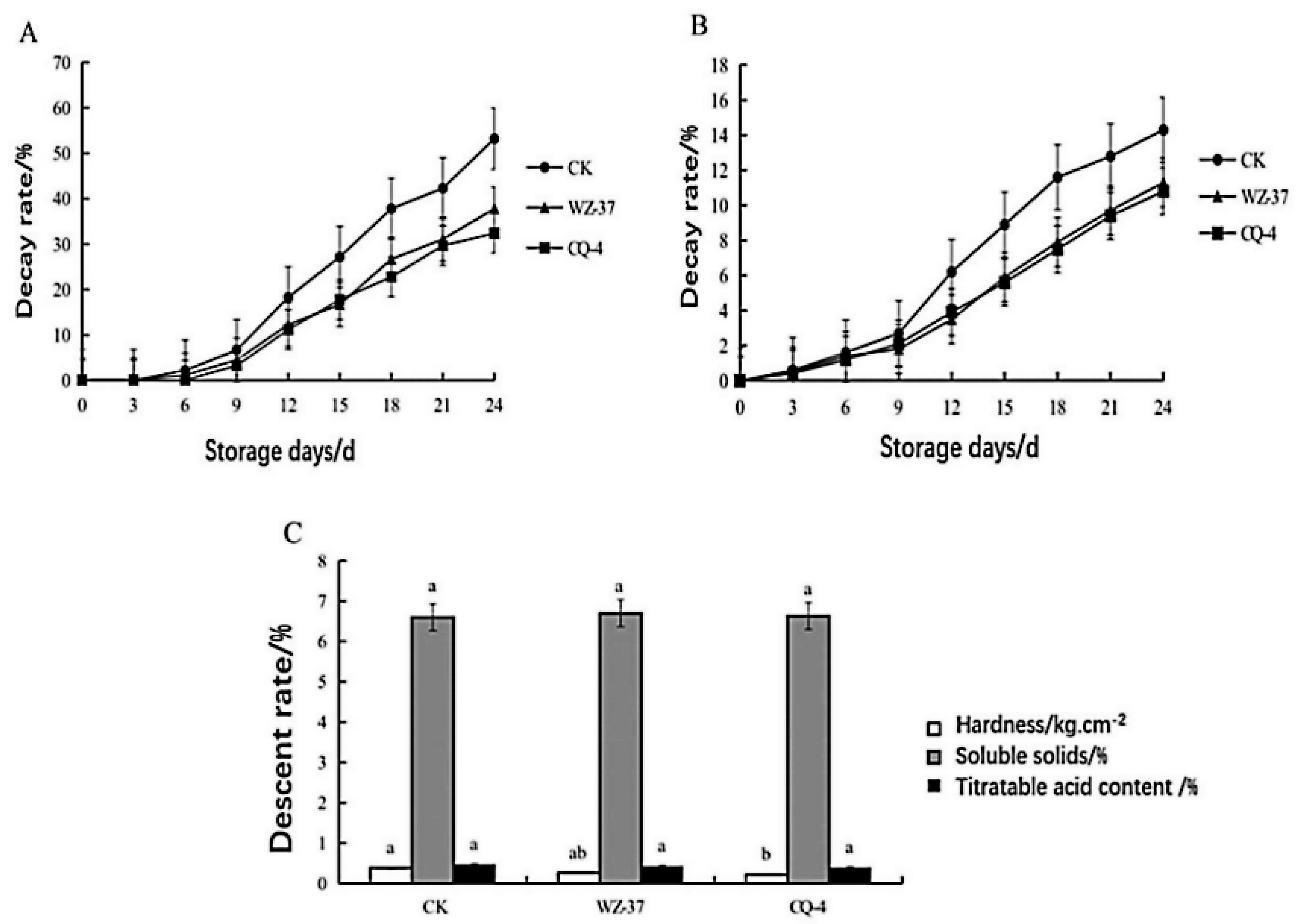
2.5. The Biocontrol Effect of Strain CQ-4 on Tomato Gray Mold
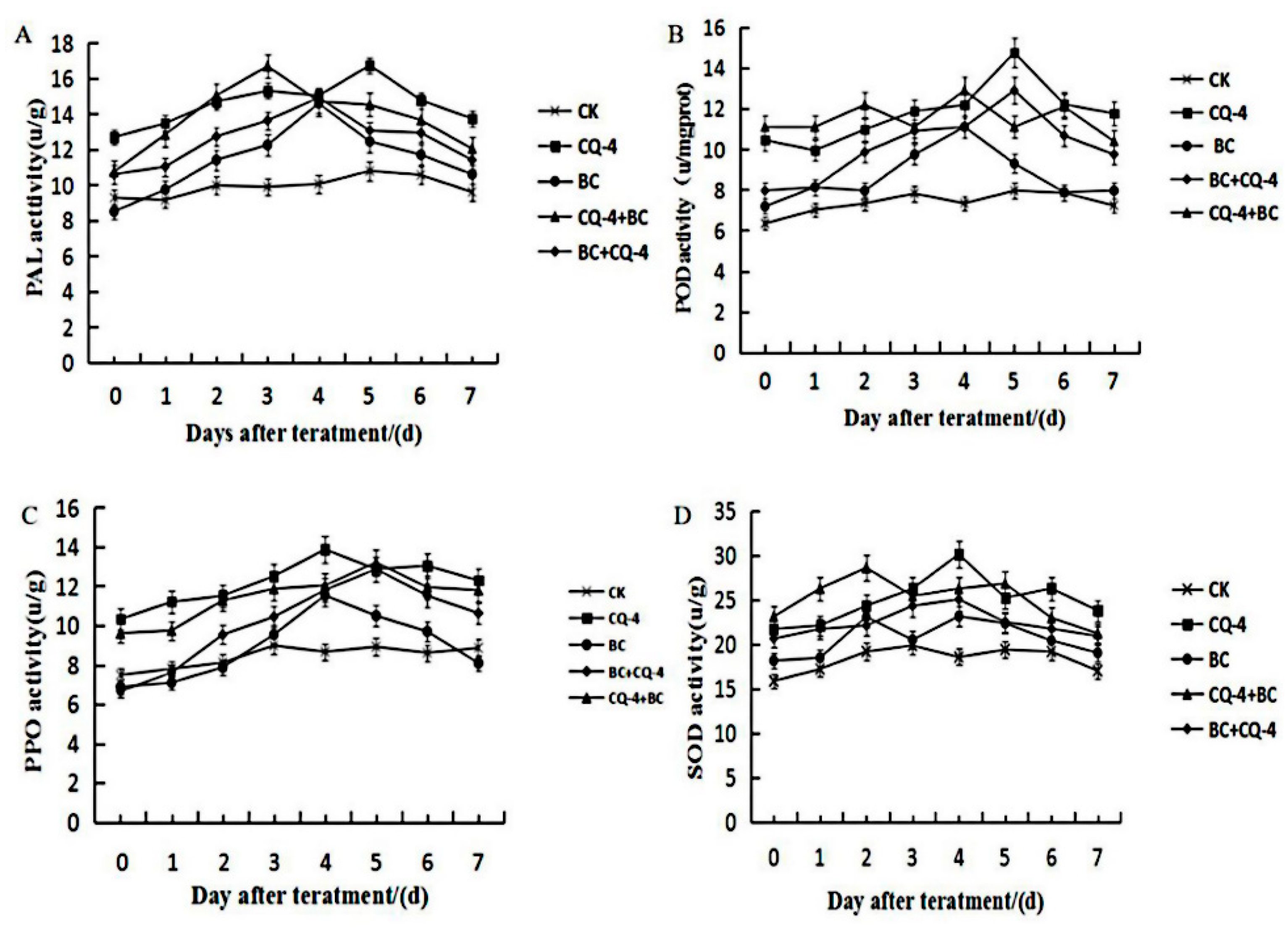
2.6. Detection of Defense Enzyme activities Induced by Biocontrol Bacteria CQ-4 in Tomato Leaves
2.6.1. Effect of Phenylalanine Ammonia Lyase (PAL) Activity on Tomato Leaves after Treatment with Biocontrol Bacteria
2.6.2. Effect of Peroxidase (POD) Activity on Tomato Leaves after Treatment with Biocontrol Bacteria
2.6.3. Effect of Polyphenol Oxidase (PPO) Activity on Tomato Leaves after Treatment with Biocontrol Bacteria
2.6.4. Effect of Superoxide Dismutase (SOD) Activity on Tomato Leaves after Treatment with Biocontrol Bacteria
2.7. Study on the Related Mechanism of Biocontrol Bacteria in Promoting Growth and Disease Resistance
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Separation, Purification and Screening of Biocontrol Bacteria
4.3. Identification of Biocontrol Strain CQ-4
4.4. Determination of Antibacterial Spectrum of Biocontrol Bacteria CQ-4
4.5. In Vitro Control Effect of Biocontrol Bacteria CQ-4 on Tomato Gray Mold
4.6. The Effect of the Biocontrol Bacteria CQ-4 on the Greenhouse Potted Control of Tomato Gray Mold
4.7. Growth-Promoting Effect of Biocontrol Bacteria CQ-4 on Tomato Seeds and Seedlings
4.8. Effect of Biocontrol Bacteria CQ-4 on the Quality of Postharvest Tomato Fruits
4.9. Effect of Biocontrol Bacteria on Inducing Defensive Enzymes Related to Tomato Leaf Resistance
4.10. Determination of the Effects of Biocontrol Bacteria on Plant Growth and Disease Resistance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botero, D.; Alvarado, C.; Bernal, A. Network Analyses in Plant Pathogens. Front. Microbiol. 2018, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Welter, S.; Lehmann, K.; Dölle, S.; Schwarz, D.; Weckwerth, W.; Scheler, C.; Worm, M.; Franken, P. Identification of putative new tomato allergens and differential interaction with IgEs of tomato allergic subjects. Clin. Exp. Allergy 2013, 43, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.N.; Wang, X.R.; Zhang, L.; Ahammed, G.J.; Li, Q.Y.; Li, X. Epigallocatechin-3-gallate enhances tomato resistance to tobacco mosaic virus by modulating RBOH1-dependent H2O2 signaling. Plant Physiol. Biochem. 2020, 150, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Ge, B.; Wang, J.; Liu, B.; Ma, J.; Wei, Q.; Zhang, K. iTRAQ-based proteomic analysis reveals the mechanisms of Botrytis cinerea controlled with Wuyiencin. BMC Microbiol. 2019, 19, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Powell, A.L.T.; Orlando, R.; Bergmann, C.; Gutierrez-Sanchez, G. Proteomic analysis of ripening tomato fruit infected by Botrytis cinerea. J. Proteome Res. 2012, 11, 2178–2192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fillinger, S.; Elad, Y. Botrytis-the Fungus, the Pathogen and Its Management in Agricultural Systems; Springer: Heidelberg, Germany, 2016; pp. 413–486. [Google Scholar]
- Ji, X.; Li, J.; Meng, Z.; Zhang, S.; Qiao, K. Synergistic Effect of Combined Application of a New Fungicide Fluopimomide with a Biocontrol Agent TA-1 for Management of Gray Mold in Tomato. Plant Dis. 2019, 103, 1991–1997. [Google Scholar] [CrossRef]
- Dik, A.J.; Elad, Y. Comparison of Antagonists of Botrytis cinerea in Greenhouse-grown Cucumber and Tomato under Different Climatic Conditions. Eur. J. Plant Pathol. 1999, 105, 123–137. [Google Scholar] [CrossRef]
- Srivastava, D.A.; Arya, G.C.; Pandaranayaka, E.P.; Manasherova, E.; Prusky, D.B.; Elad, Y.; Harel, A. Botrytis cinerea Transcriptome Profiling Data of Infection on Whole Plant. Mol. Plant Microbe Interact. 2020, 33, 1103–1107. [Google Scholar] [CrossRef]
- Yusoff, S.F.; Haron, F.F.; Tengku, M.M.M.; Asib, N.; Sakimin, S.Z.; Ismail, S.I. Antifungal Activity and Phytochemical Screening of Vernonia amygdalina Extract against Botrytis cinerea Causing Gray Mold Disease on Tomato. Fruits. Biol. 2020, 9, 286. [Google Scholar] [CrossRef]
- Patharkar, O.R. Quantification of Cauline Leaf Abscission in Response to Plant Pathogens. Methods Mol. Biol. 2019, 1991, 127–139. [Google Scholar]
- Taguchi, Y.; Hyakumachi, M.; Tsueda, H. Method for adhering Bacillus subtilis IK-1080 to bumblebees and control of gray mold disease of tomato. Jpn. J. Phytopathol. 2003, 69, 94–101. [Google Scholar] [CrossRef]
- Li, T.T.; Zhang, J.D.; Tang, J.Q.; Liu, Z.C.; Li, Y.Q.; Chen, J.; Zou, L.W. Trichoderma atroviride Combined Use of CCTCCSBW0199 and Brassinolide to Control Infection in Tomato. Plant. Dis. 2020, 104, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Postma, J.; Rattink, H. Biological control of Fusarium wilt disease of carnation by a nonpathogenic isolate of Fusarium oxysporum. Acta Hprtic. 1992, 307, 37–42. [Google Scholar]
- Mahdiyeh, M.; Asghar, H.; Nader, H.; Laleh, N. Evaluation of Pseudomonas and Bacillus Bacterial Antagonists for Biological Control of Cotton Verticillium Wilt Disease. J. Plant. Prot. Res. 2013, 53, 154–157. [Google Scholar]
- Qing, X.Z.; Ying, Z.; Ling, L.H.; Yun, H.T. Identification of a small antimycotic peptide produced by Bacillus amyloliquefaciens 6256. Pestic. Biochem. Physiol. 2018, 150, 78–82. [Google Scholar]
- Qiao, J.; Yu, X.; Liang, X.; Liu, Y.; Borriss, R. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome. BMC Microbiol. 2017, 17, 131. [Google Scholar] [CrossRef]
- Gao, P.; Qin, J.; Zhou, S. Inhibitory effect and possible mechanism of a Pseudomonas strain QBA5 against gray mold on tomato leaves and fruits caused by Botrytis cinerea. PLoS ONE 2018, 13, e0190932. [Google Scholar] [CrossRef] [Green Version]
- Huijuan, Z.; Yating, Q.; Miao, L.; Ming, J. Functions of pipecolic acid on induced resistance against Botrytis cinerea and Pseudomonas syringae pv. tomato DC3000 in tomato plants. J. Phytopathol. 2020, 168, 591–600. [Google Scholar]
- Palleroni, N.J.; Bradbury, J.K. Stren Pseudomonas aeruginosa otrophomonas a new bact-erial genus for xanthornonas maltophllia Swings. Int. J. Syst. Evol. Microbiol. 1993, 43, 606–609. [Google Scholar]
- Jatsenko, T.; Sidorenko, J.; Saumaa, S.; Kivisaar, M. DNA Polymerases ImuC and DinB Are Involved in DNA Alkylation Damage Tolerance in and Pseudomonas putida. PLoS ONE 2017, 12, e0170719. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.J. Making greater use of introduced microorganisms for Biologicalcontrol of plant pathogens. Annu Rev. Phytopathol. 1993, 31, 53–80. [Google Scholar] [CrossRef] [PubMed]
- Parvin, W.; Govender, N.; Othman, R. Phenazine from Pseudomonas aeruginosa UPMP3 induced the host resistance in oil palm (Elaeis guineensis Jacq.)-Ganoderma boninense pathosystem. Sci. Rep. 2020, 10, 15621. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Gao, Y.; Gao, T.; Zhang, D. BacillusInhibitory Abilities of Isolates and Their Culture Filtrates against the Gray Mold Caused by on Postharvest Fruit. Plant Pathol. J. 2019, 35, 425–436. [Google Scholar] [PubMed]
- Viji, G.; Uddin, W.; Romaine, C.P. Suppression of gray leaf spot (blast) of perennial ryegrass turf by Pseudomonas aeruginosa from spent mushroom substrate. Biol. Control 2003, 26, 233–243. [Google Scholar] [CrossRef]
- Guo, D.-J.; Singh, R.K.; Singh, P. Enterobacter roggenkampii Complete Genome Sequence of ED5, a Nitrogen Fixing Plant Growth Promoting Endophytic Bacterium with Biocontrol and Stress Tolerance Properties, Isolated from Sugarcane Root. Front. Microbiol. 2020, 11, 580081. [Google Scholar] [CrossRef]
- Duijff, B.J.; Meijer, J.W.; Bakker, P. Siderophore-mediated competition for iron and induced resistance in the suppression of Fusarium wilt of carnation by fluorescent Pseudomonas spp. Netherland J. Plant Pathol. 1993, 99, 277–289. [Google Scholar] [CrossRef]
- Ge, Y.H.; Huang, X.Q.; Wang, S.L. Phenazine-1-carboxylic acid is negatively regulated and pyoluteorin positively regulated by gac A in Pseudomonas sp. M18. FEMS Microbiol. Lett. 2004, 237, 41–47. [Google Scholar] [CrossRef]
- Wang, S.L.; Yieh, T.C.; Shih, I.L. Production of antifungal comp-ounds by Pseudomonas aerugi nosa K187 using shrimp and crab shellpowder as a carbon source. Enzym. Microb. Technol. 1999, 25, 142–148. [Google Scholar] [CrossRef]
- Peng, R.; Lin, J.; Wei, D. Purification and characterization of an organic solvent-tolerant lipase from Pseudomonas aeruginosa CS-2. Appl. Biochem. Biotechnol. 2010, 162, 733–743. [Google Scholar] [CrossRef]
- Sharon, C.; Furugoh, S.; Yamakido, T.; Ogawa, H.I.; Kato, Y. Purification and characterization of alipase from Pseudomonas aeruginosa KKA-5 and its role in castor oilhydrolysis. J. Ind. Microbiol. Biotechnol. 1998, 20, 304–313. [Google Scholar] [CrossRef]
- Da-Qiang, W.; Jing, Y.; Hong-Yu, O.; Xue, W.; Xianqing, H.; Yuquan, X. Genomic analysis and temperature-dependent transcriptome profiles of the rhizosphere originating strain Pseudomonas aeruginosa M18. BMC Genom. 2011, 12, 438. [Google Scholar]
- Li, Y.L.; Zhou, Z.; Yuan, Y.C.; Ye, J.R. First Report of a Leaf Spot of Radermachera sinica in China Caused by Bacillus megaterium. Plant Dis. 2014, 98, 1425. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- Yu, Y.P.; Gong, T.; Günter, J. Isolation and characterization of five lytic bacteriophages infecting a Vibrio strain closely related to Vibrio owensii. FEMS Microbiol. Lett. 2013, 348, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Zouari, I.; Masmoudi, F.; Medhioub, K. Biocontrol and plant growth-promoting potentiality of bacteria isolated from compost extract. Antonie Van Leeuwenhoek 2020, 113, 2107–2122. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, X.; Yue, J.X.; Tian, D.; Chen, J.Q. Recent duplications dominate nbs-encoding gene expansion in two woody species. Mol. Gen. Genom. 2008, 280, 187. [Google Scholar] [CrossRef]
- Wang, H.; Shi, Y.; Wang, D.; Yao, Z.; Wang, Y.; Liu, J.; Zhang, S.; Wang, A. Bacillus subtilisA Biocontrol Strain of WXCDD105 Used to Control Tomato and Cooke and Promote the Growth of Seedlings. Int. J. Mol. Sci. 2018, 19, 1371. [Google Scholar] [CrossRef] [Green Version]
- Dalia, A.; Cristina, R.; Emilio, S.; Vitale, A.; Gabriella, C. Postharvest biocontrol ability of Pseudomonas synxantha against Monilinia fructicola and Monilini a fructigena on stone fruit. Postharvest Biol. Technol. 2019, 149, 83–89. [Google Scholar]
- Castric, K.F.; Castric, P.A. Method for the rapid detection of cy-anogenic bacteria. Appl. Environ. Microbiol. 1983, 45, 701–712. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Inhibition Zone Width/mm | ||
|---|---|---|---|
| Botrytis cinerea | Fusarium oxysporum | Rhizoctonia solani | |
| 2 | 5.6 ± 0.5 bc | 1.8 ± 0.6 e | 4.8 ± 0.7 b |
| 4 | 9.6 ± 0.3 a | 10.8 ± 0.2 a | 9.3 ± 0.4 a |
| 7 | 6 ± 0.4 b | 2.1 ± 0.4 de | 3.8 ± 0.3 bc |
| 12 | 5.1 ± 0.3 c | 2.3 ± 0.3 d | 3.0 ± 0.3 c |
| 19 | 4.1 ± 0.4 cd | 3.8 ± 0.4 c | 1.8 ± 0.6 cd |
| 71 | 4.6 ± 0.1 c | 3.0 ± 0.5 cd | 4.1 ± 0.1 bc |
| 133 | 3.8 ± 0.4 d | 4.5 ± 0.2 bc | 2.5 ± 0.2 cd |
| 179 | 7.8 ± 0.2 ab | 6.2 ± 0.5 b | 1.3 ± 0.3 d |
| Treatment | Plant Height/mm | Thick Stem/mm | Taproot Length/mm | Fresh Weight/g | Dry Weight/g |
|---|---|---|---|---|---|
| CQ-4 | 80.04 ± 0.3 a | 2.69 ± 0.1 a | 88.59 ± 0.1 a | 1.66 ± 0.1 a | 0.17 ± 0.3 a |
| WZ-37 | 78.56 ± 0.7 ab | 2.07 ± 0.1 b | 71.82 ± 0.7 b | 1.34 ± 0.4 ab | 0.14 ± 0.2 b |
| CK | 69.39 ± 0.8 b | 1.68 ± 0.06 c | 59.32 ± 0.6 c | 0.72 ± 0.3 c | 0.07 ± 0.1 c |
| Treatment | Disease Index (%) | Control Index (%) | ||
|---|---|---|---|---|
| Biocontrol Bacteria + Pathogens | Pathogenic Bacteria + Biocontrol Bacteria | Biocontrol Bacteria + Pathogens | Pathogenic Bacteria + Biocontrol Bacteria | |
| CQ-4 | 20.63 c | 27.12 c | 61.25 ab | 49.06 b |
| WZ-37 | 24.71 b | 28.06 b | 53.58 c | 47.29 c |
| 40% pyrimethanil | 18.97 d | 24.88 d | 64.36 a | 53.26 a |
| CK | 53.24 a | 53.24 a | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Zhou, X.; Cai, Z.; Guo, L.; Chen, X.; Chen, X.; Liu, J.; Feng, M.; Qiu, Y.; Zhang, Y.; et al. A Biocontrol Strain of Pseudomonas aeruginosa CQ-40 Promote Growth and Control Botrytis cinerea in Tomato. Pathogens 2021, 10, 22. https://doi.org/10.3390/pathogens10010022
Wang X, Zhou X, Cai Z, Guo L, Chen X, Chen X, Liu J, Feng M, Qiu Y, Zhang Y, et al. A Biocontrol Strain of Pseudomonas aeruginosa CQ-40 Promote Growth and Control Botrytis cinerea in Tomato. Pathogens. 2021; 10(1):22. https://doi.org/10.3390/pathogens10010022
Chicago/Turabian StyleWang, Xingyuan, Xinan Zhou, Zhibo Cai, Lan Guo, Xiuling Chen, Xu Chen, Jiayin Liu, Mingfang Feng, Youwen Qiu, Yao Zhang, and et al. 2021. "A Biocontrol Strain of Pseudomonas aeruginosa CQ-40 Promote Growth and Control Botrytis cinerea in Tomato" Pathogens 10, no. 1: 22. https://doi.org/10.3390/pathogens10010022