
The Durability of Vaccine Efficacy against Ocular HSV-1 Infection Using ICP0 Mutants 0∆NLS and 0∆RING Is Lost over Time



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. A Comparison of HSV-1 ICP0 Mutants 0∆NLS and 0∆RING to Parental (WT) GFP105 Resistance to IFN-β In Vitro
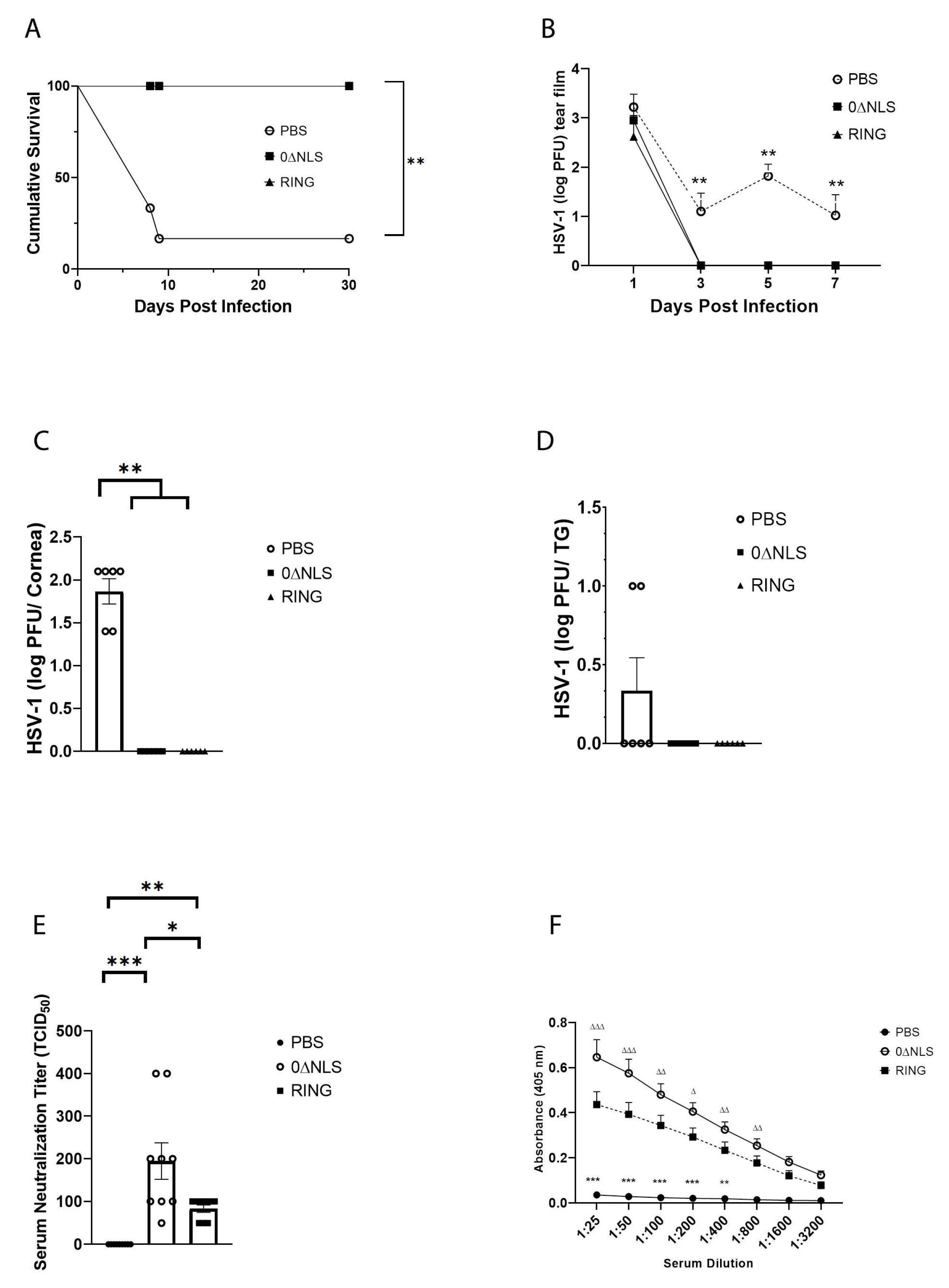
2.2. The ICP0 Mutant 0∆RING Shows Similar Degrees of Efficacy Compared to the ICP0 Mutant 0∆NLS as A Prophylactic Vaccine against HSV-1 in Mice Challenged 30 Days Post Boost
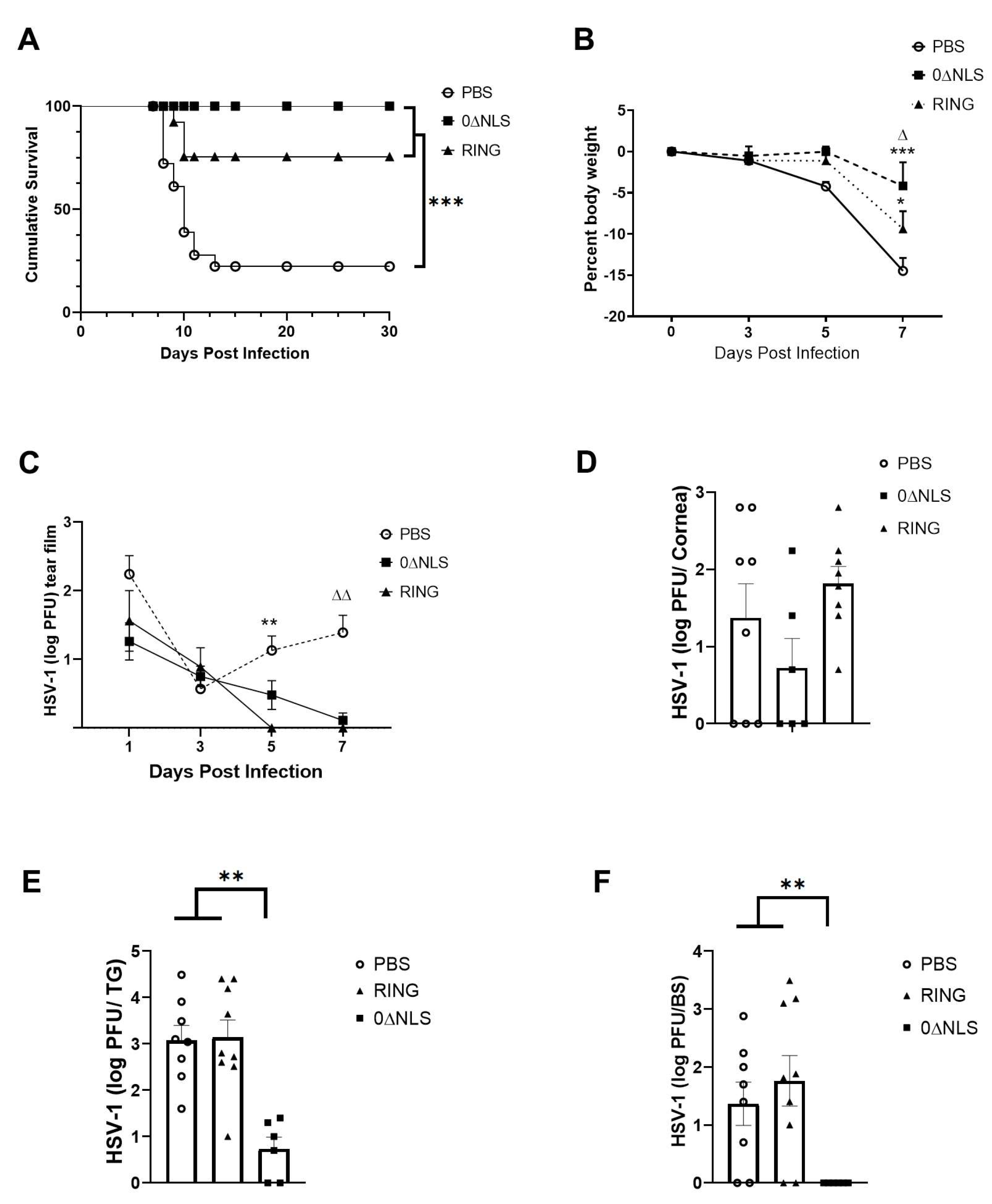
2.3. The HSV-1 0∆NLS Vaccine Displays Superior Efficacy Compared to The HSV-1 0∆RING Vaccine as Determined by Virus Replication and Weight Loss Results following Ocular HSV-1 Challenge in Long-Term Study
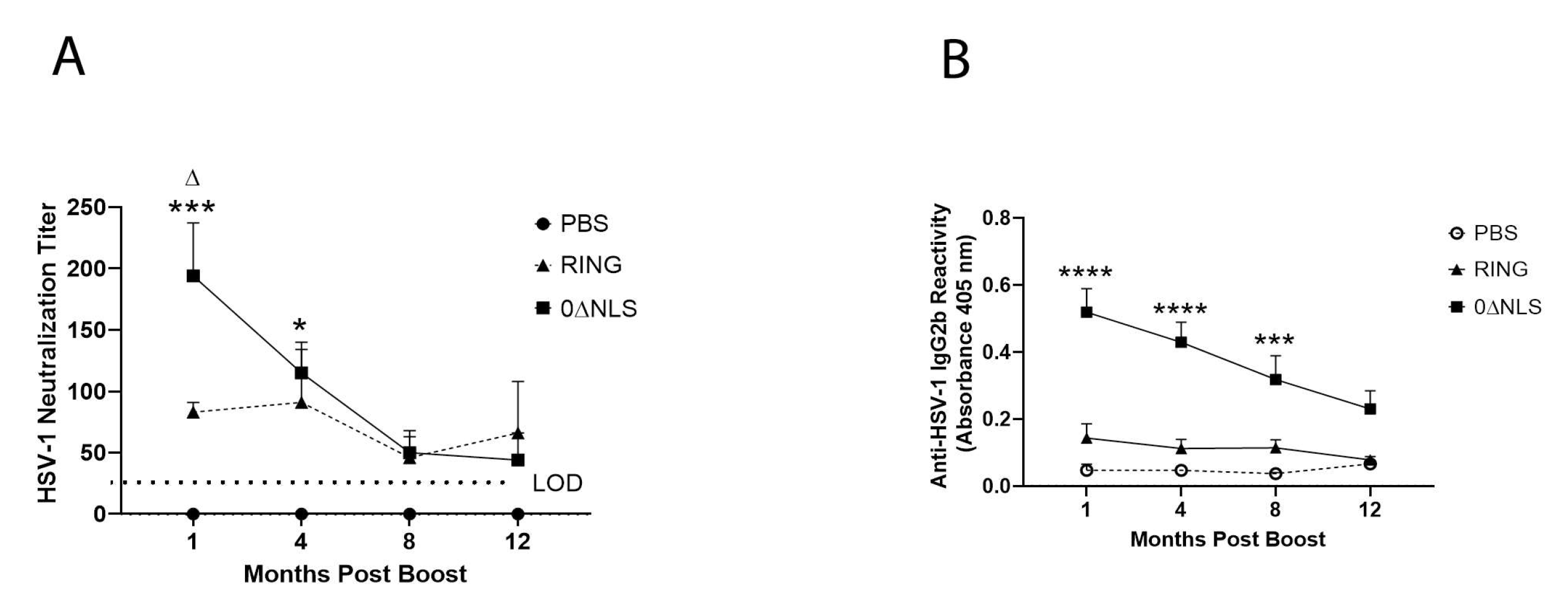
2.4. Anti-HSV-1 Antibody Titers Are Lost over Time following Vaccination with 0∆NLS or 0∆RING
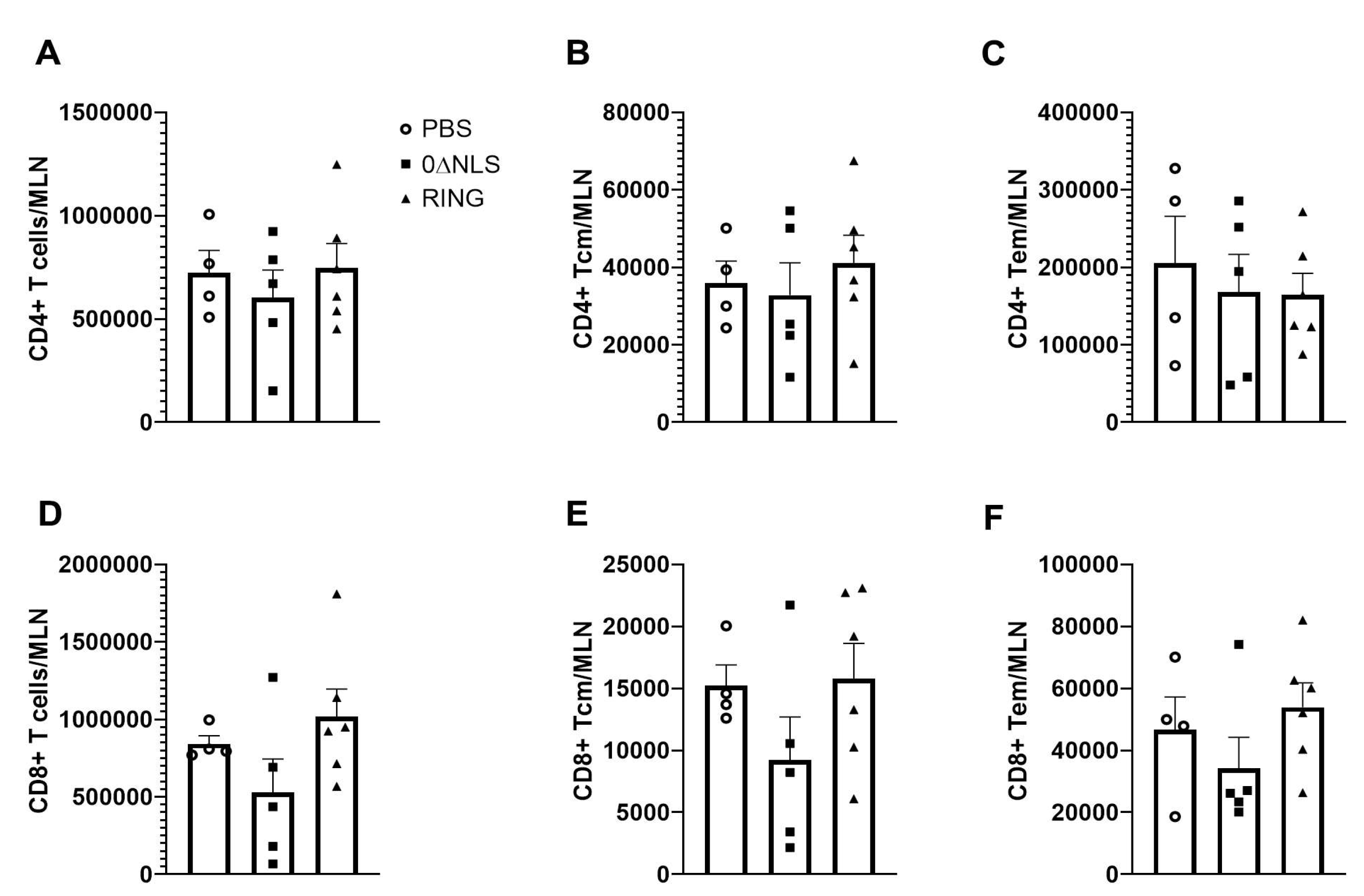
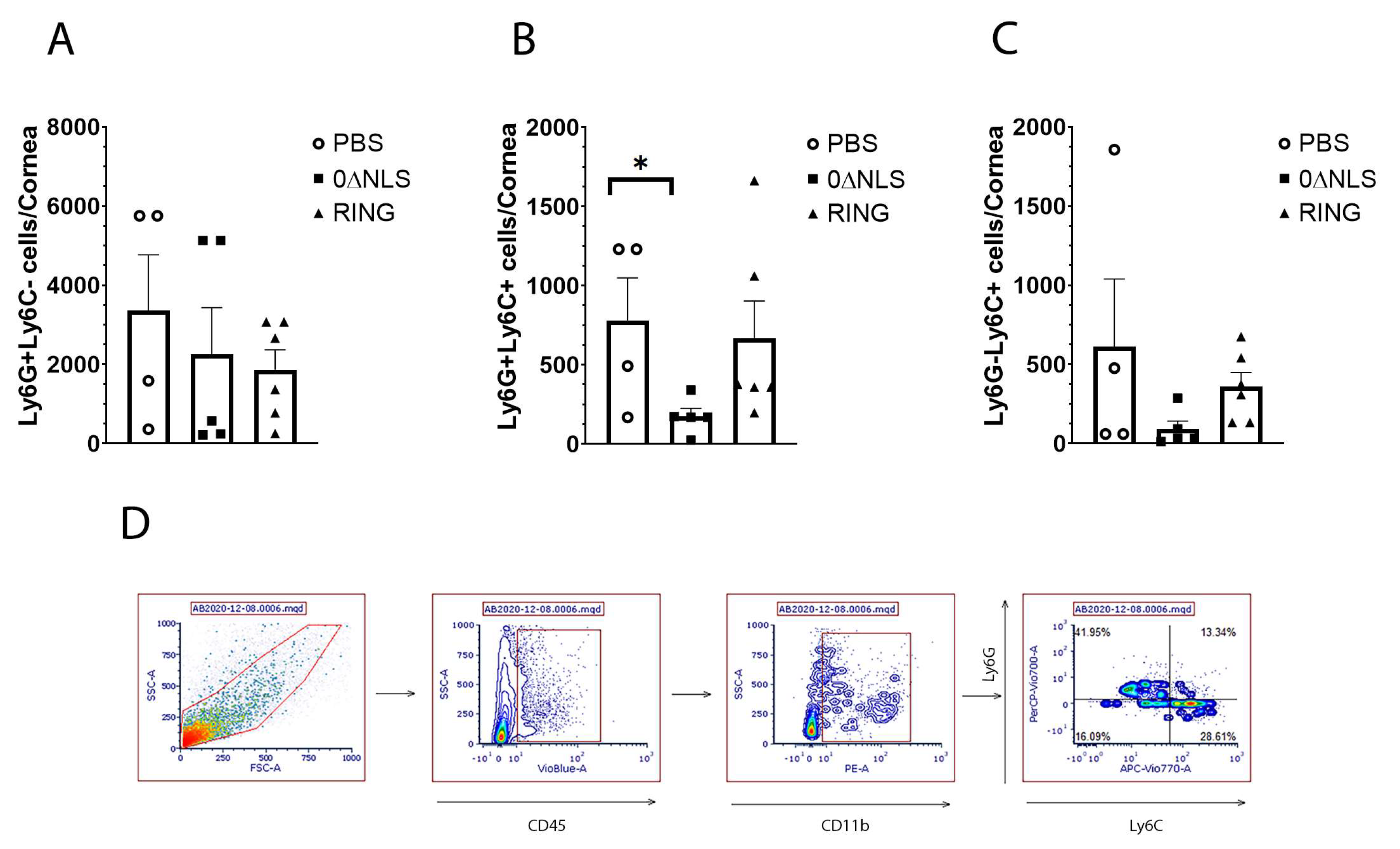
2.5. The Immune Cell Profile of Vehicle- and ICP0 Mutant-Immunized Mice Are Similar following HSV-1 Challenge in Long-Term Study
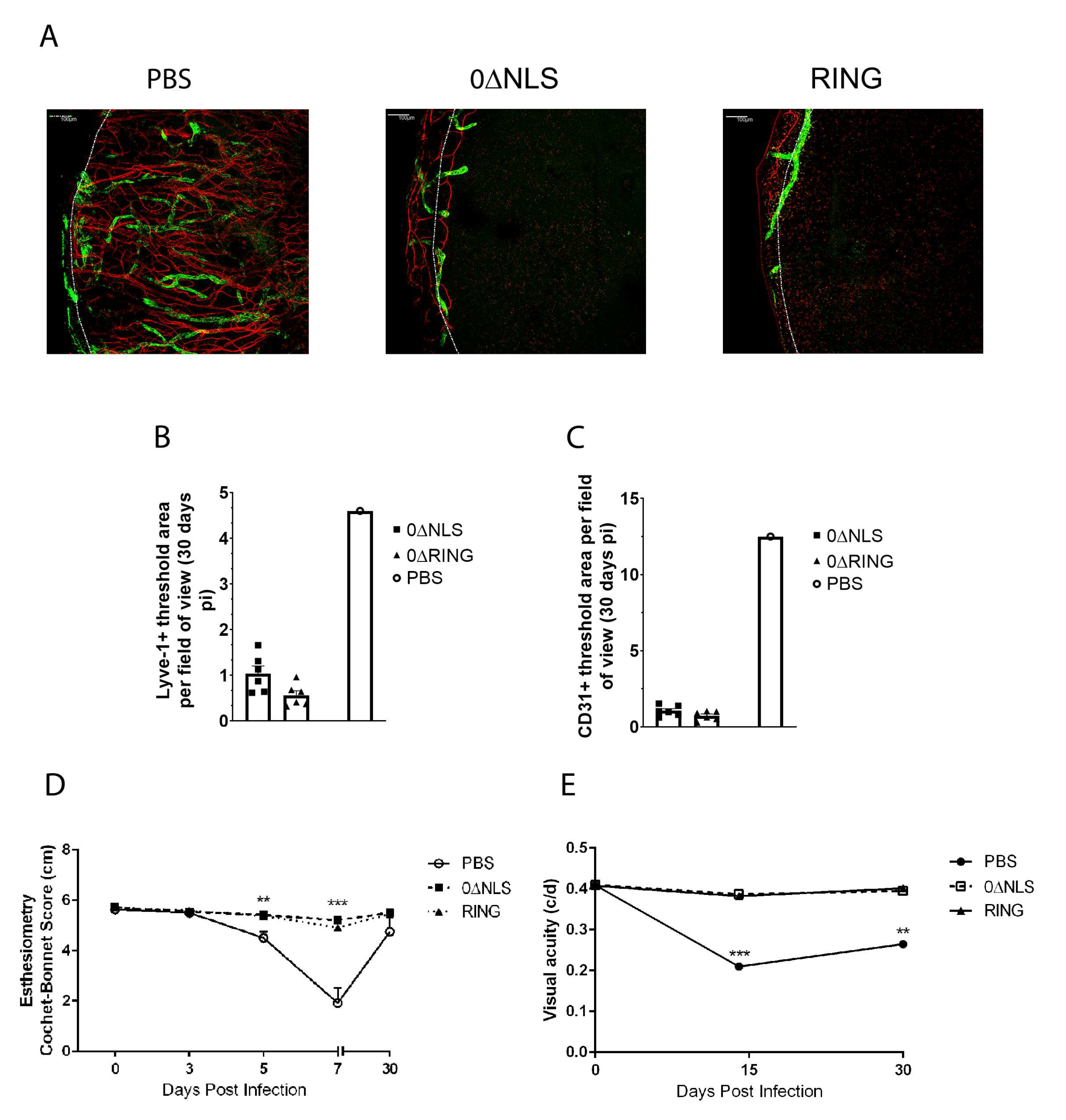
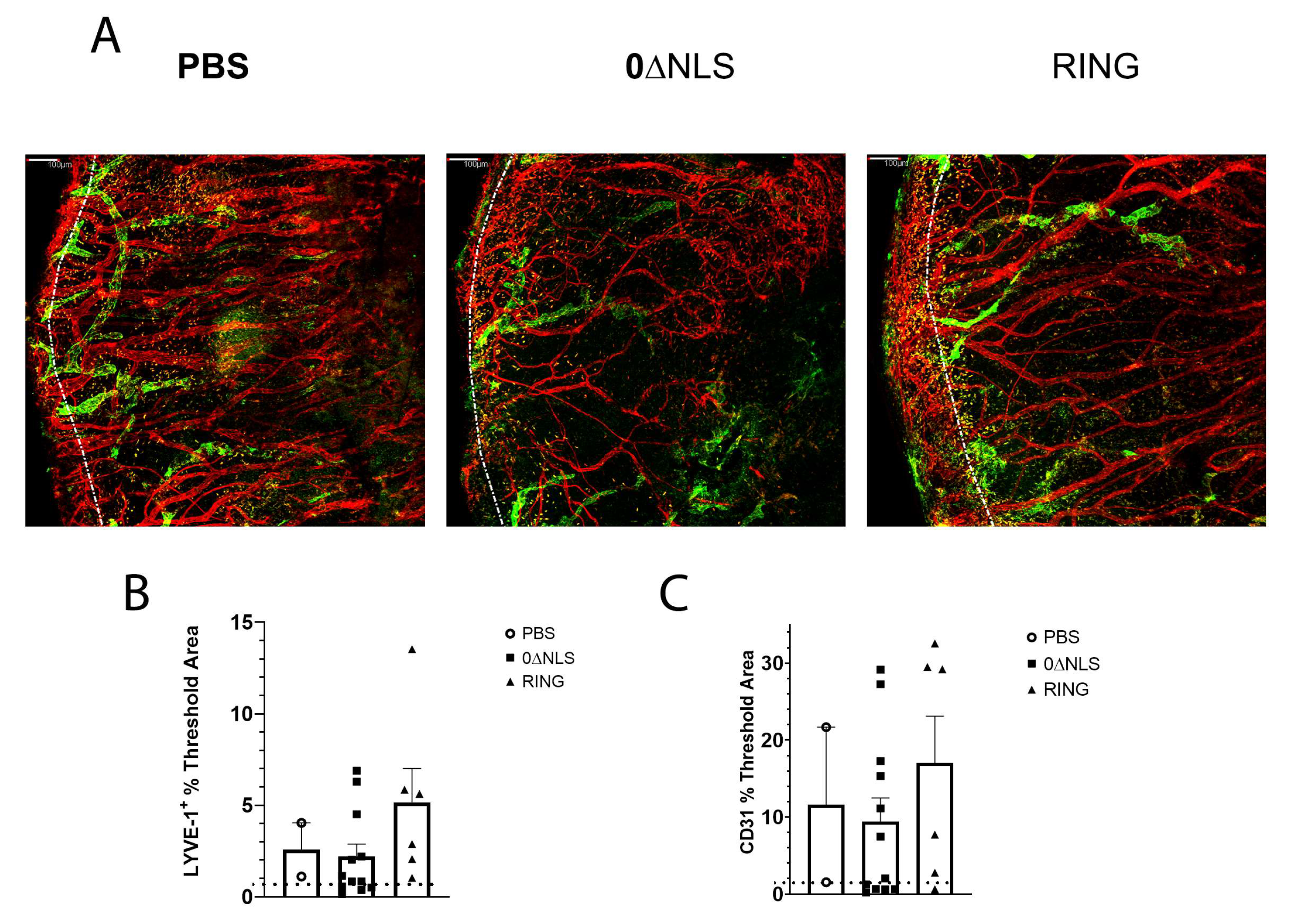
2.6. HSV-1 ICP0 Mutant Immunized Mice Are Not Protected from Virus-Induced Corneal Neovascularization upon Challenge One Year Post-Boost
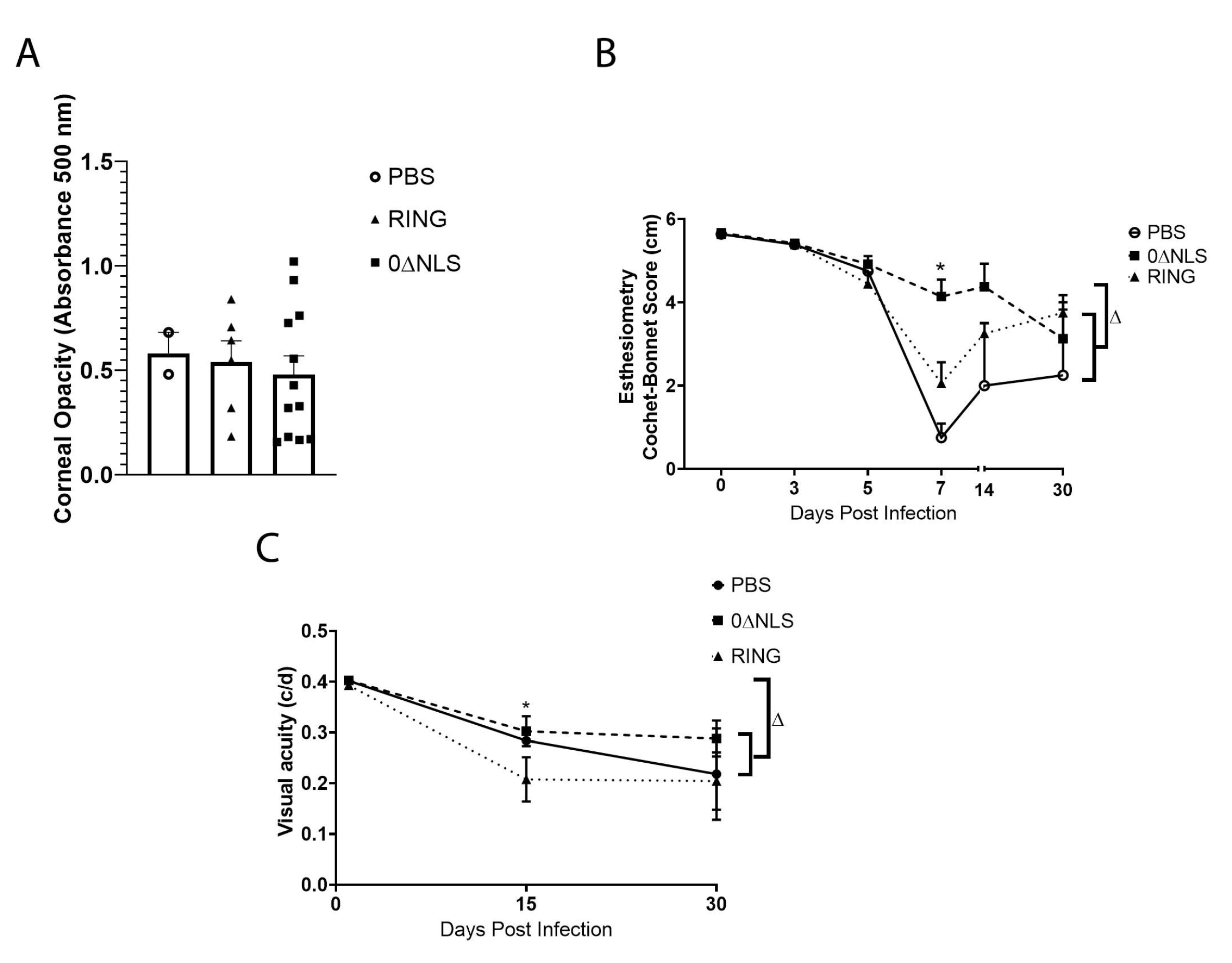
2.7. Corneal Pathology and Function of ICPO Mutant Vaccinated Mice Are Similar to Control Vaccinated Animals in Long-Term Study
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Cells, Virus and Virus Growth Curves
4.3. Immunization and Ocular Infection
4.4. Virus Plaque Assay
4.5. Anti-HSV-1 IgG2b ELISA
4.6. Ocular Pathology
4.7. Assessment of Visual Axis Function
4.8. Flow Cytometry
4.9. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davison, A.J. Herpesvirus systematics. Vet. Microbiol. 2010, 143, 52–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.E.; Vickerman, P.; Gottlieb, S.L.; Newman, L.M. Global and Regional Estimates of Prevalent and Incident Herpes Simplex Virus Type 1 Infections in 2012. PLoS ONE 2015, 10, e0140765. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zheng, C. The Race between Host Antiviral Innate Immunity and the Immune Evasion Strategies of Herpes Simplex Virus 1. Microbiol. Mol. Biol. Rev. 2020, 84, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, H.E.; Azcuy, A.M.; Varnell, E.D.; Sloop, G.D.; Thompson, H.W.; Hill, J.M. HSV-1 DNA in tears and saliva of normal adults. Investig. Ophthalmol. Vis. Sci. 2005, 46, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baringer, J.R.; Pisani, P. Herpes simplex virus genomes in human nervous system tissue analyzed by polymerase chain reaction. Ann. Neurol. 1994, 36, 823–829. [Google Scholar] [CrossRef]
- Singh, N.; Tscharke, D.C. Herpes Simplex Virus Latency Is Noisier the Closer We Look. J. Virol. 2020, 94, e01701-19. [Google Scholar] [CrossRef] [Green Version]
- Dervillez, X.; Qureshi, H.; Chentoufi, A.A.; Khan, A.A.; Kritzer, E.; Yu, D.C.; Diaz, O.R.; Gottimukkala, C.; Kalantari, M.; Villacres, M.C.; et al. Asymptomatic HLA-A*02:01-restricted epitopes from herpes simplex virus glycoprotein B preferentially recall polyfunctional CD8+ T cells from seropositive asymptomatic individuals and protect HLA transgenic mice against ocular herpes. J. Immunol. 2013, 191, 5124–5138. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.; Khan, A.A.; Garg, S.; Syed, S.A.; Furness, J.N.; Vahed, H.; Pham, T.; Yu, H.T.; Nesburn, A.B.; BenMohamed, L. Human Asymptomatic Epitopes Identified from the Herpes Simplex Virus Tegument Protein VP13/14 (UL47) Preferentially Recall Polyfunctional Effector Memory CD44high CD62Llow CD8+ TEM Cells and Protect Humanized HLA-A*02:01 Transgenic Mice against Ocular Herpesvirus Infection. J. Virol. 2017, 91, e01793-16. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.; Khan, A.A.; Spencer, D.; Vahed, H.; Lopes, P.P.; Thai, N.T.U.; Wang, C.; Pham, T.T.; Huang, J.; Scarfone, V.; et al. HLA-A02:01-restricted epitopes identified from the herpes simplex virus tegument protein VP11/12 preferentially recall polyfunctional effector memory CD8+ T cells from seropositive asymptomatic individuals and protect humanized HLA-A*02:01 transgenic mice against ocular herpes. J. Immunol. 2015, 194, 2232–2248. [Google Scholar] [CrossRef] [Green Version]
- Vahed, H.; Agrawal, A.; Srivastava, R.; Prakash, S.; Coulon, P.-G.; Roy, S.; BenMohamed, L. Unique Type I Interferon, Expansion/Survival Cytokines, and JAK/STAT Gene Signatures of Multifunctional Herpes Simplex Virus-Specific Effector Memory CD8(+) TEM Cells Are Associated with Asymptomatic Herpes in Humans. J. Virol. 2019, 93, e01882-18. [Google Scholar] [CrossRef] [Green Version]
- Khanna, K.M.; Bonneau, R.H.; Kinchington, P.R.; Hendricks, R.L. Herpes simplex virus-specific memory CD8+ T cells are selectively activated and retained in latently infected sensory ganglia. Immunity 2003, 18, 593–603. [Google Scholar] [CrossRef] [Green Version]
- Knickelbein, J.E.; Khanna, K.M.; Yee, M.B.; Baty, C.J.; Kinchington, P.R.; Hendricks, R.L. Noncytotoxic lytic granule-mediated CD8+ T cell inhibition of HSV-1 reactivation from neuronal latency. Science 2008, 322, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Slifka, M.K.; Amanna, I. How advances in immunology provide insight into improving vaccine efficacy. Vaccine 2014, 32, 2948–2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, K.J.; Mackett, M.; Wohlenberg, C.; Notkins, A.L.; Moss, B. Vaccinia virus recombinant expressing herpes simplex virus type 1 glycoprotein D prevents latent herpes in mice. Science 1985, 228, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.C.; Kim, Y.S.; Cho, J.K.; Cho, J.; Kim, S.Y.; Kang, H.; Cho, M.H.; Lee, H.H. Enhanced protection against HSV lethal challenges in mice by immunization with a combined HSV-1 glycoprotein B:H:L gene DNAs. Virus Res. 2002, 86, 21–31. [Google Scholar] [CrossRef]
- Keadle, T.L.; Morrison, L.A.; Morris, J.L.; Pepose, J.S.; Stuart, P.M. Therapeutic immunization with a virion host shutoff-defective, replication-incompetent herpes simplex virus type 1 strain limits recurrent herpetic ocular infection. J. Virol. 2002, 76, 3615–3625. [Google Scholar] [CrossRef] [Green Version]
- Keadle, T.L.; Laycock, K.A.; Miller, J.K.; Hook, K.K.; Fenoglio, E.D.; Francotte, M.; Slaoui, M.; Stuart, P.M.; Pepose, J.S. Efficacy of a recombinant glycoprotein D subunit vaccine on the development of primary and recurrent ocular infection with herpes simplex virus type 1 in mice. J. Infect. Dis. 1997, 176, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Dou, J.; Yu, F.; He, X.; Yuan, X.; Wang, Y.; Liu, C.; Gu, N. An ocular mucosal administration of nanoparticles containing DNA vaccine pRSC-gD-IL-21 confers protection against mucosal challenge with herpes simplex virus type 1 in mice. Vaccine 2011, 29, 1455–1462. [Google Scholar] [CrossRef]
- Ramsey, N.L.M.; Visciano, M.; Hunte, R.; Loh, L.N.; Aschner, C.B.; Jacobs, W.R.; Herold, B.C. A Single-Cycle Glycoprotein D Deletion Viral Vaccine Candidate, ΔgD-2, Elicits Polyfunctional Antibodies That Protect against Ocular Herpes Simplex Virus. J. Virol. 2020, 94, e00335-20. [Google Scholar] [CrossRef]
- Naidu, S.K.; Nabi, R.; Cheemarla, N.; Stanfield, B.; Rider, P.J.; Jambunathan, N.; Chouljenko, V.N.; Carter, R.; Del Piero, F.; Langohr, I.; et al. Intramuscular vaccination of mice with the human herpes simplex virus type-1(HSV-1) VC2 vaccine, but not its parental strain HSV-1(F) confers full protection against lethal ocular HSV-1 (McKrae) pathogenesis. PLoS ONE 2020, 15, e0228252. [Google Scholar] [CrossRef]
- Xu, X.; Feng, X.; Wang, L.; Yi, T.; Zheng, L.; Jiang, G.; Fan, S.; Liao, Y.; Feng, M.; Zhang, Y.; et al. A HSV1 mutant leads to an attenuated phenotype and induces immunity with a protective effect. PLoS Pathog. 2020, 16, e1008703. [Google Scholar] [CrossRef] [PubMed]
- Davido, D.J.; Tu, E.M.; Wang, H.; Korom, M.; Casals, A.G.; Reddy, P.J.; Mostafa, H.H.; Combs, B.; Haenchen, S.D.; Morrison, L.A. Attenuated Herpes Simplex Virus 1 (HSV-1) Expressing a Mutant Form of ICP6 Stimulates a Strong Immune Response That Protects Mice against HSV-1-Induced Corneal Disease. J. Virol. 2018, 92, e01036-18. [Google Scholar] [CrossRef] [Green Version]
- Patel, C.D.; Taylor, S.A.; Mehrbach, J.; Awasthi, S.; Friedman, H.M.; Leib, D.A. Trivalent Glycoprotein Subunit Vaccine Prevents Neonatal Herpes Simplex Virus Mortality and Morbidity. J. Virol. 2020, 94, e02163-19. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Nakamura, T.; Yoshida, A.; Takahashi, K.; Inoue, Y.; Shimomura, Y.; Tano, Y.; Fujisawa, Y.; Aono, A.; Hayashi, K. Preventive effect of local plasmid DNA vaccine encoding gD or gD-IL-2 on herpetic keratitis. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4209–4215. [Google Scholar]
- Opstelten, W.; Neven, A.K.; Eekhof, J. Treatment and prevention of herpes labialis. Can. Fam. Physician 2008, 54, 1683–1687. [Google Scholar]
- Royer, D.J.; Gurung, H.R.; Jinkins, J.K.; Geltz, J.J.; Wu, J.L.; Halford, W.P.; Carr, D.J.J. A Highly Efficacious Herpes Simplex Virus 1 Vaccine Blocks Viral Pathogenesis and Prevents Corneal Immunopathology via Humoral Immunity. J. Virol. 2016, 90, 5514–5529. [Google Scholar] [CrossRef] [Green Version]
- Royer, D.J.; Hendrix, J.F.; Larabee, C.M.; Reagan, A.M.; Sjoelund, V.H.; Robertson, D.M.; Carr, D.J.J. Vaccine-induced antibodies target sequestered viral antigens to prevent ocular HSV-1 pathogenesis, preserve vision, and preempt productive neuronal infection. Mucosal Immunol. 2019, 12, 827–839. [Google Scholar] [CrossRef]
- Carr, D.J.J.; Gmyrek, G.B.; Filiberti, A.; Berube, A.N.; Browne, W.P.; Gudgel, B.M.; Sjoelund, V.H. Distinguishing Features of High- and Low-Dose Vaccine against Ocular HSV-1 Infection Correlates with Recognition of Specific HSV-1-Encoded Proteins. Immunohorizons 2020, 4, 608–626. [Google Scholar] [CrossRef]
- Carr, D.J.J.; Berube, A.N.; Filiberti, A.; Gmyrek, G.B. Lack of neonatal Fc receptor does not diminish the efficacy of the HSV-1 0ΔNLS vaccine against ocular HSV-1 challenge. Vaccine 2021, 39, 2526–2536. [Google Scholar] [CrossRef]
- Mossman, K.L.; Saffran, H.A.; Smiley, J.R. Herpes simplex virus ICP0 mutants are hypersensitive to interferon. J. Virol. 2000, 74, 2052–2056. [Google Scholar] [CrossRef] [Green Version]
- Härle, P.; Sainz, B.; Jr Carr, D.J.; Halford, W.P. The immediate-early protein, ICP0, is essential for the resistance of herpes simplex virus to interferon-alpha/beta. Virology 2002, 293, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Lanfranca, M.P.; Mostafa, H.H.; Davido, D.J. HSV-1 ICP0: An E3 Ubiquitin Ligase That Counteracts Host Intrinsic and Innate Immunity. Cells 2014, 3, 438–454. [Google Scholar] [CrossRef] [PubMed]
- Shahnazaryan, D.; Khalil, R.; Wynne, C.; Jefferies, C.A.J.; Murphy, C.C. Herpes simplex virus 1 targets IRF7 via ICP0 to limit type I IFN induction. Sci. Rep. 2020, 10, 22216. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Schmidt, E.E.; Halford, W.P. ICP0 dismantles microtubule networks in herpes simplex virus-infected cells. PLoS ONE 2010, 5, e10975. [Google Scholar] [CrossRef] [PubMed]
- Halford, W.P.; Schaffer, P.A. Optimized viral dose and transient immunosuppression enable herpes simplex virus ICP0-null mutants To establish wild-type levels of latency in vivo. J. Virol. 2000, 74, 5957–5967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halford, W.P.; Püschel, R.; Rakowski, B. Herpes simplex virus 2 ICP0 mutant viruses are avirulent and immunogenic: Implications for a genital herpes vaccine. PLoS ONE 2010, 5, e12251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royer, D.J.; Carr, M.M.; Chucair-Elliott, A.J.; Halford, W.P.; Carr, D.J. Impact of Type I Interferon on the Safety and Immunogenicity of an Experimental Live-Attenuated Herpes Simplex Virus 1 Vaccine in Mice. J. Virol. 2017, 91, e02342-16. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.; Nikolich-Zugich, J. Development and migration of protective CD8+ T cells into the nervous system following ocular herpes simplex virus-1 infection. J. Immunol. 2005, 174, 2919–2925. [Google Scholar] [CrossRef] [Green Version]
- Gmyrek, G.B.; Filiberti, A.; Montgomery, M.; Chitrakar, A.; Royer, D.J.; Carr, D.J.J. Herpes Simplex Virus 1 (HSV-1) 0ΔNLS Live-Attenuated Vaccine Protects against Ocular HSV-1 Infection in the Absence of Neutralizing Antibody in HSV-1 gB T Cell Receptor-Specific Transgenic Mice. J. Virol. 2020, 94, e01000-20. [Google Scholar] [CrossRef]
- Zheng, M.; Deshpande, S.; Lee, S.; Ferrara, N.; Rouse, B.T. Contribution of vascular endothelial growth factor in the neovascularization process during the pathogenesis of herpetic stromal keratitis. J. Virol. 2001, 75, 9828–9835. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, F.; Suryawanshi, A.; Rouse, B.T. Pathogenesis of herpes stromal keratitis--a focus on corneal neovascularization. Prog. Retin. Eye Res. 2013, 33, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuest, T.R.; Carr, D.J. VEGF-A expression by HSV-1-infected cells drives corneal lymphangiogenesis. J. Exp. Med. 2010, 207, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wuest, T.; Zheng, M.; Efstathiou, S.; Halford, W.P.; Carr, D.J. The herpes simplex virus-1 transactivator infected cell protein-4 drives VEGF-A dependent neovascularization. PLoS Pathog. 2011, 7, e1002278. [Google Scholar] [CrossRef] [Green Version]
- Yun, H.; Rowe, A.M.; Lathrop, K.L.; Harvey, S.A.; Hendricks, R.L. Reversible nerve damage and corneal pathology in murine herpes simplex stromal keratitis. J. Virol. 2014, 88, 7870–7880. [Google Scholar] [CrossRef] [Green Version]
- Chucair-Elliott, A.J.; Zheng, M.; Carr, D.J. Degeneration and regeneration of corneal nerves in response to HSV-1 infection. Investig. Ophthalmol. Vis. Sci. 2015, 56, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Rowe, A.M.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef]
- Douglas, R.M.; Alam, N.M.; Silver, B.D.; McGill, T.J.; Tschetter, W.W.; Prusky, G.T. Independent visual threshold measurements in the two eyes of freely moving rats and mice using a virtual-reality optokinetic system. Vis. Neurosci. 2005, 22, 677–684. [Google Scholar] [CrossRef]
- Gurung, H.R.; Carr, M.M.; Bryant, K.; Chucair-Elliott, A.J.; Carr, D.J. Fibroblast growth factor-2 drives and maintains progressive corneal neovascularization following HSV-1 infection. Mucosal Immunol. 2018, 11, 172–185. [Google Scholar] [CrossRef] [Green Version]
- Halford, W.P.; Weisend, C.; Grace, J.; Soboleski, M.; Carr, D.J.J.; Balliet, J.W.; Imai, Y.; Margolis, T.P.; Gebhardt, B.M. ICP0 antagonizes Stat 1-dependent repression of herpes simplex virus: Implications for the regulation of viral latency. Virol. J. 2006, 3, 44. [Google Scholar] [CrossRef] [Green Version]
- Davis, W.B.; Taylor, J.A.; Oakes, J.E. Ocular infection with herpes simplex virus type 1: Prevention of acute herpetic encephalitis by systemic administration of virus-specific antibody. J. Infect. Dis. 1979, 140, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Conrady, C.D.; Zheng, M.; Van Rooijen, N.; Drevets, D.A.; Royer, D.; Alleman, A.; Carr, D.J.J. Microglia and a functional type I IFN pathway are required to counter HSV-1-driven brain lateral ventricle enlargement and encephalitis. J. Immunol. 2013, 190, 2807–2817. [Google Scholar] [CrossRef] [PubMed]
- Amanna, I.J.; Slifka, M.K. Mechanisms that determine plasma cell lifespan and the duration of humoral immunity. Immunol. Rev. 2010, 236, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Royer, D.J.; Carr, M.M.; Gurung, H.R.; Halford, W.P.; Carr, D.J.J. The Neonatal Fc Receptor and Complement Fixation Facilitate Prophylactic Vaccine-Mediated Humoral Protection against Viral Infection in the Ocular Mucosa. J. Immunol. 2017, 199, 1898–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lal, H.; Cunningham, A.L.; Godeaux, O.; Chlibek, R.; Diez-Domingo, J.; Hwang, S.-J.; Levin, M.J.; McElhaney, J.E.; Poder, A.; Puig-Barberà, J.; et al. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N. Engl. J. Med. 2015, 372, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samaniego, L.A.; Neiderhiser, L.; DeLuca, N.A. Persistence and expression of the herpes simplex virus genome in the absence of immediate-early proteins. J. Virol. 1998, 72, 3307–3320. [Google Scholar] [CrossRef] [Green Version]
- Halford, W.P.; Gebhardt, B.M.; Carr, D.J. Acyclovir blocks cytokine gene expression in trigeminal ganglia latently infected with herpes simplex virus type 1. Virology 1997, 238, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Filiberti, A.; Gmyrek, G.B.; Montgomery, M.L.; Sallack, R.; Carr, D.J.J. Loss of Osteopontin Expression Reduces HSV-1-Induced Corneal Opacity. Investig. Ophthalmol. Vis. Sci. 2020, 61, 24. [Google Scholar] [CrossRef]
- Larabee, C.; Hu, Y.; Desai, S.; Georgescu, C.; Wren, J.; Axtell, R.C.; Plafker, S.M. Myelin-specific Th17 cells induce severe relapsing optic neuritis with irreversible loss of retinal ganglion cells in C57BL/6 mice. Mol. Vis. 2016, 22, 332–341. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carr, D.J.J.; Berube, A.; Gershburg, E. The Durability of Vaccine Efficacy against Ocular HSV-1 Infection Using ICP0 Mutants 0∆NLS and 0∆RING Is Lost over Time. Pathogens 2021, 10, 1470. https://doi.org/10.3390/pathogens10111470
Carr DJJ, Berube A, Gershburg E. The Durability of Vaccine Efficacy against Ocular HSV-1 Infection Using ICP0 Mutants 0∆NLS and 0∆RING Is Lost over Time. Pathogens. 2021; 10(11):1470. https://doi.org/10.3390/pathogens10111470
Chicago/Turabian StyleCarr, Daniel J. J., Amanda Berube, and Edward Gershburg. 2021. "The Durability of Vaccine Efficacy against Ocular HSV-1 Infection Using ICP0 Mutants 0∆NLS and 0∆RING Is Lost over Time" Pathogens 10, no. 11: 1470. https://doi.org/10.3390/pathogens10111470
APA StyleCarr, D. J. J., Berube, A., & Gershburg, E. (2021). The Durability of Vaccine Efficacy against Ocular HSV-1 Infection Using ICP0 Mutants 0∆NLS and 0∆RING Is Lost over Time. Pathogens, 10(11), 1470. https://doi.org/10.3390/pathogens10111470