1. Introduction
The genus
Salmonella includes two species:
Salmonella bongori and
Salmonella enterica. The latter includes six subspecies:
S. enterica subsp.
enterica (I),
S. enterica subsp.
salamae (II),
S. enterica subsp.
arizonae (IIIa),
S. enterica subsp.
diarizonae (IIIb),
S. enterica subsp.
houtenae (IV) and
S. enterica subsp.
indica (V). Isolates currently classified in
S. enterica subsp.
diarizonae were first detected in faecal samples from reptiles [
1] and initially included into the
Salmonella “Arizona” group, later termed subgenus Arizona or subgenus III. The subgenus was subsequently divided into
S. enterica subsp.
arizonae (IIIa) and
S. enterica subsp.
diarizonae (IIIb), based on differing reactions in biochemical tests and genomic relatedness [
2].
At least 336 distinct serovars of
S. enterica subsp.
diarizonae have been detected, which was approximately 13% of all recorded serovars in the
S. enterica species [
3,
4].
S. enterica subsp.
diarizonae isolates have been most frequently recovered from samples collected from cold-blooded animals [
5,
6] or the environment and may also be harboured by domestic (e.g., sheep, details below) or wild [
7,
8] warm-blooded animals, as well as humans.
S. enterica subsp.
diarizonae mainly colonises the gastrointestinal tract of hosts, specifically the anterior part of the small intestine.
S. enterica subsp.
diarizonae is considered as the most frequently detected
Salmonella subspecies in sheep, but it has not been studied widely in the literature. Most
S. enterica subsp.
diarizonae infections are caused by isolates with antigenic type 61:k:1,5,(7) [
9], although serovars with minor modifications or with incomplete antigenic structure have also been detected.
The organism was isolated from samples from sheep in the United Kingdom [
10], Norway [
11,
12] and Germany [
13], with isolation rates varying from 1% to 76% of samples. Further, during abattoir studies in the United Kingdom [
14], Switzerland [
15] and Sweden [
16], isolation rates were always less than 1% of sheep carcasses.
The organism is a cause of clinical diseases in sheep [
17]. It has been associated mainly with gastrointestinal disorders in lambs [
18,
19,
20], abortion in ewes [
21] and chronic proliferative rhinitis in adult sheep [
22,
23].
Recovery of the organism in samples from lambs with diarrhoea can potentially lead to diagnostic problems regarding the causal agent(s) of the problem. We have already described an outbreak of diarrhoeic syndrome in a flock of sheep, in which
S. enterica subsp.
diarizonae was isolated from faecal samples from a lamb with clinical signs, as well as from samples from a clinically healthy ewe in the same farm [
20]. Despite previous relevant studies, the possible role of the pathogen in the aetiology of the diarrhoeic syndrome in lambs has not been fully clarified.
The objectives of this experimental study were the evaluation of the potential role of S. enterica subsp. diarizonae in the diarrhoeic syndrome in lambs and the investigation of facets of the pathogenesis of the infection.
3. Discussion
After challenge, intestinal infection has been established in inoculated lambs. This was corroborated by the consistent isolation of the challenge organism from the lambs and the definite evidence of inflammation during the histopathological evaluation. Nevertheless, infection was mild and did not lead to fatalities as recorded with other
Salmonella species [
25]. The organism can nevertheless be confirmed as an intestinal pathogen. The recovery of an isolate from a field case of intestinal infection in a lamb, the subsequent use of this isolate in the experimental reproduction of a mild-type intestinal infection and the re-isolation of the organism from the experimental animals confirm the causality of the disease. The mild effects on the experimental animals should not deter from confirming this association, but rather they are indicative of a mild pathogenicity of the bacterium and effective defences of the animals.
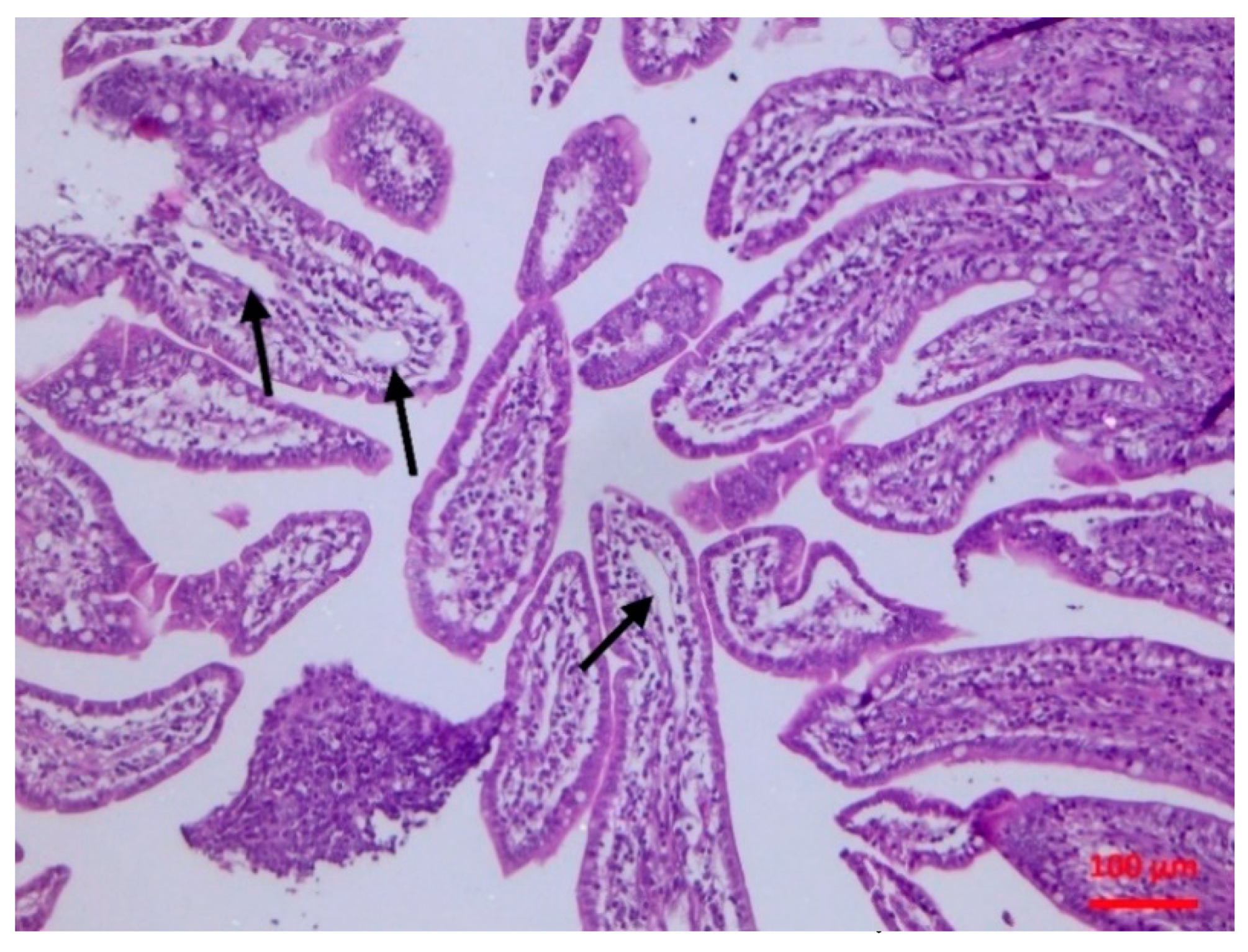
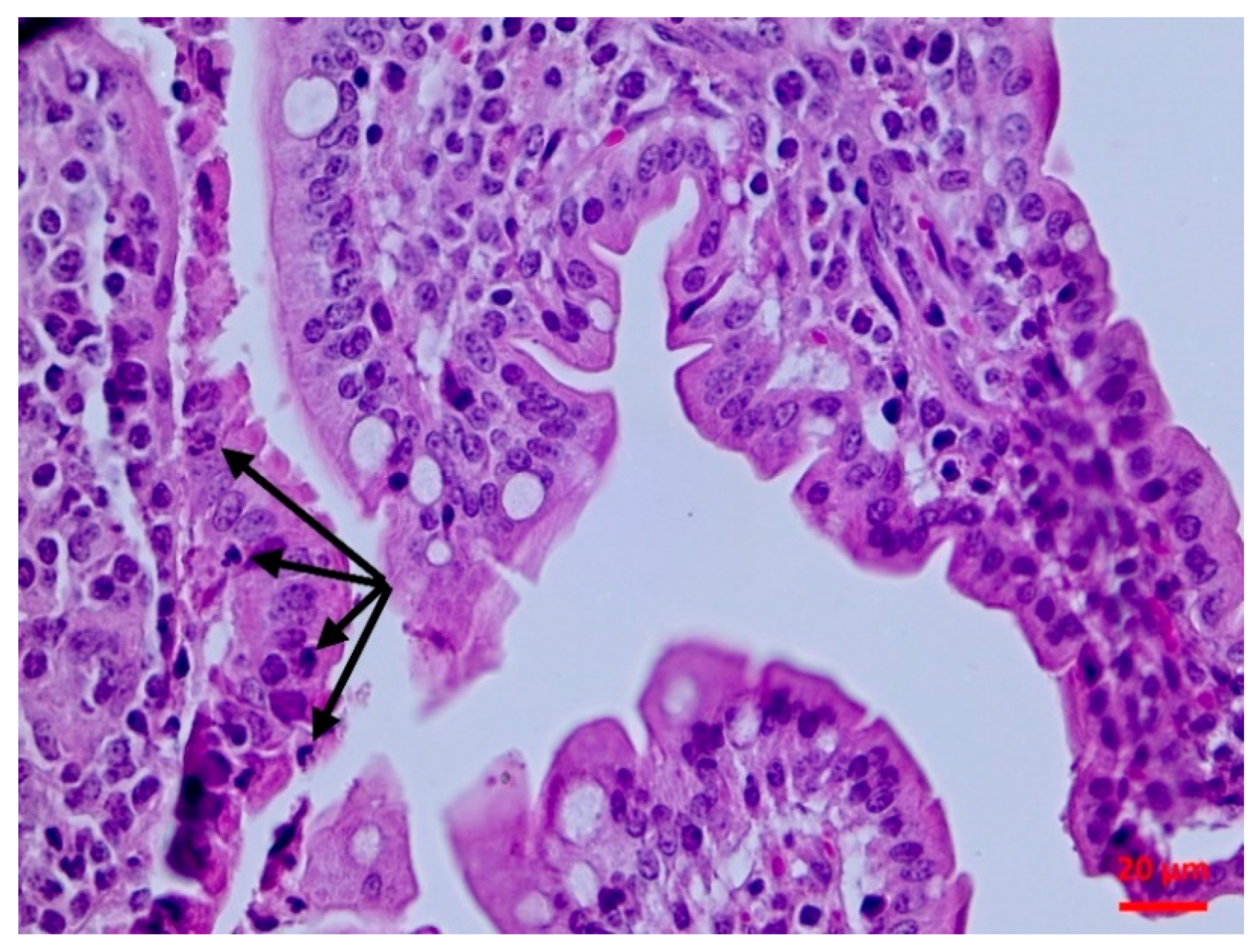
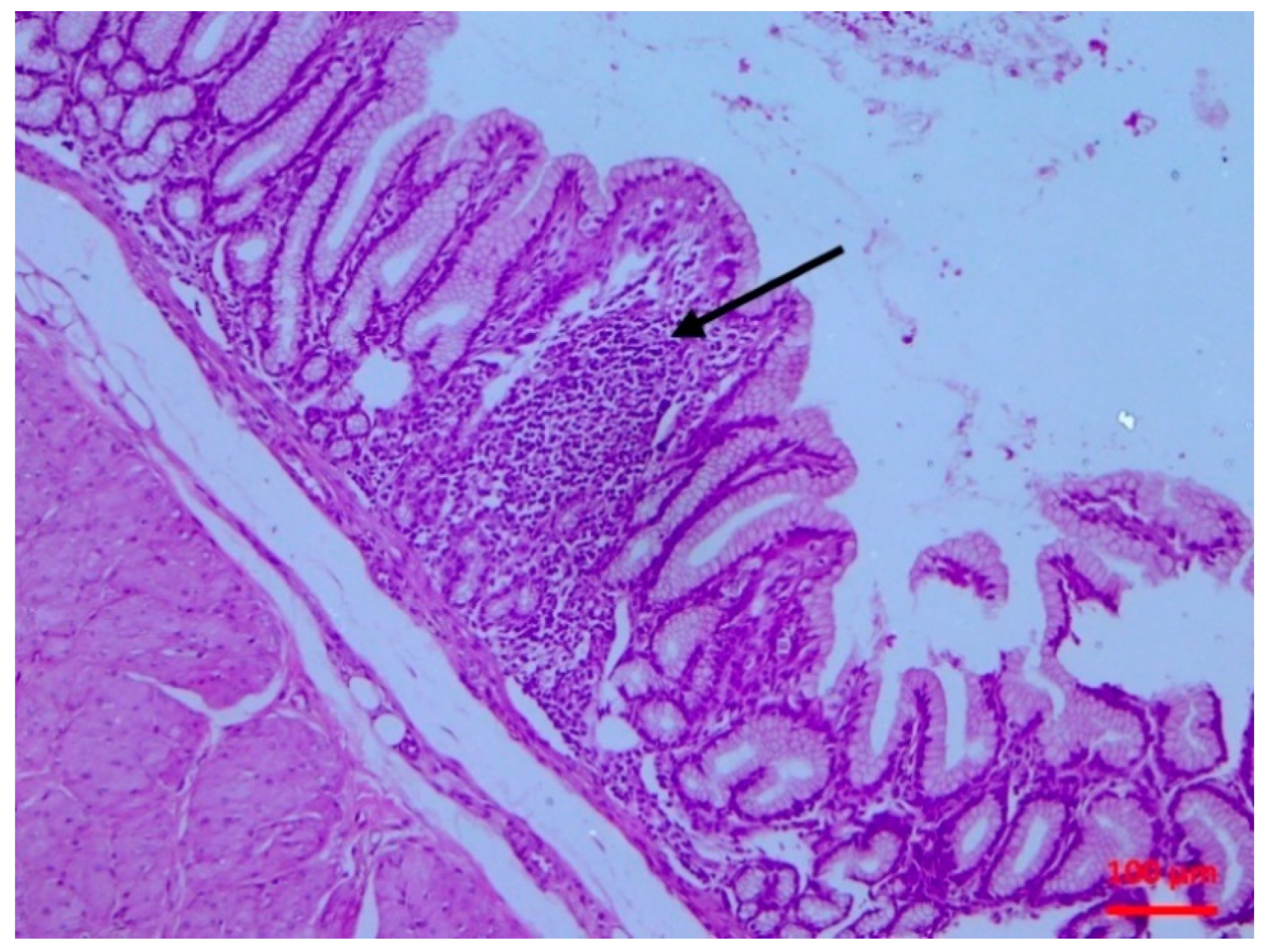
The isolate caused subclinical damage which was evident soon after inoculation. There were clear pathological findings in the intestinal mucosa, as confirmed by the histopathological examination. A possible destruction of the villi (even of mild extent) can lead to problems of nutrient absorption, whilst the observed lymphofollicular atrophy may predispose to reduced intestinal defences. Whilst
S. enterica subsp.
diarizonae may be of low pathogenic significance on its own, it may pave a way for other bacteria to exert their pathogenicity in infected lambs, causing more significant gastrointestinal problems, which are of paramount importance in lambs [
25].
At the end, the challenge isolate had disseminated outside the gastrointestinal tract and was isolated from liver and gallbladder tissue samples. The findings are in contrast to a hypothesis by Katribe et al. [
26], who indicated that
S. enterica subsp.
diarizonae was limited in the intestinal tract. Many researchers believe that this specific subspecies may be a commensal resident of sheep’s intestinal tract [
17]. However, our findings are allied more to the results of Lacasta et al. [
27], who have also reported extra-intestinal (respiratory) infection of sheep with the pathogen. This shows the possibility for invasiveness of the organism; hence, in immunocompromised hosts, it might even be able to cause mortality. The results suggest that, whilst
S. enterica subsp.
diarizonae seems to be a host-adapted
Salmonella subspecies, the organism can retain its pathogenic properties and, under certain conditions, may cause clinical conditions in sheep.
The isolation of the organism from buccal samples is of particular interest. Bacteria in the mouth of lambs could have originated from the inoculum or from regurgitation of gastric content; the latter possibility is more likely, as a gastric catheter was employed for inoculation, although the possibility of a leakage during the inoculation process cannot be ruled out. Consequentially, the isolation of
Salmonella from milk samples of a ewe is consistent with the presence of the organism within the mouth of her offspring, whence it was likely transferred during sucking by lambs. We postulate that as the lower part of the teat comes into contact with the pharynx of the lamb [
28], the organism was attached thereon, subsequently entering into the duct; perhaps the tongue of the lamb might have “pushed” the bacteria upwards into the duct. In previous studies [
29], we have presented evidence that bacteria can be transferred from the mouth of lambs to the teat duct of their dams even after a short (1 min) sucking activity.
In a field study previously reported [
20],
S. enterica subsp.
diarizonae was isolated from faecal samples of a lamb with grave clinical signs, as well as from faecal samples of a ewe. The present findings are in contrast to those of the field study. These contrasting findings can be attributed to a possibly reduced immune state of those animals, given that the field work was undertaken in a sheep flock a while earlier affected with bluetongue, which may cause immunosuppression in affected animals [
30,
31]. The finding of less frequent recoveries of the organism by microbiological techniques than the frequency of detection of nucleic acid by molecular techniques lends some support to this hypothesis. Likely, in the experimental study, effective defences of the host eliminated the challenged organism, despite the high dose administered. This is further supportive of a mild pathogenicity of the organism, as postulated above.
Detection of the
invA gene was used to confirm the presence of
Salmonella using conventional PCR. Although this gene is not specific in
S. enterica subsp.
diarizonae, it was selected because of its increased sensitivity for
Salmonella detection [
32]. Each sample that yielded the expected PCR product was interpreted to have harboured
S. enterica subsp.
diarizonae. The simultaneous presence of
Salmonella strains beyond the challenge organism was considered unlikely to have occurred, especially with the findings of the PCR allied to the bacteriological results. In particular, the following points were taken into account: (a) no
Salmonella was recovered from the animals before challenge, nor was the
invA gene detected in any sample before challenge, (b) the experimental animals remained isolated from each other throughout the study, with limited and strictly controlled access to their pens, (c)
Salmonella recovered from the samples after challenge was speciated as
S. enterica subsp.
diarizonae, (d) typing of the isolates from the experimental animals after challenge in all cases confirmed the identity of the isolates as
S. enterica subsp.
diarizonae.
The infection in sheep poses a zoonotic threat. Lamb consumption may contribute to potential human infections, as shown by the isolation of the organism from abattoir samples at a small but existent rate [
14,
15,
16]. Moreover, the recovery of this organism from milk samples (present results) should increase awareness about the presence of previously unrecognised pathogens in the milk of ewes, which may subsequently be transferred to humans in cases of inappropriate thermal processing of milk. The detection of multi-resistant [
33] and of colistin-resistant [
15] isolates of the organism from animals should also be taken into account, when assessing the significance of the potential zoonotic risk of the organism.
4. Materials and Methods
4.1. Animals
In total, 16 clinically healthy lambs from 8 ewes were enrolled into the study on the first day of life. Of these, 12 lambs were challenged with S. enterica subsp. diarizonae serovar 61:k:1,5,(7). The dams of the lambs had been housed throughout their gestation and were provided with a commercial concentrate feed plus hay and barlay straw. During the final month of gestation and after lambing, ewes were housed individually; during the latter period, lambs were also penned with their respective dam.
Two examinations of blood samples for concentrations of β-hydroxybutyrate [
34] did not reveal any problems: in all animals, concentrations were always below 0.95 mmol L
−1. Further, examination of serum blood samples, by using ELISA tests with commercially available kits, for presence of antibodies against
Mycobacterium avium subsp.
paratuberculosis (ID Screen
® Paratuberculosis Indirect; ID VET, Grabels, France),
Small Ruminant Lentivirus (ID Screen
® MVV / CAEV Indirect; ID VET) and
Bluetongue Virus (ID Screen
® Bluetongue Competition, ID VET) [
35,
36,
37], also did not reveal any problems: in no ewe, antibodies of the above pathogens were detected by any of these tests. Finally, bacteriological examination of faecal samples of the ewes performed during the final week of gestation performed by standard techniques (including the ISO 6579-1:2017 for detection of
Salmonella [
38]), did not reveal a
Salmonella infection in these animals.
The experiment was performed under a licence issued by the Veterinary Authority of the Region of Thessaly (licence no. 1997/30.01.2019), which was the competent authority to allow and monitor the experimentation. Conditions prescribed by legislation of the European Union in relation to animal experimentation procedures (Council Directive 86/809/EEC) were met during this work.
4.2. Inoculation Procedure
The lambs were challenged with a
S. enterica subsp.
diarizonae serovar 61:k:1,5,(7) that had been isolated during the investigation of the outbreak of diarrhoeic syndrome in a sheep farm [
20]. Inoculation of lambs was performed on the 1st day of life (D0).
For inoculation, the challenge isolate was cultured in brain heart infusion broth (Thermo Fisher Scientific-Oxoid, Waltham, MA, USA) for 12 h at 37 °C. The culture was centrifuged and the sediment diluted into 20 mL phosphate-buffer saline pH 7.3 (PBS). A quantity of 10 mL of PBS was aspirated with a sterile syringe and, through use of a sterile plastic gastric catheter, was slowly introduced into the abomasum of the newborn lambs, by following the standard principles of administration of oral solutions to lambs [
26]. Lambs were maintained at that position for 3 min after end of the procedure. The inoculum varied from 0.75 × 10
9 to 1.80 × 10
9 colony-forming units, as estimated by the method of Miles and Misra [
39].
For this method, 1 mL of the inoculum was diluted into 9 mL of sterile phosphate-buffer-saline (PBS) and then serial dilutions of the suspension were performed in sterile PBS (1 mL into 9 mL). A drop of 0.02 mL of each serial dilution was plated and spread onto a plate with Plate Count Agar (PCA) (Thermo Fisher Scientific-Oxoid) and allowed to stand for 20 min before aerobic incubation at 37 °C for 18 to 24 h. The dilutions were performed in triplicate and for each dilution, three plates were inoculated. Then, colony counts on plates were made in drop areas with 20 to 100 colonies. The results of the three plates from each series of dilutions were averaged; then, the three means were again averaged for the final result of the content of the inoculum.
Four lambs received 10 mL of sterile PBS by using the above technique and were used as uninfected controls.
4.3. Examination of Animals-Samplings
4.3.1. Lambs
On D0, but before challenge, a detailed clinical examination was carried out in lambs. Blood samples were collected from the jugular vein for haematological examination. Faecal and buccal swab samples were collected for bacteriological examination for detection of Salmonella spp.
Post-challenge, clinical examinations and sample collection as above were performed at 6 h (D0 + 6 h), 12 h (D0 + 12 h), 1 day (D1) and 2 (D2), 4 (D4), 7 (D7), 10 (D10), 14 (D14) and 21 (D21) days. On D1 and thereafter, lambs were euthanised (n = 1 on each of D1 and D2, n = 2 on each of D4, D7, D10, D14, D21). The uninfected controls were euthanised on D4 (n = 1), D10 (n = 1) and D21 (n = 2).
A detailed post-mortem examination was performed in all euthanised lambs. Tissue samples from the abomasum, the small intestine, the liver and the gallbladder were collected for bacteriological examination by using standard techniques and for detection of Salmonella spp. DNA by using a conventional PCR assay. Further, tissue samples from the abomasum, the small intestine and the mesenteric lymph nodes were collected for histopathological examination.
4.3.2. Ewes
On the same occasions as above, faecal swab and milk samples were collected from the ewes for bacteriological examination. Milk samples were collected aseptically, separately from each of the two mammary glands of each ewe [
40].
4.4. Laboratory Examinations
4.4.1. Haematological Examination
Samples for haematological examination were mixed by gentle repeated inversions for several seconds to avoid coagulation. They were processed within 30 min after collection. Initially, blood smears were prepared and kept dry at room temperature. A complete blood count was performed by an automated haematological analyser (Abbott Cell-Dyn 3500 System; Abbott, Abbott Park, IL, USA) previously evaluated in ovine haematology [
41]. The following parameters were determined: haematocrit, erythrocyte count, haemoglobin concentration, mean corpuscular volume, mean corpuscular haemoglobin concentration, total leucocyte count and thrombocyte count. Blood smears were evaluated for leucocyte type differentiation and detection of potential presence of morphological abnormalities.
4.4.2. Bacteriological Examination
Faecal swab samples from lambs and ewes were processed for isolation of Salmonella, after immersion into 1 mL of buffered peptone water. Buccal swab samples from lambs were processed similarly. For milk samples from ewes, a volume of 1 mL of milk was mixed with 10 mL of buffered peptone water.
Tissue samples from lambs collected during post-mortem examination were washed with PBS and were then homogenised (10 g of tissue sample with 50 mL of sterile PBS blended for 3 min) in a tissue blender (Mixwel; Alliance Bio Expertise, Guipry, France). Then, of the resulting fluid, 20 mL were added into 200 mL of buffered peptone water, which was followed by the same procedure as above.
After initial processing as above, the enriched suspensions were processed according to the ISO 6579-1:2017 for detection of
Salmonella [
38]; modified semi-solid Rappaport Vassiliadis medium (Thermo Scientific-Oxoid), XLD (Thermo Scientific-Oxoid) and Salmonella-Shigella agar (Thermo Scientific-Oxoid) agar were used. Colonies obtained were cultured onto sheep blood agar plates and McConkey plates for incubation at 37 °C for up to 48 h. Colonies grown were processed for identification by using the API rapid identification system (Biomerieux, Marcy-l’-Etoile, France). Then, the automated identification Vitek 2 system (Biomerieux) was employed for confirmation of identification of all
Salmonella isolates obtained as above. Finally, eight isolates selected at random among the isolates recovered from all clinical samples (3 from faecal, 3 from buccal and 2 from milk samples), were subcultured and sent to the Greek National Reference Laboratory for Salmonellae (in animals), which is a service of the Greek Ministry of Rural Development and Food, for serotyping.
4.4.3. Molecular Examination for Presence of Salmonella spp.
PCR Amplification
Presence of
Salmonella spp.
invA gene was detected by simple PCR assay. Details of primers and conditions employed are shown in
Table 8. Amplification was performed in a PT-100 Thermocycler (MJ Research Inc., St Bruno, QC, Canada). Reactions were performed in a total volume of 50 μL PCR mixture, containing 0.045 mL of Platinum PCR SuperMix (Applied Biosystems, Foster City, CA, USA) and approximately 150 ng of the extracted DNA. The thermal cycling procedure consisted of a pre-denaturation step at 95 °C for 2 min, 35 cycles of denaturation at 94 °C for 1 min, annealing for 1 min and extending at 72 °C for 45 s and a final elongation step at 72 °C for 7 min. Subsequently, 0.005 mL of each product was analysed by electrophoresis on 1.5% agarose gel stained with ethidium bromide (100 mL 1× TBE buffer; DGel Electrosystem, Montreal, QC, Canada), 2 g agarose (NIPPON Genetics, Tokyo, Japan), 0.005 mL ethidium bromide (Sigma-Aldrich, Saint Louis, MO, USA) and observed at ultraviolet light. Each product equal in size to the expected amplicon was considered as positive.
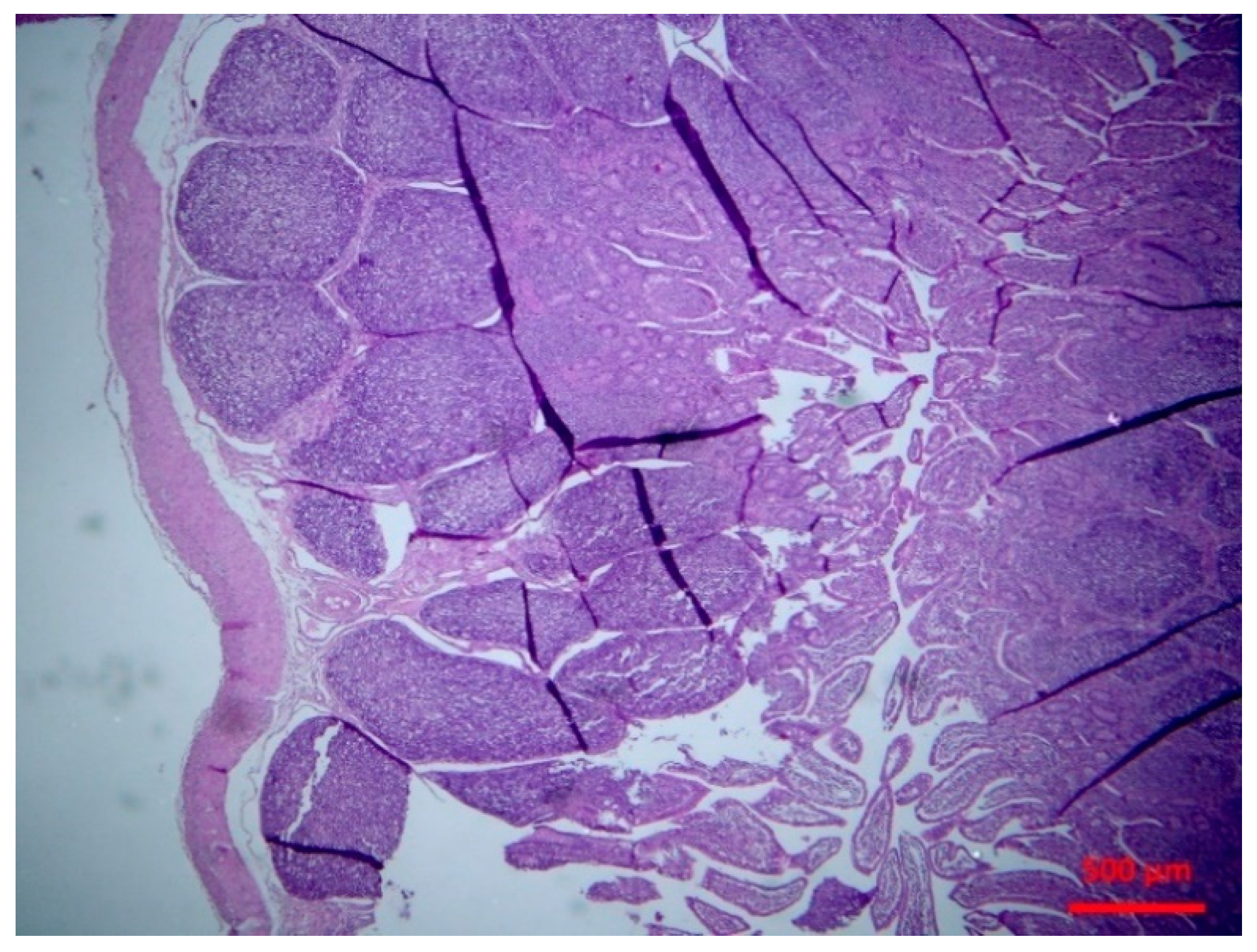
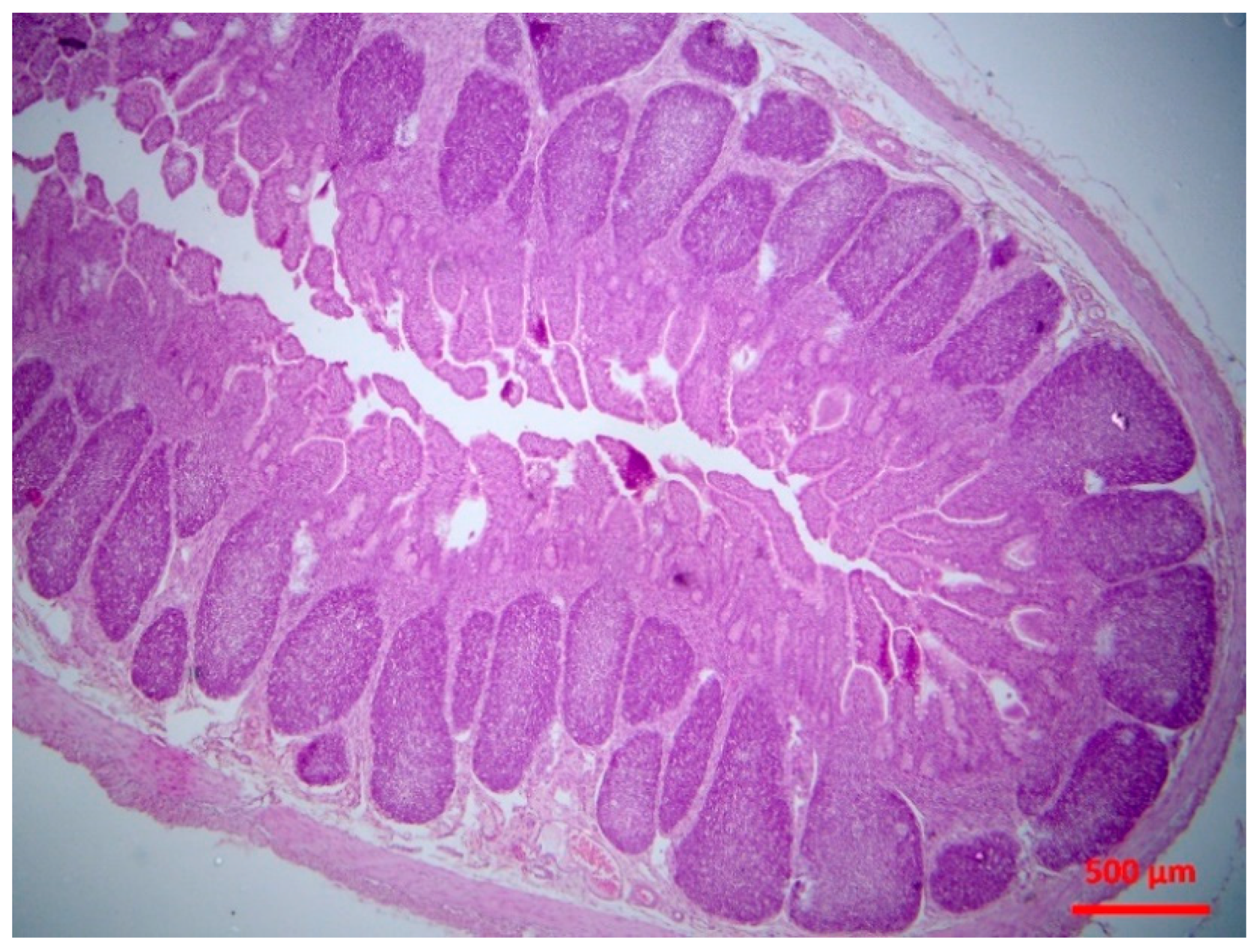
4.4.4. Histopathological Examination
Tissue samples were fixed in 10% neutral-buffered formalin and embedded in paraffin wax. Haematoxylin and eosin (H&E) standard staining procedures were performed for histopathological studies.
4.5. Data Management and Analysis
All data were entered into Microsoft Excel and analysed using IBM SPSS Statistics (ver. 21) (IBM; Armonk, NY, USA).
For estimation of incidence rates, we took into account that a lamb might change from not being infected to being infected and vice-versa; during the interval between sampling points, it was not possible to know what had happened between the two sampling points, i.e., how many cases of infection and “cures” might have occurred. The model detailed by Mavrogianni et al. [
43] was used and appropriately modified for the tissues under evaluation in the present study. Based on the above, it was possible to calculate incidence rates of the various infections. Further, it was possible to estimate the length of time for which an animal was at risk before it became infected, as well as the length of time that an animal had been infected. Incidence rate was defined as the proportion of animals at risk, which developed the condition when the time at risk was the same in each group.
Linear mixed models were used in analysis to account for repeated measures of values of haematological parametres over the course of the study. Time points of collecting data were selected as within-subject variables and group allocation as a between-subject factor. Independent variables (fixed effects) included study group, sampling point and a sampling point–study group interaction.
The various associations were evaluated in a table of cross-categorised frequency data by use of the Pearson chi-square test or the Fisher exact test as appropriate.
In all cases, statistical significance was defined as p < 0.05.

 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}