
Influence of Soil-Borne Inoculum of Plasmodiophora brassicae Measured by qPCR on Disease Severity of Clubroot-Resistant Cultivars of Winter Oilseed Rape (Brassica napus L.)


Abstract
:1. Introduction
2. Results
2.1. Field Trials 2017–2018
2.1.1. Influence of Soil-Borne Inoculum of P. brassicae on Agronomic Performance of Winter OSR Cultivars
2.1.2. Visual Assessment of Clubroot Disease Severity and Disease Incidence 2017
2.2. Field Trials 2018–2019
2.2.1. Influence of Clubroot on Agronomic Performance of Winter OSR Cultivars
2.2.2. Visual Assessment of Clubroot Severity and Clubroot Incidence
2.3. Soil Bioassays and Real-Time qPCR Analysis
3. Discussion
4. Materials and Methods
4.1. Field Trial Assessment of Resistant Cultivars of Winter OSR
4.2. Soil Sampling and Preparation
4.2.1. Soil Sampling for Selecting Trial Fields
4.2.2. Soil Sampling for Determination of Physiochemical Parameters
4.2.3. Soil Sampling for Bioassays
4.2.4. Determination of Soil Chemical Properties
4.3. Soil Bioassay
4.4. Soil DNA Extraction and Real-Time PCR Analysis
4.5. Field Assessment of Disease
4.6. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ackman, R.D. Canola Fatty Acids- An Ideal Mixture for Health, Nutrition and Food Use. In Canola and Rapeseed: Production, Chemistry, Nutrition and Processing Technology, 1st ed.; Shahidi, F., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1990; pp. 81–98. [Google Scholar]
- Wallenhammar, A.-C.; Almquist, C.; Schwelm, A.; Roos, J.; Marcez-Schmidt, K.; Jonsson, A.; Dixelius, C. Clubroot, a persitent threat to Swedish oil seed rape production. Can. J. Plant Pathol. 2014, 36, 135–141. [Google Scholar] [CrossRef]
- Zheng, X.; Koopman, B.; Ulber, B.; von Tiderman, A. A Global Survey on Diseases and Pests in Oilseed Rape- Current Challenges and Innovative Strategies of Control. Front. Agron. 2020, 2, 15–21. [Google Scholar] [CrossRef]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Dixon, G.R. Clubroot (Plasmodiophora brassicae Woronin)- an agricultural and biological challenge world-wide. Can. J. Plant Pathol. 2014, 36, 5–18. [Google Scholar] [CrossRef]
- Sierra, R.; Canas-Duarte, S.J.; Burki, F.; Schwelm, A.; Fogelqvist, J.; Dixelius, C.; González-Garcia, L.N.; Gile, G.H.; Slamovits, C.; Restrepo, S.; et al. Evolutionary origins of Rhiazaran parasites. Mol. Biol. Evol. 2015, 33, 980–983. [Google Scholar] [CrossRef] [Green Version]
- Schwelm, A.; Fogelqvist, J.; Knaust, A.; Julke, S.; Lilja, T.; Bonilla-Rosso, G.; Karlsson, M.; Shevchenko, A.; Dhanapandi, V.; Ryun Choi, S.; et al. The Plasmodiophora brassicae genome reveals insightis its life cycle and ancestry of chitin synthases. Nat. Sci. Rep. 2015, 5, 1–12. [Google Scholar]
- Strelkov, S.E.; Hwang, S.-F. Clubroot in the Canadian Canola crop: 10 years into the outbreak. Can. J. Plant Pathol. 2014, 36, 27–36. [Google Scholar] [CrossRef]
- Stelkov, S.E.; Hwang, S.-F.; Maniolii, V.P.; Turnbull, G.; Freuda-Agyeman, R.; Hollman, K.; Kaus, S. Characterisation of clubroot (Plasmodiophora brassicae) from canola (Brassica napus) in the Peace Country of Alberta, Canada. Can. J. Plant Pathol. 2020, 1–7. [Google Scholar] [CrossRef]
- Chapara, V.; Kalwar, N.; Lubenow, L.; Chirumamilla, A. Prevalence of Clubroot on Canola in North Dakota. J. Agron. Agric. Sci. 2019, 2, 8. [Google Scholar] [CrossRef]
- Rastas, M.; Latvala, S.; Hannukkala, A. Occurrence of Plasmodiophora brassicae in Finnish turnip rape and oilseed rape fields. Agric. Food Sci. 2012, 21, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Botero, A.; García, C.; Gossen, B.D.; Strelkov, S.E.; Todd, C.D.; Bonham-Smith, P.C.; Pérez-López, E. Clubroot disease in Latin America: Distribution and management strategies. Plant Pathol. 2019, 68, 827–833. [Google Scholar] [CrossRef] [Green Version]
- McGrann, G.M.D.; Gladders, P.; Smith, J.A.; Burnett, F. Control of clubroot (Plasmodiophora brassicae) in oilseed rape using varietal resistance and soil amendments. Field Crop. Res. 2016, 186, 146–156. [Google Scholar] [CrossRef]
- Diedichsen, E.; Frauen, M.; Ludwig-Muller, J. Clubroot management challenges from a German perspective. Can. J. Plant Pathol. 2014, 36, 85–98. [Google Scholar] [CrossRef]
- Jedryczka, M.; Kasprzyk, I.; Korbas, M.; Jajor, E.; Kaczmarek, J. Infestation of Polish agricultural soils by Plasmodiophora brassicae along the Polish—Ukrainian border. J. Plant Prot. Res. 2016, 54, 23–241. [Google Scholar] [CrossRef] [Green Version]
- Czubbatka-Biénkowska, A.; Kaczmarek, J.; Marzec-Schmidt, M.; Nieróbca, A.; Czajka, A.; Jedryczka, M. Country- wide qPCR based assessment of Plasmodiophora brassicae spread in agricultural soils and recommendations for the cultivation of Brassicaceae crops in Poland. Pathogens 2020, 9, 1070. [Google Scholar] [CrossRef]
- Řičařová, V.; Kaczmarek, J.; Strelkov, S.E.; Kazda, J.; Lueders, W.; Rysanek, P.; Manolii, V.; Jedryczka, M. Pathotypes of Plasmodiophora brassicae causing damage to oilseed rape in the Czech Republic and Poland. Eur. J. Plant Pathol. 2016, 145, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Chai, A.L.; Xie, X.W.; Shi, Y.X.; Li, B.J. Research status of clubroot (Plasmodiophora brassicae) on cruciferous crops in China. Can. J. Plant Pathol. 2014, 36, 1421–1453. [Google Scholar] [CrossRef]
- Wallenhammar, A.-C.; Hansson, F.; Gunnarsson, A.; Jonsson, A. Quantification of Plasmodiophora brassicae Using a DNA-Based Soil Test Facilitates Sustainable Oilseed Rape Production. Plants 2016, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihlstrand, A. Biologisk markkartering håller koll på klumprotsjuka. Svensk Frötidning 2016, 2, 24. Available online: https://www.svenskraps.se/kunskap/pdf/01854.pdf (accessed on 15 January 2021). (In Swedish).
- Wallenhammar, A.-C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Crute, I.R.; Phelps, K.; Barnes, A.; Buczacki, S.T.; Crisp, P. The relationship between genotypes of three Brassica species and collections of Plasmodiophora brassicae Woron. Plant Pathol. 1983, 32, 405–420. [Google Scholar] [CrossRef]
- Jönsson, R. Klumprotsjuka. Litteraturöversikt och resultat av resistensarbeten i oljeväxter vid Svalöf 1962–1968. Sveriges Utsädesförenings Tidskr. 1969, 79, 28–56. (In Swedish) [Google Scholar]
- Strelkov, S.E.; Hwang, S.-F.; Manolii, V.P.; Cao, T.; Feindel, D. Emergence of new virulence phenotypes pf Plasmodiophora brassicae on canola (Brassica napus). Eur. J. Plant Pathol. 2016, 145, 517–529. [Google Scholar] [CrossRef]
- Zamani-Noor, N. Variation in pathotypes and virulence of Plasmodiophora brassicae populations in Germany. Plant Pathol. 2017, 66, 316–324. [Google Scholar] [CrossRef]
- Cao, T.; Manolii, V.P.; Hwang, S.F.; Howard, R.J.; Strelkov, S.E. Virulence and spread of Plasmodiophora brassicae [clubroot] in Alberta, Canada. Can. J. Plant Pathol. 2009, 31, 321–329. [Google Scholar] [CrossRef]
- Rennie, D.C.; Holtz, M.D.; Turkington, T.K.; Leboldous, J.M.; Hwang, S.-F.; Howard, R.J.; Strelkov, S.E. Movement of Plasmodiophora brassicae resting spores in windblown dust. Can. J. Plant Pathol. 2015, 37, 188–196. [Google Scholar] [CrossRef]
- Faggian, R.; Bulman, S.R.; Lawrie, A.C.; Porter, I.J. Specific polymerase chain reaction primers for the detection of Plasmodiophora brassicae in soil and water. Phytopathology 1999, 89, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, K.; Asano, T. Life cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 2009, 28, 203–211. [Google Scholar] [CrossRef]
- Ludvig-Müller, J. Glucosinolates and the clubroot disease: Defense compounds or auxin precursors? Phytochem. Rev. 2009, 8, 135–148. [Google Scholar] [CrossRef]
- Wallenhammar, A.-C.; Almquist, C.; Söderström, M.; Jonsson, A. In-field distribution of Plasmodiophoraa brasssice measured using quantitative real-time PCR. Plant Pathol. 2012, 61, 16–28. [Google Scholar] [CrossRef]
- Jonsson, A.; Wallenhammar, A.-C.; Dixelius, C. Long-term soil data sets reveals shifts in population of Plasmodiophora brassica. In Proceedings of the 6th International Symposium on Brassica and 18th Crucifer Genetics Workshop, Catania, Italy, 12–16 November 2012. [Google Scholar]
- Zamani-Noor, N.; Krohne, I.; Koopmann, B. Greenhouse Evaluation of Clubroot Resistant-Brassica napus cv. Mendel and Its Efficacy Concerning Virulence and Soil Inoculum Levels of Plasmodiophora brassicae. Pathogens 2021, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Wallenhammar, A.-C.; Johnsson, L.; Gerhardson, B. Agronomic performance of partly clubroot-resistant spring oilseed turnip rape. J. Phytopathol. 2000, 148, 495–499. [Google Scholar] [CrossRef]
- Gunnarsson, A. Höstrapssorter med klumprotresistens. Field Trial Report of central Sweden. Rural Economy and Agricultural Society Skaraborg: Falköping 2015, 155–156. (In Swedish) [Google Scholar]
- Diedichsen, E.; Wagenblatt, B.; Schallehn, V. Production of pure genotype isolates of Plasmodiophora brassicae Wor.—Comparison of inoculations with root hairs containing single sporangiosori or with single resting spores. Eur. J. Plant Pathol. 2016, 145, 621–627. [Google Scholar] [CrossRef]
- Hwang, S.-F.; Strelkov, S.E.; Feng, J.; Gossen, B.D.; Howard, R.J. Plasmodiophora brassicae: A review of an emerging a pathogen of the Canadian canola (Brassica napus) crop. Mol. Plant Pathol. 2012, 13, 105–113. [Google Scholar] [CrossRef]
- Peng, G.; Lahlali, R.; Hwang, S.-F.; Pageau, D.; Hynes, R.K.; McDonald, M.R.; Gossen, B.D.; Strelkov, S.E. Crop rotation, cultivar resistance, and fungicides/ biofungicides for managing clubroot (Plasmodiophora brassice) on canola. Can. J. Plant Pathol. 2014, 36, 99–112. [Google Scholar] [CrossRef]
- Wallenhammar, A.-C. Observations from yield loss from Plasmodiophora brassicae infections in spring oilseed rape. J. Plant Dis. Prot. 1998, 105, 1–7. [Google Scholar]
- Liu, L.; Qin, L.; Cheng, X.; Zhang, Y.; Xu, L.; Liu, F.; Tong, C.; Huang, J.; Liu, S.; Wei, Y. Comparing the Infection Biology of Plasmodiophora brassicae in Clubroot Susceptible and Resistant Hosts and Non-hosts. Front Microbiol. 2020, 11, 507036. [Google Scholar] [CrossRef] [PubMed]
- Ernst, T.W.; Kher, S.; Stanton, D.; Rennie, D.C.; Hwang, S.F.; Strelkow, S.E. Plasmodiophora brassicae resting spore dynamics in clubroot resistant canola (Brassica napus) cropping systems. Plant Pathol. 2018, 68, 399–408. [Google Scholar] [CrossRef]
- Dobson, R.; Gabrielson, R.L.; Baker, A.S. Soil Water Matric Potential Requirements for Root-Hair and Cortical infection of Chinese Cabbage by Plasmodiophora brassicae. Phytopathology 1982, 12, 1598–1600. [Google Scholar] [CrossRef]
- Wellman, F.L. Clubroot of crucifers. Tech. Bull. U. S. Dept. Agric. 1930, 181, 1–31. [Google Scholar]
- Monteith, J. Relation of soil temperature and soil moisture to infection by Plasmodiophora brassicae. J. Agric. Res. 1924, 28, 549–561. [Google Scholar]
- Gossen, B.D.; Adhikari, K.K.C.; McDonald, M.R. Effects on temperature of infection and subsequent development of clubroot under controlled conditions. Plant Pathol. 2012, 61, 593–599. [Google Scholar] [CrossRef]
- Zhar, K.; Sarkes, A.; Yang, Y.; Zhou, Q.; Feindel, D.; Harding, M.W. Plasmodiophora brassicae in its environment- effects of temperature and light on resting spore survival in soil. Phytopathology 2021. [CrossRef]
- Dixon, G.R. Plasmodiophora brassicae in its environment. J. Plant Growth Regul. 2009, 28, 212–228. [Google Scholar] [CrossRef]
- Mattsson, L. Skördevariationer Inom ENSKILDA fält. Storlek och Tänkbara Orsaker; Report 196; Department of Soil Sciences, Division of Soil Fertility, Swedish University of Agricultural Sciences: Uppsala, Sweden, 1995. (In Swedish) [Google Scholar]
- Strelkov, S.E.; Hwang, S.-F.; Maniolii, V.P.; Cao, T.; Freuda-Agyeman, R.; Harding, M.W.; Peng, G.; Gossen, B.D.; McDonald, M.R.; Fiendel, D. Virulence and pathotype classification of Plasmodiophora brassicae populations collected from clubroot resistant canola (Brassica napus) in Canada. Can. J. Plant Pathol. 2018, 40, 284–298. [Google Scholar] [CrossRef]
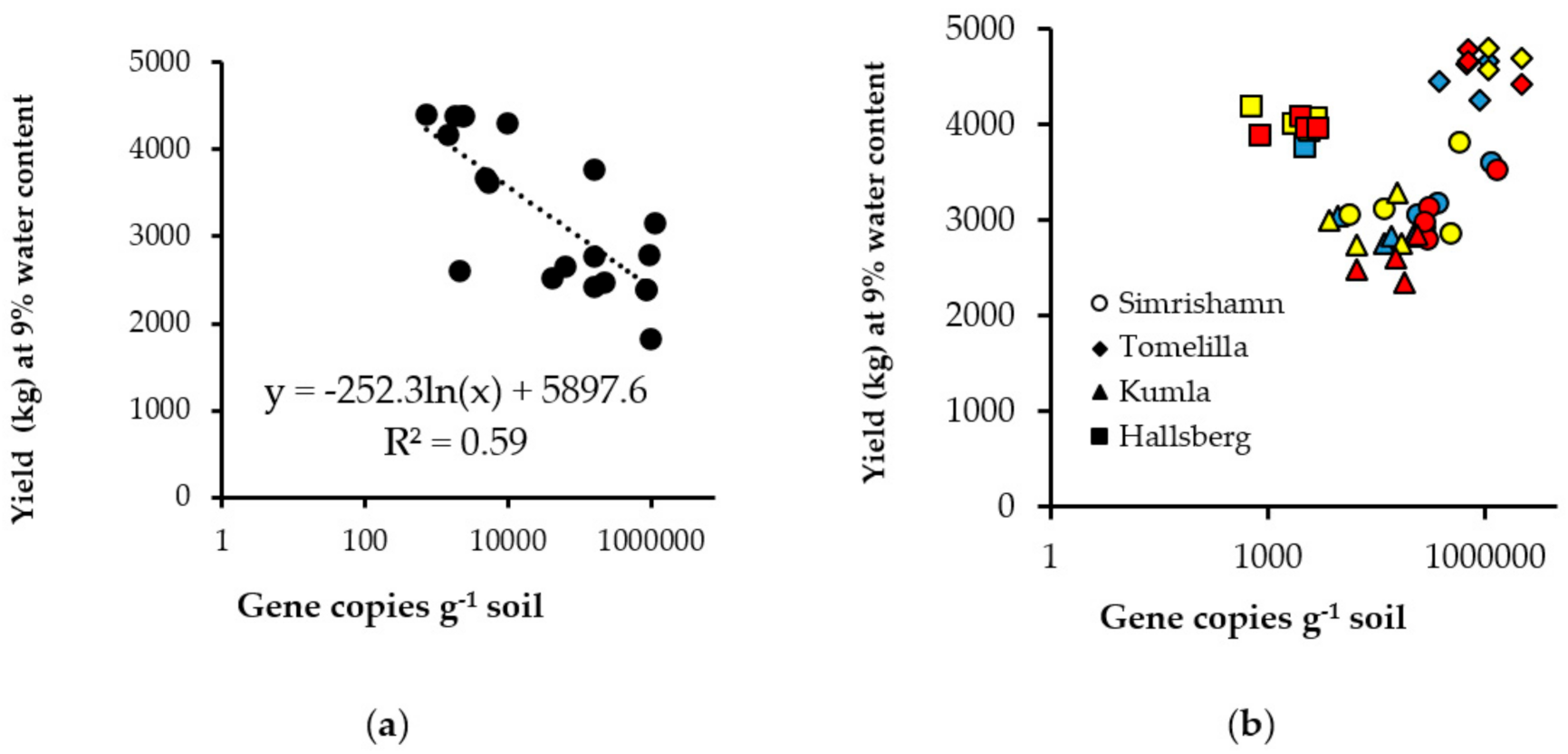
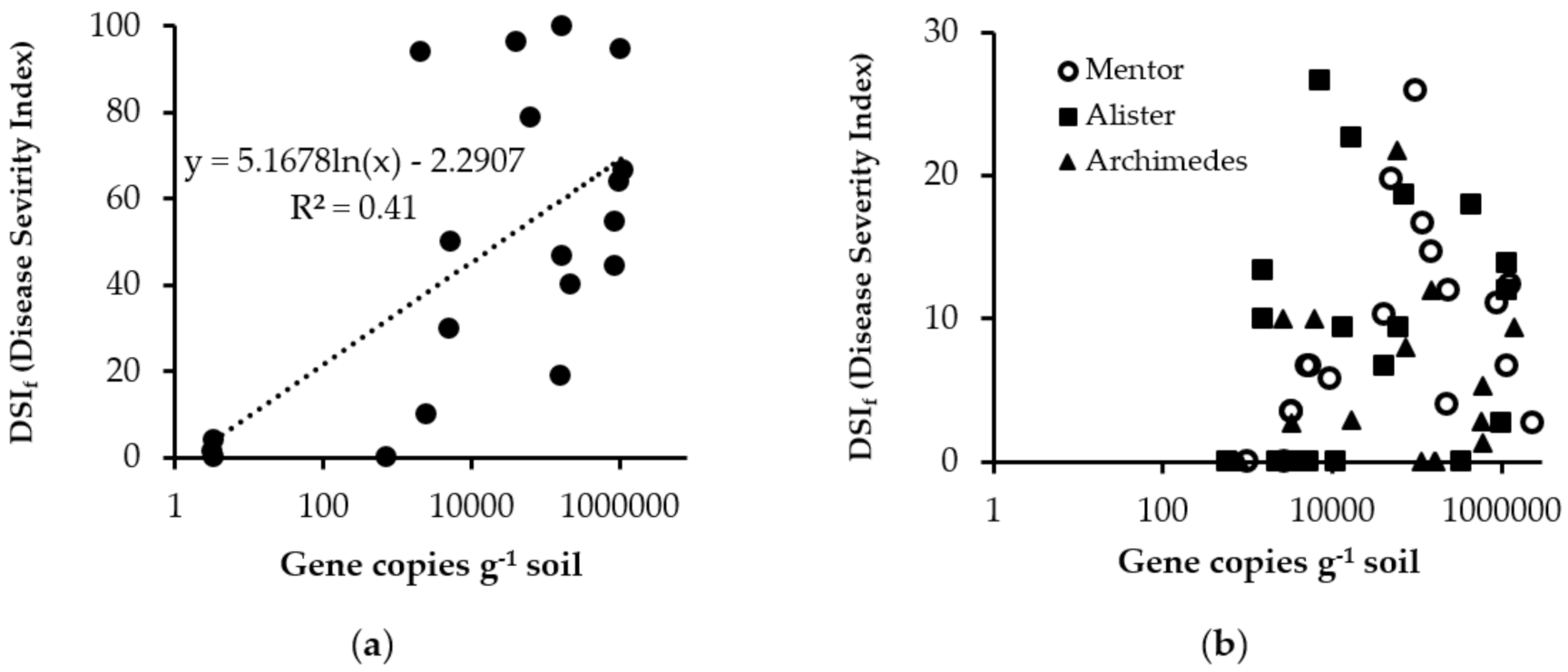



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment and Location | DSIf | DIf (%) | ||
|---|---|---|---|---|
| Simrishamn | ||||
| Cultivar mix | 52.1 | b * | 67.9 | b |
| cv. Mentor | 12.0 | c | 15.6 | cd |
| cv. Alister | 8.5 | c | 10.8 | cd |
| cv. Archimedes | 5.3 | c | 6.0 | cd |
| Tomelilla | ||||
| Cultivar mix | 55.4 | b | 62.5 | b |
| cv. Mentor | 8.3 | c | 11.2 | cd |
| cv. Alister | 8.0 | c | 9.2 | cd |
| cv. Archimedes | 2.4 | c | 4.0 | cd |
| Kumla | ||||
| Cultivar mix | 92.3 | a | 100.0 | a |
| cv. Mentor | 15.5 | c | 15.1 | cd |
| cv. Alister | 19.5 | c | 16.9 | c |
| cv. Archimedes | 8.3 | c | 6.4 | cd |
| Hallsberg | ||||
| Cultivar mix | 1.8 | c | 2.7 | cd |
| cv. Mentor | 0 | c | 0 | d |
| cv. Alister | 0 | c | 0 | d |
| cv. Archimedes | 0.7 | c | 2.0 | cd |
| p-value | <0.001 | <0.001 | ||
| Coefficient of variation | 56.1 | 47.4 | ||
| DSIf (0–100) | DIf (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | Autumn 2018 | July 2019 | Both years | Autumn 2018 | July 2019 | Both years | ||||
| Simrishamn | ||||||||||
| Cultivar mix | 3.3 | 0.0 | 1.7 | 5.0 | 0.0 | 2.5 | ||||
| cv. Mentor | 0.0 | 0.7 | 0.3 | 0.0 | 2.0 | 1.0 | ||||
| cv. Alister | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||||
| cv. Archimedes | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||||
| p-value | ns | ns | ns | ns | ns | ns | ||||
| Coeff. of var. † | 400 | 231 | 475 | 400 | 231 | 418 | ||||
| Tomelilla | ||||||||||
| Cultivar mix | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||||
| cv. Mentor | 0.0 | 1.0 | 0.5 | 0.0 | 1.0 | 1.5 | ||||
| cv. Alister | 1.9 | 0.0 | 0.9 | 5.6 | 0.0 | 2.8 | ||||
| cv. Archimedes | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||||
| p-value | ns | ns | ns | ns | ns | ns | ||||
| Coeff. of var. | 231 | 400 | 311 | 231 | 400 | 311 | ||||
| Kumla | ||||||||||
| Cultivar mix | 22.5 | 27.5 | a * | 25.0 | a | 25.0 | 43.2 | a | 34.1 | a |
| cv. Mentor | 3.3 | 6.9 | ab | 5.9 | b | 7.5 | 10.9 | ab | 9.2 | b |
| cv. Alister | 5.8 | 1.7 | b | 5.1 | b | 7.5 | 8.0 | ab | 4.8 | b |
| cv. Archimedes | 6.7 | 5.3 | b | 3.8 | b | 6.7 | 2.0 | b | 7.4 | b |
| p-value | ns | 0.016 | 0.001 | ns | 0.025 | 0.004 | ||||
| Coeff. of var. | 128 | 99 | 106 | 131 | 109 | 115 | ||||
| Treatment | DSIb (0–100) | DSIb (%) | Gene Copies (No. g soil−1) | DSIb × Gene Copies | DIb × Gene Copies | |||
|---|---|---|---|---|---|---|---|---|
| Gene copies P. brassicae, average: | ||||||||
| Simrishamn | 419,200 | b * | ||||||
| Tomelilla | 1,187,100 | a | ||||||
| Kumla | 55,600 | c | ||||||
| Hallsberg | 2800 | c | ||||||
| p-value | <0.001 | |||||||
| Coeff. of var. † | 119 | |||||||
| Field-wise: | ||||||||
| Simrishamn | ||||||||
| Cultivar mix | 77.5 | a * | 94.5 | a | 562,300 | p = 0.035 | ns | |
| cv. Mentor | 21.9 | bcd | 30.5 | bcd | 435,700 | p < 0.001 | p < 0.001 | |
| cv. Alister | 38.6 | b | 49.2 | b | 204,100 | p = 0.012 | p = 0.027 | |
| cv. Archimedes | 34.4 | b | 46.6 | b | 474,700 | ns | ns | |
| p-value | 0.020 | 0.020 | ns | |||||
| Coeff. of var. | 141 | 134 | 103 | |||||
| Tomelilla | ||||||||
| Cultivar mix | 75.2 | a | 86.4 | a | 784,300 | ns | ns | |
| cv. Mentor | 31.6 | b | 40.2 | bc | 1,129,800 | ns | ns | |
| cv. Alister | 32.8 | b | 42.9 | b | 1,603,200 | ns | ns | |
| cv. Archimedes | 7.0 | de | 16.5 | cde | 1,231,000 | ns | ns | |
| p-value | <0.001 | ns | ns | |||||
| Coeff. of var. | 71 | 66 | 72 | |||||
| Kumla | ||||||||
| Cultivar mix | 29.9 | bc | 42.0 | bc | 68,300 | ns | ns | |
| cv. Mentor | 9.0 | cde | 14.8 | de | 49,500 | p = 0.011 | p = 0.009 | |
| cv. Alister | 7.5 | cde | 9.5 | de | 38,800 | ns | ns | |
| cv. Archimedes | 4.4 | de | 8.2 | de | 65,800 | p = 0.017 | p = 0.009 | |
| p-value | ns | ns | ns | |||||
| Coeff. of var. | 323 | 323 | 76 | |||||
| Hallsberg | ||||||||
| Cultivar mix | 1.3 | de | 2.8 | e | 3800 | ns | ns | |
| cv. Mentor | 0.5 | e | 1.2 | e | 1700 | ns | ns | |
| cv. Alister | 0.8 | e | 1.8 | e | 2800 | ns | ns | |
| cv. Archimedes | 0.0 | e | 0.0 | e | 3020 | ns | ns | |
| p-value | ns | ns | ns | |||||
| Coeff. of var. | 323 | 323 | 77 | |||||
| Treatment | DSIb (0–100) | DIb (%) | Gene Copies (No. g soil−1) | DSIb × Gene Copies | DIb × Gene Copies | |||
|---|---|---|---|---|---|---|---|---|
| Gene copies P. brassicae, average | ||||||||
| Simrishamn | 442,300 | a * | ||||||
| Tomelilla | 38,000 | b | ||||||
| Kumla | 3700 | b | ||||||
| P-value | <0.001 | |||||||
| Coeff. of var. † | 113 | |||||||
| Field wise | ||||||||
| Simrishamn | ||||||||
| Cultivar mix | 15.8 | a | 19.4 | a | 633,900 | a | ns | ns |
| cv. Mentor | 4.6 | a | 5.6 | b | 471,500 | ab | ns | ns |
| cv. Alister | 16.2 | a | 19.2 | a | 299,900 | b | ns | ns |
| cv. Archimedes | 4.0 | a | 5.8 | b | 364,300 | ab | ns | ns |
| p-value | 0.020 | 0.020 | 0.015 | |||||
| Coeff. of var. | 141 | 134 | 4.0 | |||||
| Tomelilla | ||||||||
| Cultivar mix | 54.6 | a | 61.8 | 48,400 | a | ns | ns | |
| cv. Mentor | 22.9 | b | 25.7 | 42,400 | ab | ns | ns | |
| cv. Alister | 29.8 | b | 33.7 | 27,300 | c | ns | ns | |
| cv. Archimedes | 14.6 | b | 16.9 | 35,200 | bc | ns | ns | |
| p-value | <0.001 | ns | <0.001 | |||||
| Coeff. of var. | 71.2 | 66.2 | 2.9 | |||||
| Kumla | ||||||||
| Cultivar mix | 2.5 | 2.5 | a | 3400 | ns | ns | ||
| cv. Mentor | 0.7 | 0.7 | ab | 3500 | ns | ns | ||
| cv. Alister | 0.0 | 0.0 | b | 4000 | ns | ns | ||
| cv. Archimedes | 1.7 | 1.7 | b | 3600 | ns | ns | ||
| p-value | ns | ns | ns | |||||
| Coeff. of var. | 323 | 323 | 75 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallenhammar, A.-C.; Omer, Z.S.; Edin, E.; Jonsson, A. Influence of Soil-Borne Inoculum of Plasmodiophora brassicae Measured by qPCR on Disease Severity of Clubroot-Resistant Cultivars of Winter Oilseed Rape (Brassica napus L.). Pathogens 2021, 10, 433. https://doi.org/10.3390/pathogens10040433
Wallenhammar A-C, Omer ZS, Edin E, Jonsson A. Influence of Soil-Borne Inoculum of Plasmodiophora brassicae Measured by qPCR on Disease Severity of Clubroot-Resistant Cultivars of Winter Oilseed Rape (Brassica napus L.). Pathogens. 2021; 10(4):433. https://doi.org/10.3390/pathogens10040433
Chicago/Turabian StyleWallenhammar, Ann-Charlotte, Zahra Saad Omer, Eva Edin, and Anders Jonsson. 2021. "Influence of Soil-Borne Inoculum of Plasmodiophora brassicae Measured by qPCR on Disease Severity of Clubroot-Resistant Cultivars of Winter Oilseed Rape (Brassica napus L.)" Pathogens 10, no. 4: 433. https://doi.org/10.3390/pathogens10040433