
Characterization of a Dengue Virus Serotype 1 Isolated from a Patient in Ciudad Juarez, Mexico

 , , and
, , and
Abstract
:1. Introduction
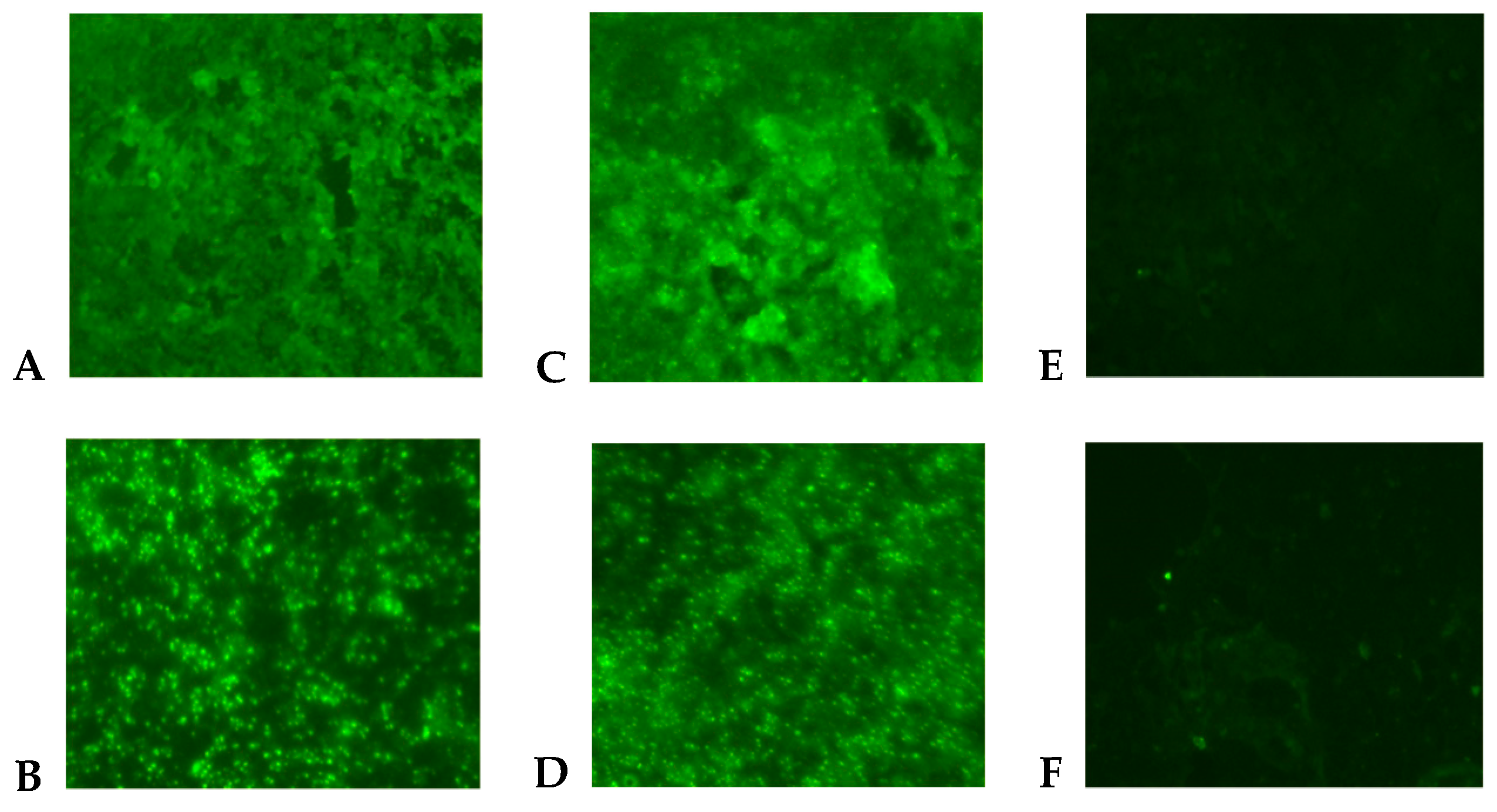
2. Results
3. Discussion
4. Materials and Methods
4.1. Patient and Sample Collection and Processing
4.2. Viral Isolation
4.3. Immunofluorescence Assay (IFA)
4.4. Viremia Levels
4.5. Molecular Identification
4.6. Envelope Gene Sequencing
4.7. Viral Whole-Genome Sequencing
4.8. Replication Curve
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci. Data 2015, 2, 150035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, M.B.; Eisen, L.; McAllister, J.; Savage, H.M.; Mutebi, J.P.; Eisen, R.J. Updated Reported Distribution of Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in the United States, 1995–2016. J. Med. Entomol. 2017, 5, 1420–1424. [Google Scholar] [CrossRef] [PubMed]
- San Martín, J.L.; Brathwaite, O.; Zambrano, B.; Solórzano, J.O.; Bouckenooghe, A.; Dayan, G.H.; Guzmán, M.G. The epidemiology of dengue in the Americas over the last three decades: A worrisome reality. Am. J. Trop. Med. Hyg. 2010, 1, 128–135. [Google Scholar] [CrossRef]
- De Lourdes Muñoz, M.; Mercado-Curiel, R.F.; Diaz-Badillo, A.; Pérez Ramirez, G.; Black, W.C. Gene flow pattern among Aedes aegypti populations in Mexico. J. Am. Mosq Control. Assoc. 2013, 29, 1–18. [Google Scholar] [CrossRef] [PubMed]
- De la Mora-Covarrubias, A.; Jiménez-Vega, F.; Treviño-Aguilar, S.M. Geospatial distribution and detection of dengue virus in Aedes (Stegomyia) aegypti mosquitos in Ciudad Juárez, Chihuahua, Mexico. Salud Publica Mex. 2010, 52, 127–133. [Google Scholar] [PubMed]
- Palermo, P.M.; De la Mora-Covarrubias, A.; Jimenez-Vega, F.; Watts, D.M. Serological evidence of Dengue and West Nile Virus human infection in Juarez City, Mexico. Vector Borne Zoonotic Dis. 2019, 2, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Jarman, R.G.; Nisalak, A.; Anderson, K.B.; Klungthong, C.; Thaisomboonsuk, B.; Kaneechit, W.; Kalayanarooj, S.; Gibbons, R.V. Factors influencing dengue virus isolation by C6/36 cell culture and mosquito inoculation of nested PCR-positive clinical samples. Am. J. Trop. Med. Hyg. 2011, 2, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Arredondo-García, J.L.; Aguilar-López, E.G.; Aguilar Lugo-Gerez, J.; Osnaya-Romero, N.; Pérez-Guillé, G.; Medina-Cortina, H. Epidemiological panorama of dengue in Mexico 2000–2019. Rev. Latin Infect. Pediatr. 2020, 33, 78–83. [Google Scholar]
- González-Durán, E.; Vázquez-Pichardo, M.; Torres-Flores, J.M.; Garcés-Ayala, F.; Méndez-Tenorio, A.; Curiel-Quesada, E.; Ortiz-Alcántara, J.M.; Castelán-Sánchez, H.G.; Salas-Benito, J.S.; Torres-Longoria, B.; et al. Genotypic variability analysis of DENV-1 in Mexico reveals the presence of a novel Mexican lineage. Arch. Virol. 2018, 163, 1643–1647. [Google Scholar] [CrossRef]
- Villabona-Arenas, C.J.; Zanotto, P.M. Worldwide spread of Dengue virus type 1. PLoS ONE 2013, 8, e62649. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.E.; Martin, D.P.; Oliveira, L.M.; Ribeiro, B.M.; Nagata, T. Comparative analysis of American Dengue virus type 1 full-genome sequences. Virus Genes 2010, 40, 60–66. [Google Scholar] [CrossRef]
- Prestwood, T.R.; Prigozhin, D.M.; Sharar, K.L.; Zellweger, R.M.; Shresta, S. A mouse-passaged dengue virus strain with reduced affinity for heparan sulfate causes severe disease in mice by establishing increased systemic viral loads. J. Virol. 2010, 82, 8411–8421. [Google Scholar] [CrossRef] [Green Version]
- Duarte dos Santos, C.N.; Frenkiel, M.P.; Courageot, M.P.; Rocha, C.F.; Vazeille-Falcoz, M.C.; Wien, M.W.; Rey, F.A.; Deubel, V.; Desprès, P. Determinants in the envelope E protein and viral RNA helicase NS3 that influence the induction of apoptosis in response to infection with dengue type 1 virus. Virology 2000, 274, 292–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OhAinle, M.; Balmaseda, A.; Macalalad, A.R.; Tellez, Y.; Zody, M.C.; Saborío, S.; Nuñez, A.; Lennon, N.J.; Birren, B.W.; Gordon, A.; et al. Dynamics of dengue disease severity determined by the interplay between viral genetics and serotype-specific immunity. Sci. Transl. Med. 2011, 3, 114ra128. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Rodpradit, P.; Chinnawirotpisan, P.; Mammen, M.P.; Li, T., Jr.; Lynch, J.A.; Putnak, R.; Zhang, C. Comparative analysis of full-length genomic sequences of 10 dengue serotype 1 viruses associated with different genotypes, epidemics, and disease severity isolated in Thailand over 22 years. Am. J. Trop Med. Hyg. 2010, 83, 1156–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Direccion General de Epidemiologia. Mexico. Panorama Epidemiologico de Dengue 2015. Available online: https://www.gob.mx/cms/uploads/attachment/file/45495/Pano_dengue_sem_52_2015.pdf (accessed on 21 May 2021).
- Direccion General de Epidemiologia. Mexico. Panorama Epidemiologico de Dengue 2016. Available online: https://www.gob.mx/cms/uploads/attachment/file/178952/Pano_dengue_sem_52_2016.pdf (accessed on 21 May 2021).
- Direccion General de Epidemiologia. Mexico. Panorama Epidemiologico de Dengue 2017. Available online: https://www.gob.mx/cms/uploads/attachment/file/285237/Pano_dengue_sem_52_2017.pdf (accessed on 21 May 2021).
- Direccion General de Epidemiologia. Mexico. Panorama Epidemiologico de Dengue 2019. Available online: https://www.gob.mx/cms/uploads/attachment/file/524262/Pano_dengue_52_2019.pdf (accessed on 21 May 2021).
- Rawlings, J.A.; Hendricks, K.A.; Burgess, C.R.; Campman, R.M.; Clark, G.G.; Tabony, L.J.; Patterson, M.A. Dengue surveillance in Texas, 1995. Am. J. Trop. Med. Hyg. 1998, 59, 95–99. [Google Scholar] [CrossRef]
- Ramos, M.M.; Mohammed, H.; Zielinski-Gutierrez, E.; Hayden, M.H.; Lopez, J.L.; Fournier, M.; Trujillo, A.R.; Burton, R.; Brunkard, J.M.; Anaya-Lopez, L.; et al. Epidemic dengue and dengue hemorrhagic fever at the Texas-Mexico border: Results of a household-based seroepidemiologic survey, December 2005. Am. J. Trop. Med. Hyg. 2008, 78, 364–369. [Google Scholar] [CrossRef]
- Thomas, D.L.; Santiago, G.A.; Abeyta, R.; Hinojosa, S.; Torres-Velasquez, B.; Adam, J.K.; Evert, N.; Caraballo, E.; Hunsperger, E.; Muñoz-Jordán, J.L.; et al. Reemergence of dengue in southern Texas, 2013. Emerg. Infect. Dis. 2016, 22, 1002–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.J.; Chen, S.L.; Chien, L.J.; Chen, C.C.; King, C.C.; Harn, M.R.; Hwang, K.P.; Fang, J.H. Silent transmission of the dengue virus in southern Taiwan. Am. J. Trop. Med. Hyg. 1996, 1, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, A.C.; Fernandez-Sesma, A. The burden of dengue and chikungunya worldwide: Implications for the Southern United States and California. Ann. Glob. Health 2014, 80, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morens, D.M.; Halstead, S.B.; Repik, P.M.; Putvatana, R.; Raybourne, N. Simplified plaque reduction neutralization assay for dengue viruses by semimicro methods in BHK-21 cells: Comparison of the BHK suspension test with standard plaque reduction neutralization. J. Clin. Microbiol. 1985, 22, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Díaz, F.J.; Black, W.C.; Farfán-Ale, J.A.; Loroño-Pino, M.A.; Olson, K.E.; Beaty, B.J. Dengue virus circulation and evolution in Mexico: A phylogenetic perspective. Arch. Med. Res. 2006, 37, 760–773. [Google Scholar] [CrossRef]
- Warrilow, D.; Northill, J.A.; Pyke, A.T. Sources of dengue viruses imported into Queensland, australia, 2002–2010. Emerg. Infect. Dis. 2012, 18, 1850–1857. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Name | Collection Year | Strain Name | GenBank Accession No. | NS1 | NS2A | NS3 | ||
|---|---|---|---|---|---|---|---|---|
| 139 | 41 | 218 | 332 | 338 | ||||
| DENV1_Mexico_2006_BID_V3658 | 2006 | BID_V3658 | GU131958.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V3673 | 2007 | BID-V3673 | GU131964.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V3679 | 2007 | BID-V3679 | GU131966.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V3709 | 2007 | BID-V3709 | GQ868513.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V3727 | 2007 | BID-V3727 | GQ868521.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V3739 | 2007 | BID-V3739 | GQ868527.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V7572 | 2007 | BID-V7572 | KJ189317.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V7589 | 2007 | BID-V7589 | KJ189323.1 | D | L | K | V | R |
| DENV1_Mexico_2007_BID-V7590 | 2007 | BID-V7590 | KJ189324.1 | D | L | K | V | R |
| DENV1_Mexico_2008_BID-V3758 | 2008 | BID-V3758 | GQ868537.1 | D | L | K | V | R |
| DENV1_Mexico_2011_BID-V7298 | 2011 | BID-V7298 | KJ189306.1 | D | L | K | V | R |
| DENV1_Mexico_2011_BID-V7624 | 2011 | BID-V7624 | KJ189348.1 | D | L | K | V | R |
| DENV1_Mexico_2011_BID-V7625 | 2011 | BID-V7625 | KJ189349.1 | D | L | K | V | R |
| DENV1_Mexico_2012_BID-V8195 | 2012 | BID-V8195 | KJ189368.1 | D | L | K | V | R |
| DENV1_Mexico_2015_CiudadJuarez | 2015 | CiudadJuarez | MZ343259 | E | F | R | A | K |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palermo, P.M.; de la Mora-Covarrubias, A.; Orbegozo, J.; Plante, J.A.; Plante, K.S.; Jimenez-Vega, F.; Watts, D.M. Characterization of a Dengue Virus Serotype 1 Isolated from a Patient in Ciudad Juarez, Mexico. Pathogens 2021, 10, 872. https://doi.org/10.3390/pathogens10070872
Palermo PM, de la Mora-Covarrubias A, Orbegozo J, Plante JA, Plante KS, Jimenez-Vega F, Watts DM. Characterization of a Dengue Virus Serotype 1 Isolated from a Patient in Ciudad Juarez, Mexico. Pathogens. 2021; 10(7):872. https://doi.org/10.3390/pathogens10070872
Chicago/Turabian StylePalermo, Pedro M., Antonio de la Mora-Covarrubias, Jeanette Orbegozo, Jessica A. Plante, Kenneth S. Plante, Florinda Jimenez-Vega, and Douglas M. Watts. 2021. "Characterization of a Dengue Virus Serotype 1 Isolated from a Patient in Ciudad Juarez, Mexico" Pathogens 10, no. 7: 872. https://doi.org/10.3390/pathogens10070872
APA StylePalermo, P. M., de la Mora-Covarrubias, A., Orbegozo, J., Plante, J. A., Plante, K. S., Jimenez-Vega, F., & Watts, D. M. (2021). Characterization of a Dengue Virus Serotype 1 Isolated from a Patient in Ciudad Juarez, Mexico. Pathogens, 10(7), 872. https://doi.org/10.3390/pathogens10070872