
2′FL and LNnT Exert Antipathogenic Effects against C. difficile ATCC 9689 In Vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. HMOs Exerted Potent Antimicrobial Effects against C. difficile during 48 h Fecal Batch Incubations, Which Was Likely Mediated via Microbiome Modulation
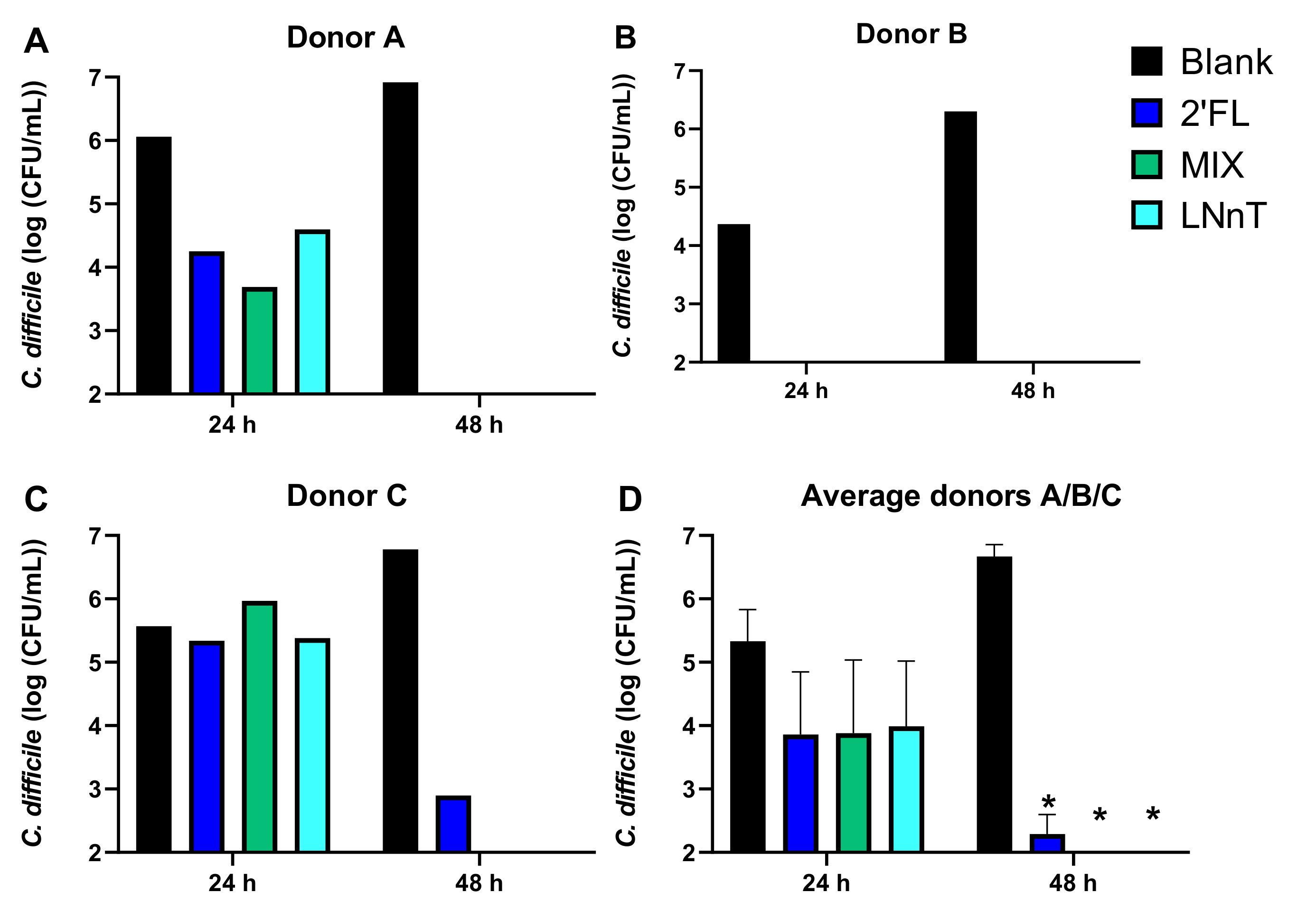
2.1.1. C. difficile Levels
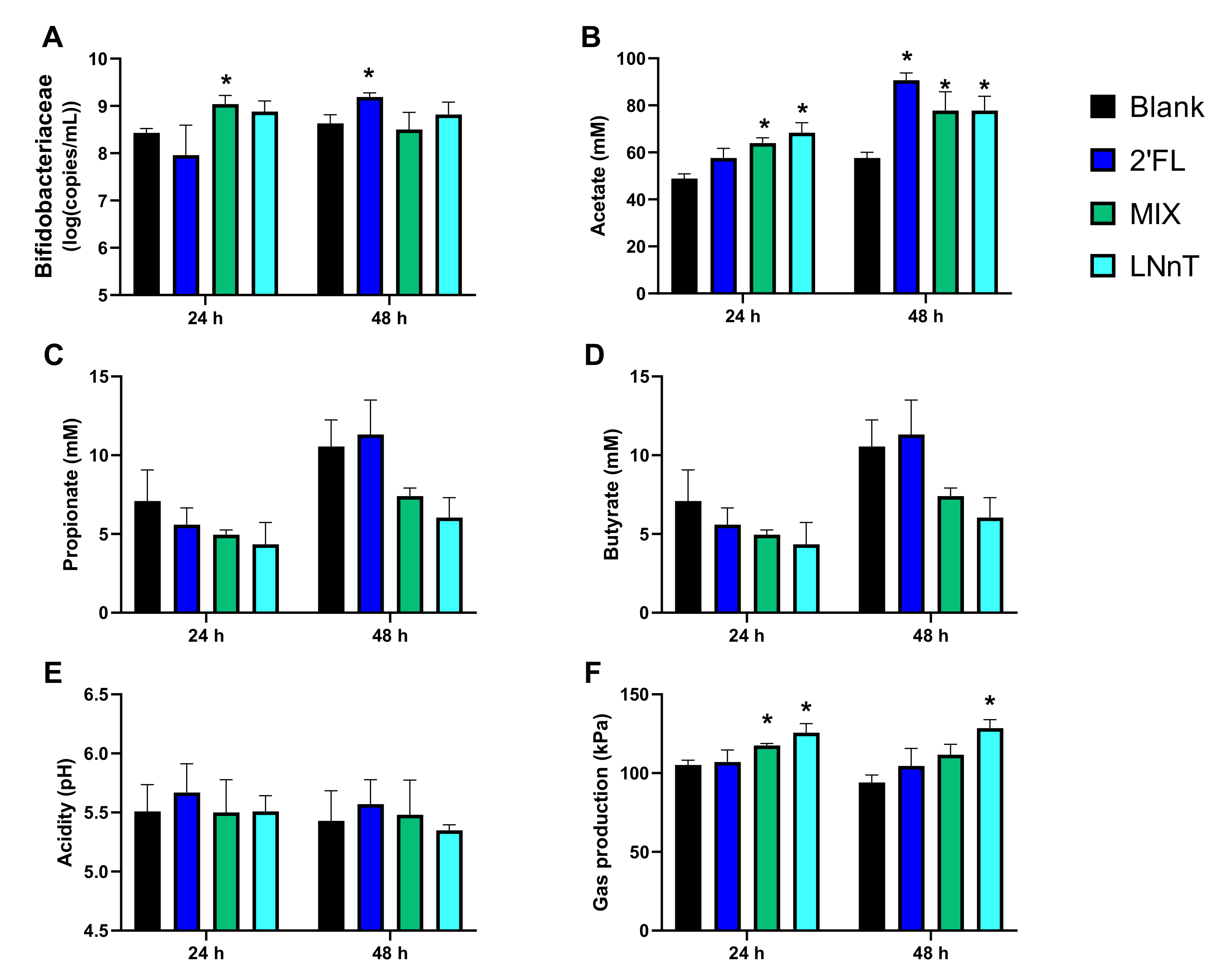
2.1.2. Microbial Activity and Bifidobacteriaceae Levels
2.2. 2′FL and LNnT Boosted Microbial Activity during Vancomycin Treatment, Coincinding with the Absence of CDI Recurrence for 2′FL during the Long-Term PathogutTM Study (Test Two)
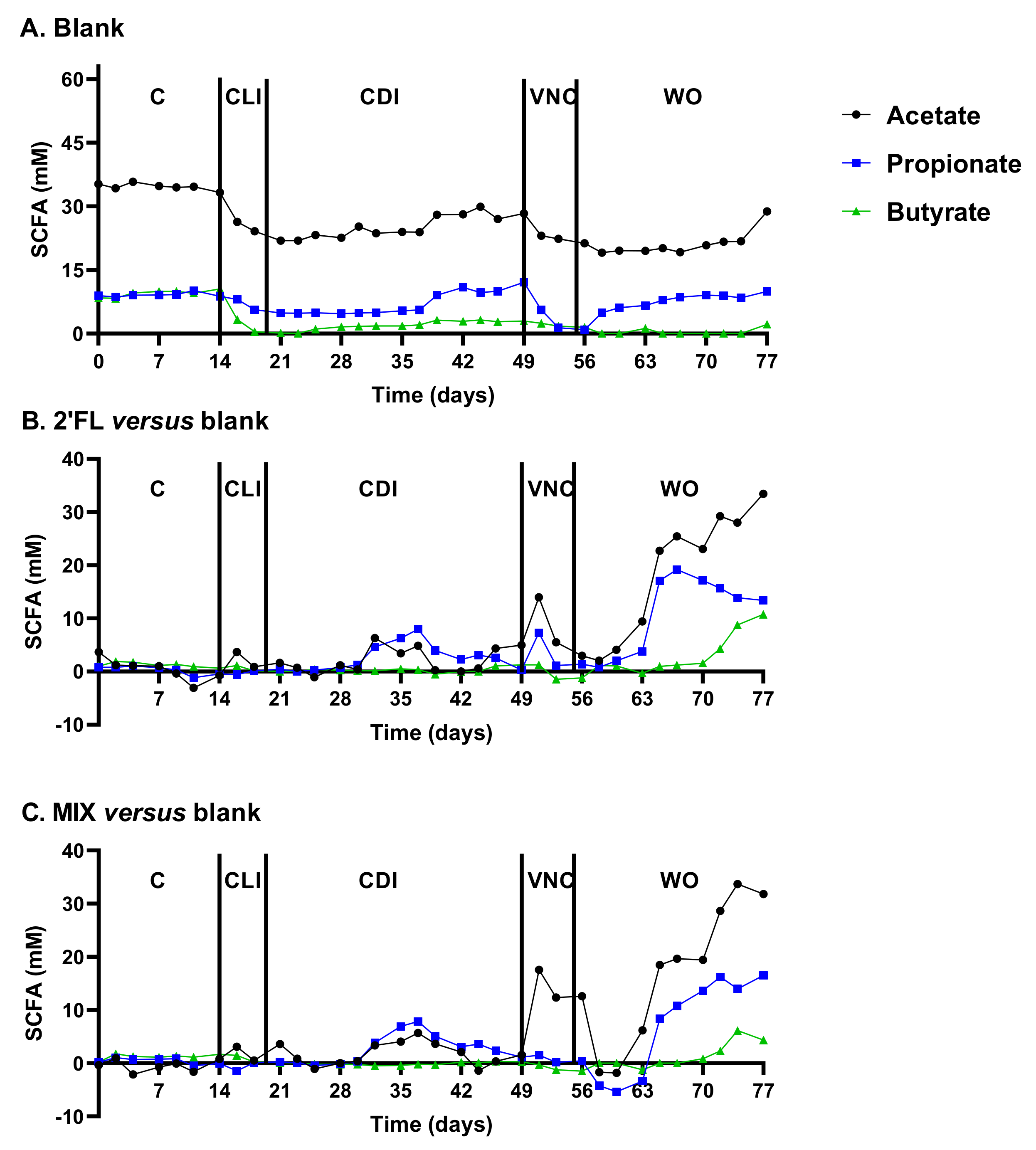
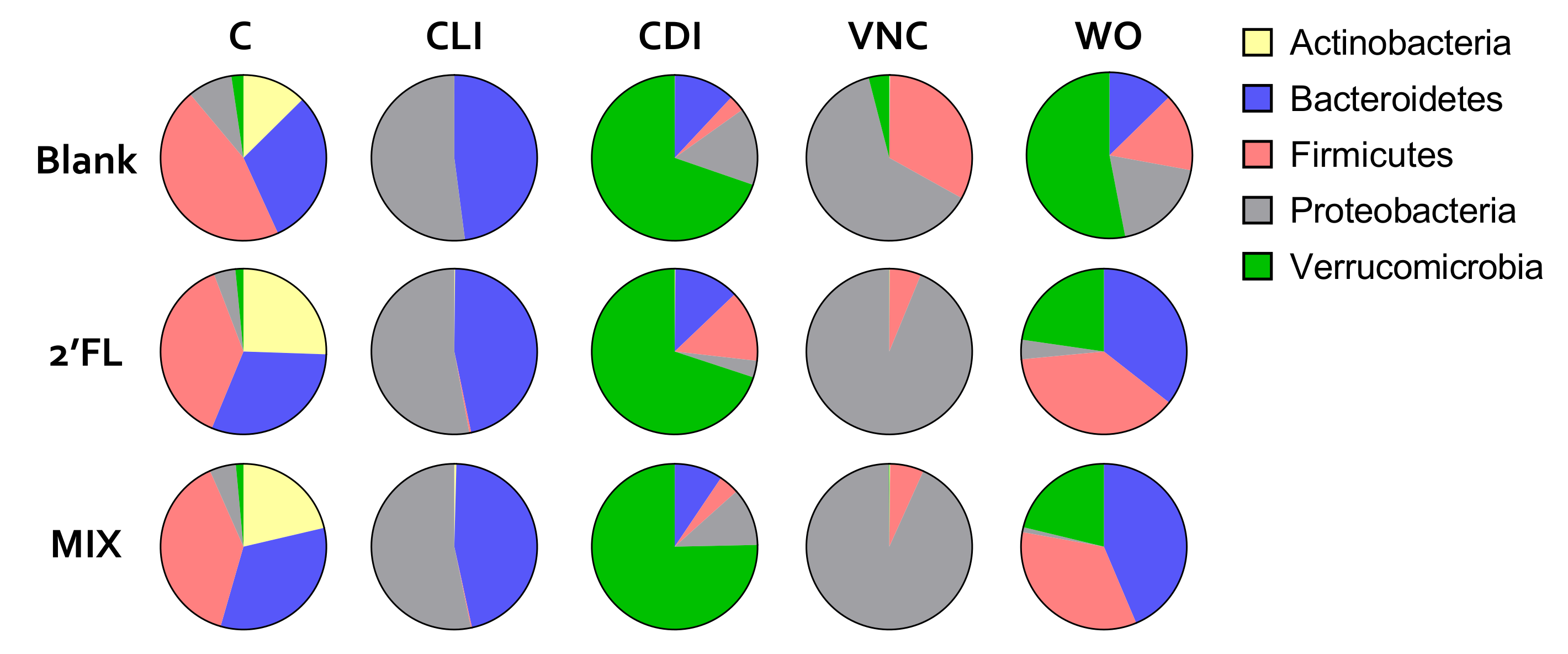
2.2.1. Reproducible Model Operation during the Control, Clindamycin, and CDI Periods
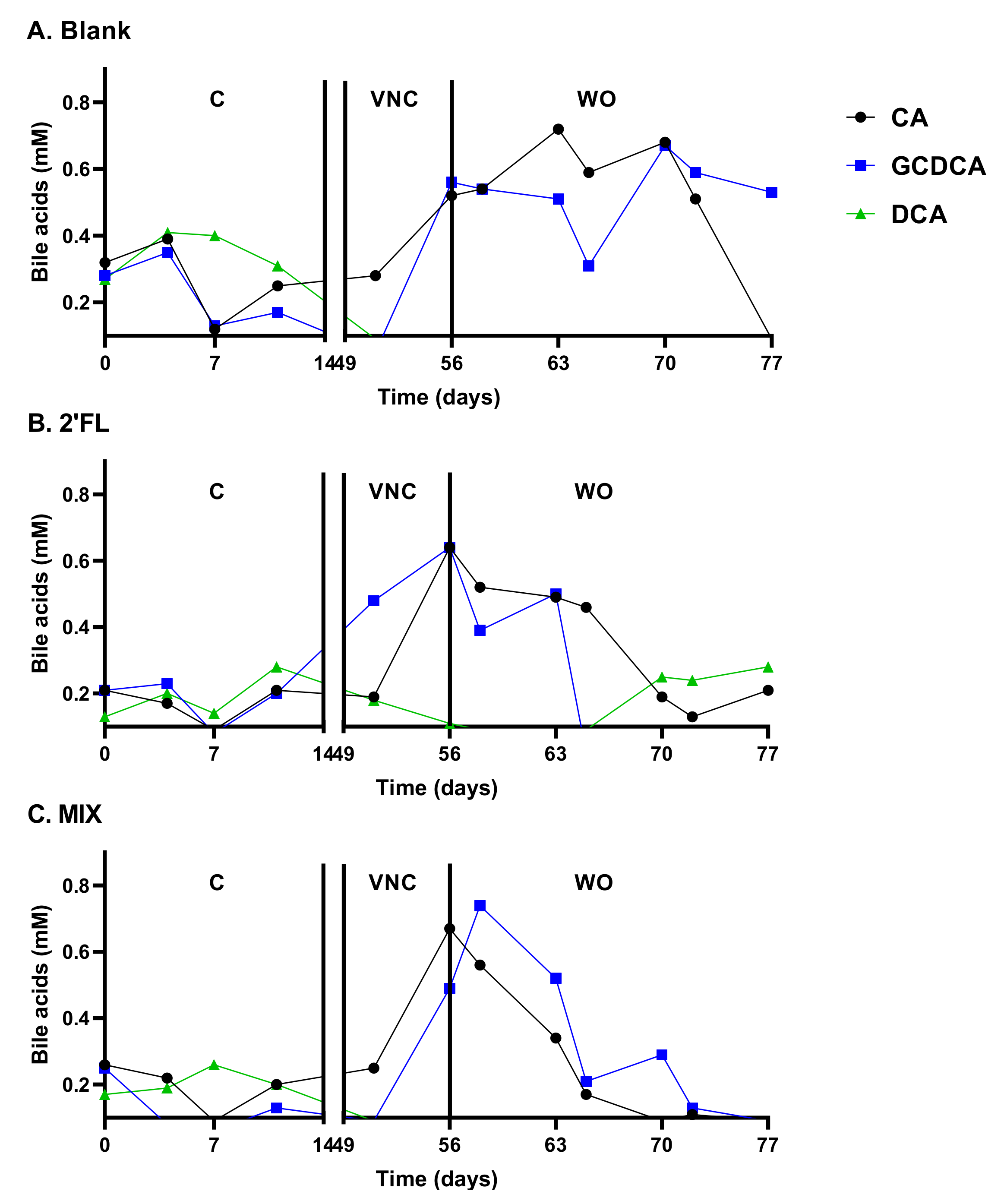
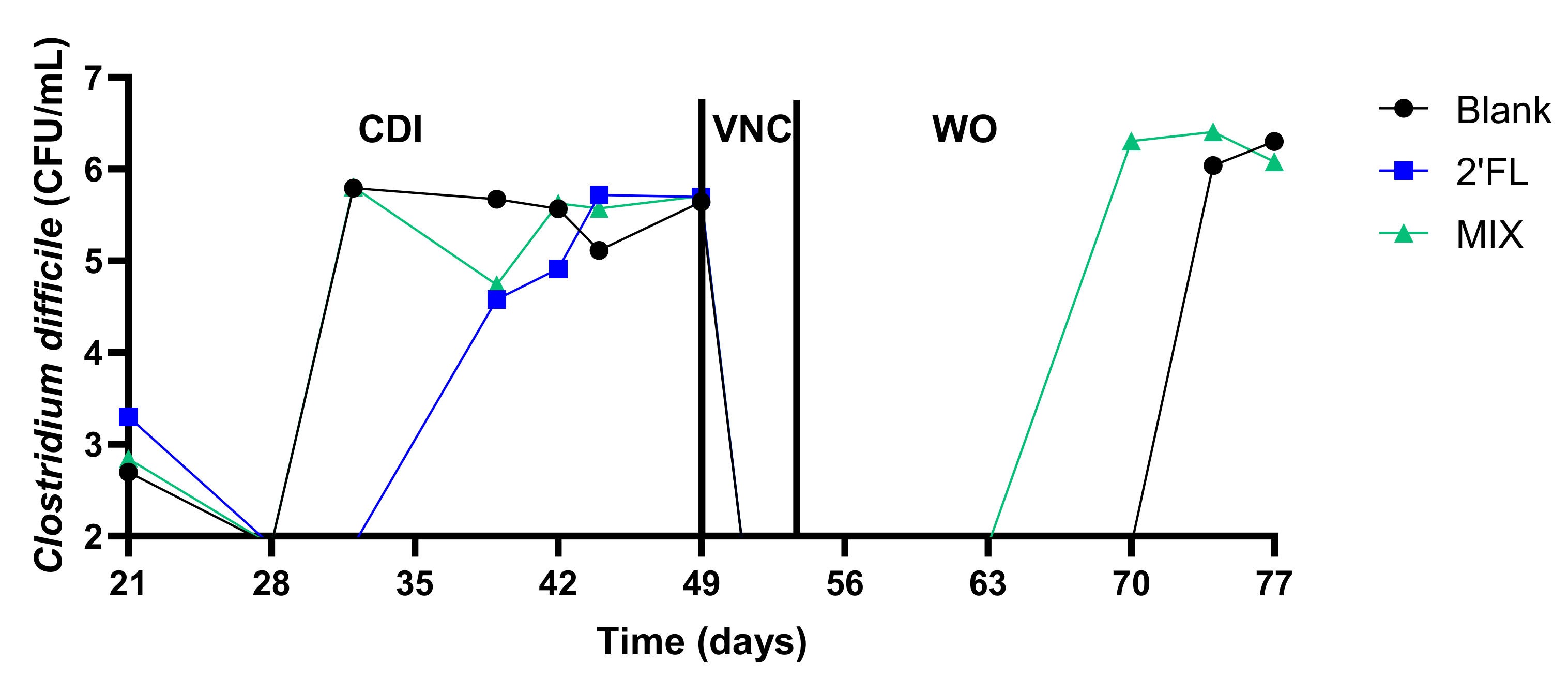
2.2.2. Vancomycin Treatment (VNC: 49–54 Days)
2.2.3. Washout Period (WO: 54–77 Days)
- Adverse effect of CLI on Bifidobacteriaceae, Firmicutes and Verrucomicrobia;
- Adverse effect of VNC on mostly Bacteroidetes and Verrucomicrobia;
- Bloom of Enterobacteriaceae during CLI and VNC treatment;
- Bloom of Verrucomicrobia after CLI and VNC treatment;
- Stimulation of Firmicutes, Bacteroidetes and Verrucomicrobia by 2′FL and MIX.
3. Discussion
4. Materials and Methods
4.1. Chemicals (General + HMOs)
4.2. Single Strain and Growth Conditions
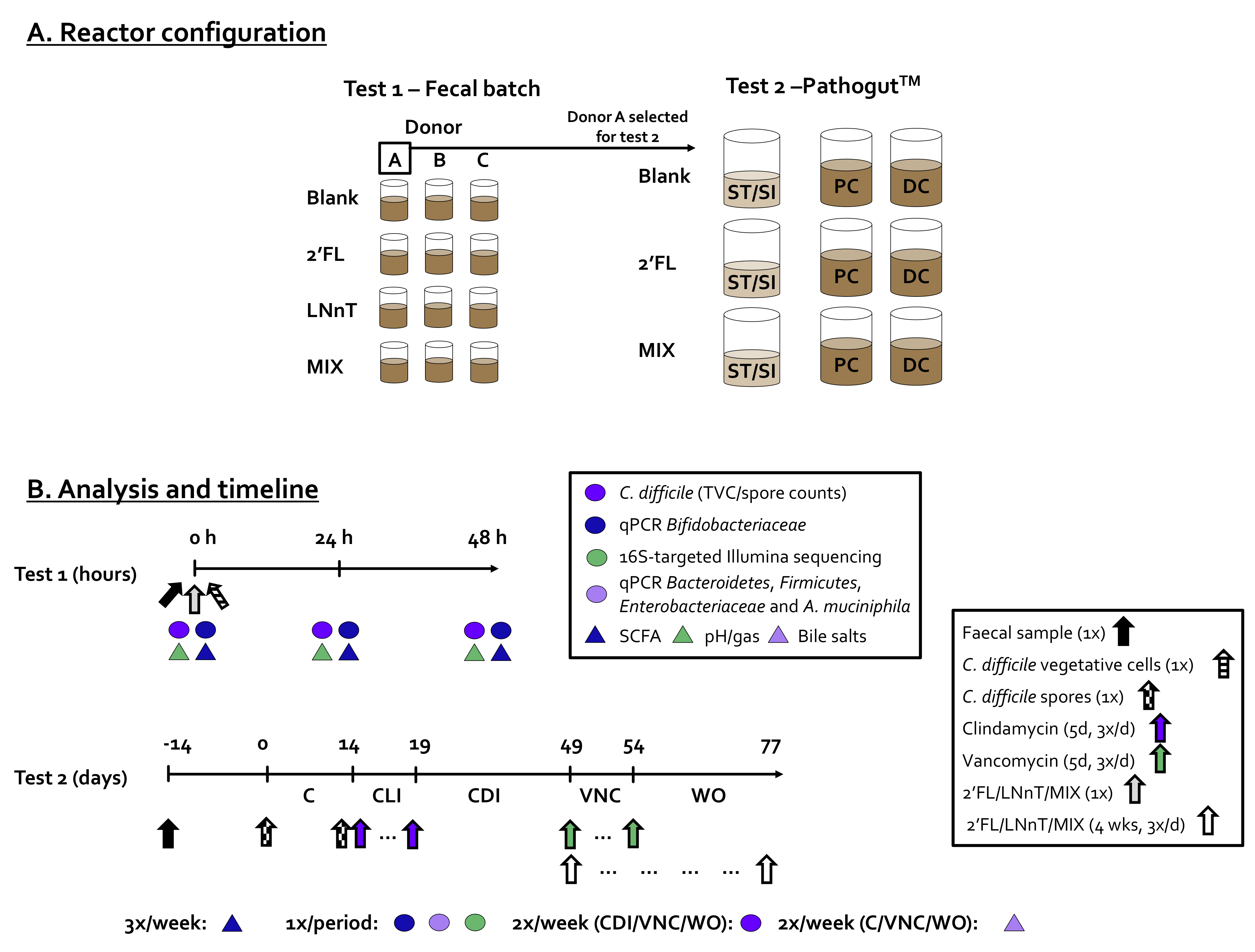
4.3. Incubation Strategies: Fecal Batch Incubation (Test 1) and PathogutTM Model (Test 2)
4.4. Activity Analysis
4.5. Compositional Analysis
4.6. Statistics
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lawson, P.A.; Citron, D.M.; Tyrrell, K.L.; Finegold, S.M. Reclassification of Clostridium Difficile as Clostridioides Difficile (Hall and O’Toole 1935) Prévot 1938. Anaerobe 2016, 40, 95–99. [Google Scholar] [CrossRef]
- Martin, J.S.H.; Monaghan, T.M.; Wilcox, M.H. Clostridium Difficile Infection: Epidemiology, Diagnosis and Understanding Transmission. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Rupnik, M.; Wilcox, M.H.; Gerding, D.N. Clostridium Difficile Infection: New Developments in Epidemiology and Pathogenesis. Nat. Rev. Microbiol. 2009, 7, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.Y.; Antonopoulos, D.A.; Kalra, A.; Tonelli, A.; Khalife, W.T.; Schmidt, T.M.; Young, V.B. Decreased Diversity of the Fecal Microbiome in Recurrent Clostridium Difficile-Associated Diarrhea. J. Infect. Dis. 2008, 197, 435–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, G.L.; Ogunsola, F.T.; Brazier, J.S.; Duerden, B.I. Modification of a PCR Ribotyping Method for Application as a Routine Typing Scheme ForClostridium Difficile. Anaerobe 1996, 2, 205–209. [Google Scholar] [CrossRef]
- Kelly, C.P. Can We Identify Patients at High Risk of Recurrent Clostridium Difficile Infection? Clin. Microbiol. Infect. 2012, 18, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Gough, E.; Shaikh, H.; Manges, A.R. Systematic Review of Intestinal Microbiota Transplantation (Fecal Bacteriotherapy) for Recurrent Clostridium Difficile Infection. Clin. Infect. Dis. 2011, 53, 994–1002. [Google Scholar] [CrossRef] [Green Version]
- Buffie, C.G.; Bucci, V.; Stein, R.R.; McKenney, P.T.; Ling, L.; Gobourne, A.; No, D.; Liu, H.; Kinnebrew, M.; Viale, A.; et al. Precision Microbiome Restoration of Bile Acid-Mediated Resistance to Clostridium Difficile. Nature 2015, 517, 205–208. [Google Scholar] [CrossRef] [Green Version]
- Abutaleb, N.S.; Seleem, M.N. In Vivo Efficacy of Auranofin in a Hamster Model of Clostridioides Difficile Infection. Sci. Rep. 2021, 11, 7093. [Google Scholar] [CrossRef]
- Borriello, S.P.; Barclay, F.E. An In-Vitro Model of Colonisation Resistance to Clostridium Difficile Infection. J. Med. Microbiol. 1986, 21, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, J.; Baines, S.D.; Jabes, D.; Wilcox, M.H. Comparison of the Efficacy of Ramoplanin and Vancomycin in Both in Vitro and in Vivo Models of Clindamycin-Induced Clostridium Difficile Infection. J. Antimicrob. Chemother. 2005, 56, 717–725. [Google Scholar] [CrossRef]
- Freeman, J.; Baines, S.D.; Saxton, K.; Wilcox, M.H. Effect of Metronidazole on Growth and Toxin Production by Epidemic Clostridium Difficile PCR Ribotypes 001 and 027 in a Human Gut Model. J. Antimicrob. Chemother. 2007, 60, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, J.; O’Neill, F.J.; Wilcox, M.H. Effects of Cefotaxime and Desacetylcefotaxime upon Clostridium Difficile Proliferation and Toxin Production in a Triple-Stage Chemostat Model of the Human Gut. J. Antimicrob. Chemother. 2003, 52, 96–102. [Google Scholar] [CrossRef]
- Goudarzi, M.; Goudarzi, H.; Alebouyeh, M.; Azimi Rad, M.; Shayegan Mehr, F.S.; Zali, M.R.; Aslani, M.M. Antimicrobial Susceptibility of Clostridium Difficile Clinical Isolates in Iran. Iran. Red. Crescent. Med. J. 2013, 15, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Totten, S.M.; Wu, L.D.; Parker, E.A.; Davis, J.C.C.; Hua, S.; Stroble, C.; Ruhaak, L.R.; Smilowitz, J.T.; German, J.B.; Lebrilla, C.B. Rapid-Throughput Glycomics Applied to Human Milk Oligosaccharide Profiling for Large Human Studies. Anal. Bioanal. Chem. 2014, 406, 7925–7935. [Google Scholar] [CrossRef] [PubMed]
- Chen, X. Human Milk Oligosaccharides (HMOS): Structure, Function, and Enzyme-Catalyzed Synthesis. Adv. Carbohydr. Chem. Biochem. 2015, 72, 113–190. [Google Scholar] [CrossRef]
- Bode, L. Human Milk Oligosaccharides: Prebiotics and Beyond. Nutr. Rev. 2009, 67 (Suppl. 2), S183–S191. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [Green Version]
- Elison, E.; Vigsnaes, L.K.; Rindom Krogsgaard, L.; Rasmussen, J.; Sørensen, N.; McConnell, B.; Hennet, T.; Sommer, M.O.A.; Bytzer, P. Oral Supplementation of Healthy Adults with 2′-O-Fucosyllactose and Lacto-N-Neotetraose Is Well Tolerated and Shifts the Intestinal Microbiota. Br. J. Nutr. 2016, 116, 1356–1368. [Google Scholar] [CrossRef] [Green Version]
- Iribarren, C.; Törnblom, H.; Aziz, I.; Magnusson, M.K.; Sundin, J.; Vigsnaes, L.K.; Amundsen, I.D.; McConnell, B.; Seitzberg, D.; Öhman, L.; et al. Human Milk Oligosaccharide Supplementation in Irritable Bowel Syndrome Patients: A Parallel, Randomized, Double-Blind, Placebo-Controlled Study. Neurogastroenterol. Motil. 2020, 32, e13920. [Google Scholar] [CrossRef] [PubMed]
- Palsson, O.S.; Peery, A.; Seitzberg, D.; Amundsen, I.D.; McConnell, B.; Simrén, M. Human Milk Oligosaccharides Support Normal Bowel Function and Improve Symptoms of Irritable Bowel Syndrome: A Multicenter, Open-Label Trial. Clin. Transl. Gastroenterol. 2020, 11, e00276. [Google Scholar] [CrossRef] [PubMed]
- Šuligoj, T.; Vigsnæs, L.K.; den Abbeele, P.V.; Apostolou, A.; Karalis, K.; Savva, G.M.; McConnell, B.; Juge, N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients 2020, 12, 2808. [Google Scholar] [CrossRef] [PubMed]
- Chichlowski, M.; De Lartigue, G.; German, J.B.; Raybould, H.E.; Mills, D.A. Bifidobacteria Isolated from Infants and Cultured on Human Milk Oligosaccharides Affect Intestinal Epithelial Function. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, S.M.; Comstock, S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016, 69 (Suppl. 2), 42–51. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.; Elderman, M.; Cheng, L.; de Haan, B.; Nauta, A.; de Vos, P. Modulation of Intestinal Epithelial Glycocalyx Development by Human Milk Oligosaccharides and Non-Digestible Carbohydrates. Mol. Nutr. Food Res. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Facinelli, B.; Marini, E.; Magi, G.; Zampini, L.; Santoro, L.; Catassi, C.; Monachesi, C.; Gabrielli, O.; Coppa, G.V. Breast Milk Oligosaccharides: Effects of 2’-Fucosyllactose and 6’-Sialyllactose on the Adhesion of Escherichia Coli and Salmonella Fyris to Caco-2 Cells. J. Matern Fetal. Neonatal Med. 2019, 32, 2950–2952. [Google Scholar] [CrossRef]
- Laucirica, D.R.; Triantis, V.; Schoemaker, R.; Estes, M.K.; Ramani, S. Milk Oligosaccharides Inhibit Human Rotavirus Infectivity in MA104 Cells. J. Nutr. 2017, 147, 1709–1714. [Google Scholar] [CrossRef] [Green Version]
- Triantis, V.; Bode, L.; van Neerven, R.J.J. Immunological Effects of Human Milk Oligosaccharides. Front. Pediatr. 2018, 6, 190. [Google Scholar] [CrossRef]
- Bode, L. The Functional Biology of Human Milk Oligosaccharides. Early Hum. Dev. 2015, 91, 619–622. [Google Scholar] [CrossRef]
- Asadpoor, M.; Peeters, C.; Henricks, P.A.J.; Varasteh, S.; Pieters, R.J.; Folkerts, G.; Braber, S. Anti-Pathogenic Functions of Non-Digestible Oligosaccharides In Vitro. Nutrients 2020, 12, 1789. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Grootaert, C.; Marzorati, M.; Possemiers, S.; Verstraete, W.; Gérard, P.; Rabot, S.; Bruneau, A.; El Aidy, S.; Derrien, M.; et al. Microbial Community Development in a Dynamic Gut Model Is Reproducible, Colon Region Specific, and Selective for Bacteroidetes and Clostridium Cluster IX. Appl. Environ. Microbiol. 2010, 76, 5237–5246. [Google Scholar] [CrossRef] [Green Version]
- Van den Abbeele, P.; Van de Wiele, T.; Verstraete, W.; Possemiers, S. The Host Selects Mucosal and Luminal Associations of Coevolved Gut Microorganisms: A Novel Concept. FEMS Microbiol. Rev. 2011, 35, 681–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orrhage, K.; Brismar, B.; Nord, C.E. Effect of Supplements with Bifidobacterium longum and Lactobacillus acidophilus on the Intestinal Microbiota during Administration of Clindamycin. Microb. Ecol. Health Dis. 1994, 7, 17–25. [Google Scholar] [CrossRef]
- De Vuyst, L.; Moens, F.; Selak, M.; Rivière, A.; Leroy, F. Summer Meeting 2013: Growth and Physiology of Bifidobacteria. J. Appl. Microbiol. 2014, 116, 477–491. [Google Scholar] [CrossRef]
- Valdés-Varela, L.; Hernández-Barranco, A.M.; Ruas-Madiedo, P.; Gueimonde, M. Effect of Bifidobacterium upon Clostridium Difficile Growth and Toxicity When Co-Cultured in Different Prebiotic Substrates. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Yang, J.; Yang, H. Antibacterial Activity of Bifidobacterium Breve Against Clostridioides Difficile. Front. Cell Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Yang, F.; Wu, Q.; Gao, J.; Liu, W.; Liu, C.; Guo, X.; Suwal, S.; Kou, Y.; Zhang, B.; et al. Protective Effects of Bifidobacterial Strains Against Toxigenic Clostridium Difficile. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Mavromatis, K.; Stackebrandt, E.; Munk, C.; Lapidus, A.; Nolan, M.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.-F.; Tapia, R.; et al. Complete Genome Sequence of the Bile-Resistant Pigment-Producing Anaerobe Alistipes Finegoldii Type Strain (AHN2437T). Stand. Genom. Sci. 2013, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Connors, J.; Dunn, K.A.; Allott, J.; Bandsma, R.; Rashid, M.; Otley, A.R.; Bielawski, J.P.; Van Limbergen, J. The Relationship between Fecal Bile Acids and Microbiome Community Structure in Pediatric Crohn’s Disease. ISME J. 2020, 14, 702–713. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Yakubov, S.; Tin, K.; Zea, D.; Garankina, O.; Ghitan, M.; Chapnick, E.; Homel, P.; Lin, Y.; Koegel, M. Does Alkaline Colonic PH Predispose to Clostridium Difficile Infection? South. Med. J. 2016, 109, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Antharam, V.C.; Li, E.C.; Ishmael, A.; Sharma, A.; Mai, V.; Rand, K.H.; Wang, G.P. Intestinal Dysbiosis and Depletion of Butyrogenic Bacteria in Clostridium Difficile Infection and Nosocomial Diarrhea. J. Clin. Microbiol. 2013, 51, 2884–2892. [Google Scholar] [CrossRef] [Green Version]
- Ng, K.M.; Aranda-Diaz, A.; Tropini, C.; Frankel, M.R.; Treuren, W.W.V.; O’Laughlin, C.; Merrill, B.D.; Yu, F.B.; Pruss, K.M.; Oliveira, R.A.; et al. Recovery of the Gut Microbiota after Antibiotics Depends on Host Diet and Environmental Reservoirs. bioRxiv 2019, 717686. [Google Scholar] [CrossRef]
- Van Herreweghen, F.; Van den Abbeele, P.; De Mulder, T.; De Weirdt, R.; Geirnaert, A.; Hernandez-Sanabria, E.; Vilchez-Vargas, R.; Jauregui, R.; Pieper, D.H.; Belzer, C.; et al. In vitro colonisation of the distal colon by Akkermansia muciniphila is largely mucin and pH dependent. Benef. Microbes 2017, 8, 81–96. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, S.D.; Boissinot, M.; Clairoux, N.; Picard François, J.; Bergeron, M.G. Rapid Detection of Clostridium Difficile in Feces by Real-Time PCR. J. Clin. Microbiol. 2003, 41, 730–734. [Google Scholar] [CrossRef] [Green Version]
- Boon, N.; Top, E.M.; Verstraete, W.; Siciliano, S.D. Bioaugmentation as a Tool To Protect the Structure and Function of an Activated-Sludge Microbial Community against a 3-Chloroaniline Shock Load. Appl. Environ. Microbiol. 2003, 69, 1511–1520. [Google Scholar] [CrossRef] [Green Version]
- Duysburgh, C.; Van den Abbeele, P.; Krishnan, K.; Bayne, T.F.; Marzorati, M. A Synbiotic Concept Containing Spore-Forming Bacillus Strains and a Prebiotic Fiber Blend Consistently Enhanced Metabolic Activity by Modulation of the Gut Microbiome in Vitro. Int. J. Pharm. 2019, 1, 100021. [Google Scholar] [CrossRef] [PubMed]
- Kok, R.G.; de Waal, A.; Schut, F.; Welling, G.W.; Weenk, G.; Hellingwerf, K.J. Specific Detection and Analysis of a Probiotic Bifidobacterium Strain in Infant Feces. Appl. Environ. Microbiol. 1996, 62, 3668–3672. [Google Scholar] [CrossRef] [Green Version]
- Edwards, A.N.; McBride, S.M. Isolating and Purifying Clostridium Difficile Spores. Methods Mol. Biol. 2016, 1476, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Moens, F.; Duysburgh, C.; van den Abbeele, P.; Morera, M.; Marzorati, M. Lactobacillus Rhamnosus GG and Saccharomyces Cerevisiae Boulardii Exert Synergistic Antipathogenic Activity in Vitro against Enterotoxigenic Escherichia coli. Benef. Microbes 2019, 10, 923–935. [Google Scholar] [CrossRef] [Green Version]
- Van den Abbeele, P.; Duysburgh, C.; Ghyselinck, J.; Goltz, S.; Berezhnaya, Y.; Boileau, T.; De Blaiser, A.; Marzorati, M. Fructans with Varying Degree of Polymerization Enhance the Selective Growth of Bifidobacterium Animalis Subsp. Lactis BB-12 in the Human Gut Microbiome In Vitro. Appl. Sci. 2021, 11, 598. [Google Scholar] [CrossRef]
- Molly, K.; Woestyne, M.V.; Smet, I.D.; Verstraete, W. Validation of the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) Reactor Using Microorganism-Associated Activities. Microb. Ecol. Health Dis. 1994, 7, 191–200. [Google Scholar] [CrossRef]
- Moens, F.; Van den Abbeele, P.; Basit, A.W.; Dodoo, C.; Chatterjee, R.; Smith, B.; Gaisford, S. A Four-Strain Probiotic Exerts Positive Immunomodulatory Effects by Enhancing Colonic Butyrate Production in Vitro. Int. J. Pharm. 2019, 555, 1–10. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a Three-Stage Compound Continuous Culture System for Investigating the Effect of Retention Time on the Ecology and Metabolism of Bacteria in the Human Colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef]
- Kunz, C.; Meyer, C.; Collado, M.C.; Geiger, L.; García-Mantrana, I.; Bertua-Ríos, B.; Martínez-Costa, C.; Borsch, C.; Rudloff, S. Influence of Gestational Age, Secretor, and Lewis Blood Group Status on the Oligosaccharide Content of Human Milk. J. Pediatr. Gastroenterol. Nutr. 2017, 64, 789–798. [Google Scholar] [CrossRef]
- De Weirdt, R.; Possemiers, S.; Vermeulen, G.; Moerdijk-Poortvliet, T.C.W.; Boschker, H.T.S.; Verstraete, W.; Van de Wiele, T. Human Faecal Microbiota Display Variable Patterns of Glycerol Metabolism. FEMS Microbiol. Ecol. 2010, 74, 601–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.L.; Chen, H.; Ouyang, W.; Metz, T.; Prakash, S. Method for Bile Acid Determination by High Performance Liquid Chromatography. J. Med. Sci. Tapei 2003, 23, 277–280. [Google Scholar]
- Rinttilä, T.; Kassinen, A.; Malinen, E.; Krogius, L.; Palva, A. Development of an Extensive Set of 16S RDNA-Targeted Primers for Quantification of Pathogenic and Indigenous Bacteria in Faecal Samples by Real-Time PCR. J. Appl. Microbiol. 2004, 97, 1166–1177. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Derrien, M.; Isolauri, E.; de Vos, W.M.; Salminen, S. Intestinal Integrity and Akkermansia Muciniphila, a Mucin-Degrading Member of the Intestinal Microbiota Present in Infants, Adults, and the Elderly. Appl. Environ. Microbiol. 2007, 73, 7767–7770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a Real-Time PCR Method for Firmicutes and Bacteroidetes in Faeces and Its Application to Quantify Intestinal Population of Obese and Lean Pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef]
- Nakano, S.; Kobayashi, T.; Funabiki, K.; Matsumura, A.; Nagao, Y.; Yamada, T. Development of a PCR Assay for Detection of Enterobacteriaceae in Foods. J. Food Prot. 2003, 66, 1798–1804. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Verstrepen, L.; Ghyselinck, J.; Albers, R.; Marzorati, M.; Mercenier, A. A Novel Non-Digestible, Carrot-Derived Polysaccharide (CRG-I) Selectively Modulates the Human Gut Microbiota While Promoting Gut Barrier Integrity: An Integrated in Vitro Approach. Nutrients 2020, 12, 1917. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, D.K. What Is the Proper Way to Apply the Multiple Comparison Test? Korean J. Anesth. 2018, 71, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigsnaes, L.K.; Ghyselinck, J.; Van den Abbeele, P.; McConnell, B.; Moens, F.; Marzorati, M.; Bajic, D. 2′FL and LNnT Exert Antipathogenic Effects against C. difficile ATCC 9689 In Vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids. Pathogens 2021, 10, 927. https://doi.org/10.3390/pathogens10080927
Vigsnaes LK, Ghyselinck J, Van den Abbeele P, McConnell B, Moens F, Marzorati M, Bajic D. 2′FL and LNnT Exert Antipathogenic Effects against C. difficile ATCC 9689 In Vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids. Pathogens. 2021; 10(8):927. https://doi.org/10.3390/pathogens10080927
Chicago/Turabian StyleVigsnaes, Louise Kristine, Jonas Ghyselinck, Pieter Van den Abbeele, Bruce McConnell, Frédéric Moens, Massimo Marzorati, and Danica Bajic. 2021. "2′FL and LNnT Exert Antipathogenic Effects against C. difficile ATCC 9689 In Vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids" Pathogens 10, no. 8: 927. https://doi.org/10.3390/pathogens10080927
APA StyleVigsnaes, L. K., Ghyselinck, J., Van den Abbeele, P., McConnell, B., Moens, F., Marzorati, M., & Bajic, D. (2021). 2′FL and LNnT Exert Antipathogenic Effects against C. difficile ATCC 9689 In Vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids. Pathogens, 10(8), 927. https://doi.org/10.3390/pathogens10080927