
Comprehensive Review of Fungi on Coffee

 ,
,  ,
,  ,
,  , , and
, , and 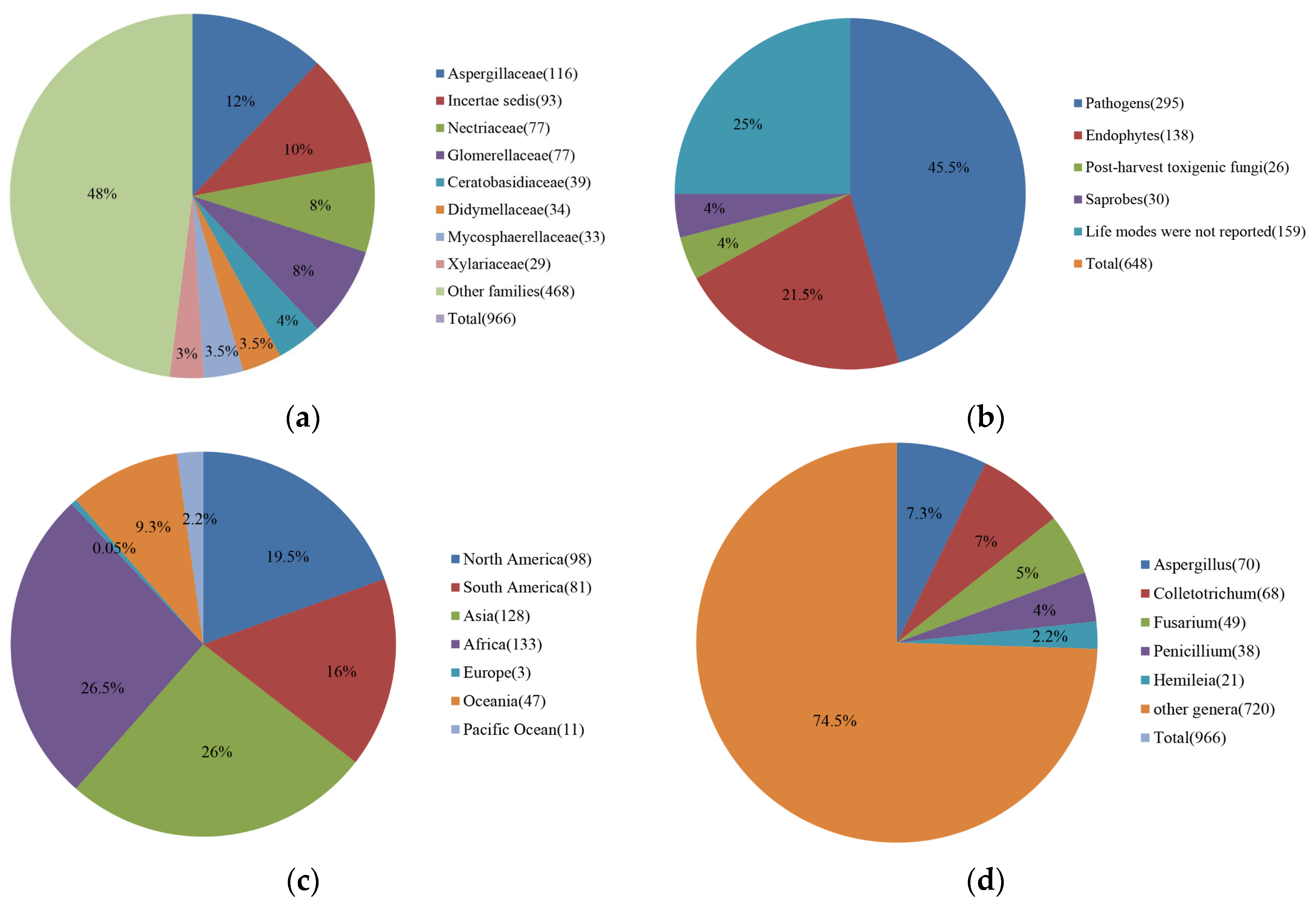{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Records of Coffee Fungi
2.2. Endophyte Role in Coffee Plants
2.3. Pathogen Effect on Coffee and Coffee Disease
2.3.1. Detection and Identification of Diseases
2.3.2. Coffee Leaf Rust
- Hosts: C. arabica (arabica coffee) and C. canephora (robusta coffee), the two most important commercial coffee species [56].
- Symptoms and signs: Infection occurs on the leaves of coffee. The first observable symptoms are small, and light-yellow spots on the upper surface of leaves. As the diameter of these points gradually increases, a large number of orange urediniospores (=uredospores) appear under the leaf surface. The fungus forms spores through stomata instead of penetrating the epidermis like most rust-causing species, so it does not form many typical rust pustules. Powdery lesions under leaves appear orange-yellow to red-orange with a high degree of variance. Although disease spots can develop anywhere on the leaf, they tend to concentrate around the edges, where dew and raindrops gather. The center of the spot eventually dries out and turns brown, while edges of the lesions continue to expand and produce new spores. At the beginning of the season, the disease usually first appears on the lower branches, and infection progresses slowly up the tree. Infected leaves fall prematurely, leaving long branches without leaves [14,56,57,58,59].
- Pathogen biology: Hemileia vastatrix mainly exists in the form of dikaryotic, and nutrient-absorbing mycelium between cells in leaves of its coffee host. Short pedicels are clustered throughout stomata and below leaves, with dual-nucleated spores. Towards the end of the season, sometimes under cool, dry conditions, spores are produced from polyspores on older attached leaves. After nuclear division and meiosis, these sporozoites germinate to produce basidia, each of which forms four haploid sporozoites [14,56,57,58,59].
- Disease cycle: Urediniosporic life cycle as its most important source of inoculum, can cause infection and develop into lesions, producing more urediniospores. Spore adhesion to the host surface, germination of urediniospores, formation of an adhesion layer on stomata, penetration, and intercellular and intracellular colonization are various steps of the disease cycle. The disease cycle of Coffee Leaf Rust is discussed in Talhinhas et al. [22].
- Disease management:
- Cultural practices: Agroforestry practices of tree-crop mixing, timely pruning, handling and de-suckering, regular change of crop cycle [49].
- Biological practices: Pichia membranifaciens is a yeast strain isolated from soil that can reduce the Hemileia vastatrix spore viability [59].
- Resistant varieties: Such as HDT (Hıbrido de Timor), Catimor and Sarchimor populations [22].
2.3.3. Coffee Berry Disease
- Pathogen: Colletotrichum kahawae is a particularly devastating pathogen that affects developing berries, leading to berry rot and shedding before bean formation. Colletotrichum kahawae has not been reported outside Africa or in low altitudes. Coffee Berry Disease was first detected and identified by McDonald in Kenya in 1922 [14].
- Symptoms and signs: Characteristic symptoms are progressive anthracnose of young and expanding coffee berries. Symptoms present as small water-soaked lesions that rapidly become dark and sunken. These lesions expand, causing rot of the entire berry under humid conditions, and pink spore masses become visible on the lesion surface. Berries are often shed from branches at an early stage of the disease. Lesions may also occur on young berry stalks, causing them to shed before lesions are evident on berries. Pale, corky lesions (scab lesions) also appear on young and mature berries that are resistant to infection. They may completely heal or remain dormant until berries ripen. This disease also affects ripening berries, causing a ‘brown blight’ phase as typical dark, sunken anthracnose lesions that envelop the red berries. Colletotrichum kahawae may also infect flowers under wet conditions, causing brown lesions on petals [14,58,60].
- Pathogen biology: The fungus settles in the mature bark of coffee buds and infects flowers, mature fruits, and leaves. Under high humidity and high temperature, conidia germinate and form germ tubes and appendages when contacted with susceptible tissues [61].
- Disease cycle: The Coffee Berry Disease cycle begins each year at the first rain event and is subsequently maintained by rain-splash dispersal and secondary inoculation of conidia onto healthy berries in the rainy season. The disease cycle of Coffee Berry Disease is discussed in De Silva et al. [62].
- Disease management:
- Cultural practices: Shading with fruit trees and irrigation to induce early flowering to decrease the severity and all berries should be removed at the end of the planting season to prevent them from becoming a source of inoculation for new crops [60].
- Biological control: Many components in the microbiota (fungi and bacteria) on coffee trees show very high antagonistic levels and have a strong antagonistic effect on Colletotrichum kahawae. However, these agents have not been developed into commercial biocontrol agents [60].
2.3.4. Coffee Wilt Disease
- Symptoms and signs: First, leaves turn yellow before withering and developing brown necrotic lesions. Finally, leaves curl, dry, and fall. This process can start from any part of the plant, but eventually, symptoms spread to the rest of the plant. Symptoms first present on the coffee stem, where fungi colonize, and the host response blocks vascular bundles, resulting in blue-black stains [14,47,58,63].
- Pathogen biology: Conidia and ascospores are spread by wind, rain and through human activities (harvesting, pruning). Pathogens penetrate wounds, so any agent causing wounds aids the spread of the fungus. Krantz and Mogk in 1973 noted that most dying and dead trees had been wounded during weeding. Insects may also spread the disease from one tree to another tree [64].
- Disease cycle: Incubation period from first symptoms to death of tree varies, although most affected trees die 2–3 months after initial symptoms were observed. It usually quickly kills infected mature trees within just 6 months after the first external symptoms appear, resulting in a decline of total yield. Coffee quality may also be affected by premature berry ripening. The disease cycle of Coffee Wilt Disease is discussed in Alemu et al. [65].
- Disease management:
- Cultural practices: Frequent inspection, along with burning infected material and spraying soil surfaces with 2.5% copper (II) sulphate. Replanting should not be done until 6 months after uprooting infected trees to allow the viability of soil inoculum to decline. It is recommended to grow cover crops such as Desmodium sp. and haricot bean, which are very efficient in suppressing weeds (so reducing the need for slashing) and as legumes, promote the growth of coffee trees [63].
- Chemical control: Ridomil Gold (metalxyl 8% + Mancozeb 64%) 68% Wp 2.5 kg/ha, when disease on set, used at 7, 14, 21, 28 days. Pencase 80% WP (Mancozeb) at the rate of 2.5 kg/ha, when disease on set, used at 7, 14, 21 days [63].
- Biological control: The strain of Bacillus subtilis (AUBB20) is the most antagonistic to this disease. Tricoderma viride and Tricoderma harzianum have shown good potential in inhibiting the mycelial growth of Fusarium xylarioides, but no effective methods of biological control are currently available [63].
2.3.5. Brown Eye Spot or Cercospora Blotch
- Symptoms and signs: on the leaves, small, round to irregular spots, and brown to light brown lesions first appear. The number and size of lesions then increase before eventually the entire leaf is affected. The edge of the lesion may appear dark purple or black, and it may be encircled by a yellow halo. Severely infected leaves turn yellow and fall off; lesions on green berries are initially brown, sunken, longitudinal, irregular or oval with a gray center. Infection can occur at any stage of berry growth; on the red cherries, first, large, sunken, and blackened areas cover with silvery fungal spores. Penetration into the seeds may cause the pulp to stick to parchment paper during processing, and damage the product. Cercospora coffeicola reduces productivity and lowers the beverage quality of coffee [14,57,67].
- Pathogen biology: Wind, splashing water and human activities cause spores (conidia) to be deposited on leaves and petioles, beginning the disease cycle. Conidia germinate at moderate to warm (20–28 °C) temperatures [68].
- Disease cycle: In warm and humid periods, new infections and sporulation occur every 7 to 10 days. Pathogen easily spreads in fields via wind, rain, and irrigation water. It survives as a pathogen in weeds and infested crop fragments, where it is capable of re-infecting grown plants. The disease cycle of Cercospora Blotch is discussed in Souza et al. [69].
- Disease management:
- Biological control: No biological control measures have been developed [68].
- Cultural practices: Elimination of crop debris, weed hosts and provide 35–65% shade. In order to maintain adequate plant nutrition, nitrogen fertilizers are used. Plant only high-quality seeds, and destroy infected crops in time after the final harvest and before replanting. Select a reasonable planting density (10 ft × 10 ft for robusta while 8 ft × 8 ft for arabica). Avoid planting coffee transplants too deep in soils [68].
- Chemical control: Fungicide sprays are necessary for disease control in wet conditions, but proper fungicides, rates, and fungicide rotations such as Chlorothalonil and Chlorothalonil Mixtures, Strobilurins and Strobilurin Mixtures should be followed [68].
2.3.6. Armillaria Root Rot
- Pathogen biology: This fungus usually exists in soil, and when coffee trees grow in unfavorable conditions, it infects the tree. For example, drought, soil compaction, root injury, and nutrient deficiency may induce it to infect the coffee tree. The fungus produces filaments on the ground, and these filaments can invade healthy roots, move to the root collar, and spread throughout the trunk. The infection causes sapwood in the affected area to rot and eventually kill the tree [70].
- Disease cycle: Armillaria spreads in two ways. The first method of transmission is through airborne sexual spores, which can sometimes lead to the creation of new infection centers. The second method of spread is through the growth of pathogens from infected trees to neighboring trees via mycelial transfer at the location where the diseased roots come into contact with each other or via rhizomes that grow through the ground. Depending on the climate, stump size, and other factors, Armillaria can live up to 50 years or more. The disease cycle of Armillaria root rot is discussed in Jayawardena et al. [71].
- Disease management: The affected trees are incurable. However, if the disease is detected early enough, host trees can be preserved. It is important to prevent this disease by avoiding conditions that cause the tree’s vitality to decline. As Armillaria root rot can last for many years, avoid replanting where the diseased trees have been removed [14,58,70].
- Biological control: Use antagonistic fungi to preemptively settle or eliminate Armillaria species in coffee plants.
- Soil barrier: Creating a barrier in the soil to root and rhizomorph growth may be a practical way to limit the pathogen’s spread, and this is called trenching. This is done by digging a trench down to 1 m (about 3 ft), lining it with plastic, and backfilling.
- Chemical control: Usually after stump removal and before planting, fumigants such as chloropicrin, carbon disulphide, and methyl bromide are used in orchards to eradicate inocula from the soil.
2.4. Distribution of Pathogenic Fungi on Coffee Worldwide
2.5. Pathogenic and Toxigenic Fungi on Coffee
3. Discussion and Conclusions
4. Materials and Methods
4.1. Fungal Taxa Reported on Coffee
4.2. Tables of Coffee Fungi
4.3. Graphs of Coffee Fungi
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Besttoppers. Available online: https://besttoppers.com/top-10-widely-consumed-drinks (accessed on 16 February 2022).
- Zhang, S.J.; De Bruyn, F.; Pothakos, V.; Contreras, G.F.; Cai, Z.; Moccand, C.; Weckx, S.; De Vuyst, L. Influence of various processing parameters on the microbial community dynamics, metabolomic profiles, and cup quality during wet coffee processing. Front. Microbiol. 2019, 10, 2621. [Google Scholar] [CrossRef] [PubMed]
- Treanor, N.B.; Saunders, J. Tackling (Illegal) Deforestation in Coffee Supply Chains: What Impact Can Demand-Side Regulations Have? Forest Policy Trade and Finance Initiative Report; Forest Trends: Washington, DC, USA, 2021. [Google Scholar]
- International Coffee Organization. Available online: http://www.ico.org/coffee_prices.asp (accessed on 16 February 2022).
- Veloso, T.G.R.; Silva, M.D.C.S.D.; Cardoso, W.S.; Guarçoni, R.C.; Kasuya, M.C.M.; Pereira, L.L. Effects of environmental factors on microbiota of fruits and soil of Coffea arabica in Brazil. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Coffee Consumption by Country 2022. Available online: https://worldpopulationreview.com/country-rankings/coffee-consumption-by-country (accessed on 16 February 2022).
- Vega, F.E.; Ziska, L.H.; Simpkins, A.; Infante, F.; Davis, A.P.; Rivera, J.A.; Barnaby, J.Y.; Wolf, J. Early growth phase and caffeine content response to recent and projected increases in atmospheric carbon dioxide in coffee (Coffea arabica and C. canephora). Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitão, A.L. Occurrence of ochratoxin A in coffee: Threads and solutions—A mini-review. Beverages 2019, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Harvey, C.A.; Rakotobe, Z.L.; Rao, N.S.; Dave, R.; Razafimahatratra, H.; Rabarijohn, R.H.; Rajaofara, H.; MacKinnon, J.L. Extreme vulnerability of smallholder farmers to agricultural risks and climate change in Madagascar. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The impact of climate change and variability on coffee production: A systematic review. Clim. Chang. 2019, 156, 609–630. [Google Scholar] [CrossRef]
- Rutherford, M.A.; Phiri, N. Pests and Diseases of Coffee in Eastern Africa: A Technical and Advisory Manual; CAB International: Wallingford, UK, 2006. [Google Scholar]
- Ribeyre, F.; Avelino, J. Impact of field pests and diseases on coffee quality. In Specialty Coffee: Managing Coffee; International Plant Nutrition Institute; IPNI [Southeast Asia]: Penang, Malaysia, 2012; pp. 151–176. [Google Scholar]
- Santamaría, J.; Bayman, P. Fungal epiphytes and endophytes of coffee leaves (Coffea arabica). Microb. Ecol. 2005, 50, 1–8. [Google Scholar] [CrossRef]
- Muller, R.A.; Berry, D.; Avelino, J.; Bieysse, D. Coffee diseases. In Coffee: Growing, Processing, Sustainable Production. A Guidebook for Growers, Processors, Traders and Researchers; Germany Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2009; pp. 495–549. [Google Scholar]
- Botrel, D.A.; Laborde, M.C.F.; De Medeiros, F.H.V.; Resende, M.L.V.; Júnio, P.M.R.; Pascholati, S.F.; Gusmão, L.F.P. Saprobic fungi as biocontrol agents of halo blight (Pseudomonas syringae pv. garcae) in coffee clones. Coffee Sci. 2018, 13, 283–291. [Google Scholar] [CrossRef]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on endophytic fungi related to secondary metabolites and biotransformation applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 1–14. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.A.; Pereira, R.G.F.A.; Lira, N.D.A.; da Glória, E.M.; Chalfoun, S.M.; Batista, L.R. Fungi associated to beans infested with coffee berry borer and the risk of ochratoxin A. Food Control 2020, 113, 107204. [Google Scholar] [CrossRef]
- Huang, X.; Ren, J.; Li, P.; Feng, S.; Dong, P.; Ren, M. Potential of microbial endophytes to enhance the resistance to postharvest diseases of fruit and vegetables. J. Sci. Food Agric. 2020, 101, 1744–1757. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Vagvolgyi, C.; Toth, B. Chemical, physical and biological approaches to prevent ochratoxin induced toxicoses in humans and animals. Toxins 2010, 2, 1718–1750. [Google Scholar] [CrossRef] [PubMed]
- McCook, S. Global rust belt: Hemileia vastatrix and the ecological integration of world coffee production since 1850. J. Glob. Hist. 2006, 1, 177–195. [Google Scholar] [CrossRef] [Green Version]
- Talhinhas, P.; Batista, D.; Diniz, I.; Vieira, A.; Silva, D.N.; Loureiro, A.; Tavares, S.; Pereira, A.P.; Azinheira, H.G.; Guerra-Guimarães, L. The coffee leaf rust pathogen Hemileia vastatrix: One and a half centuries around the tropics. Mol. Plant Pathol. 2017, 18, 1039–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coates, L.; Johnson, G. Postharvest diseases of fruit and vegetables. In Plant Pathogens and Plant Diseases; Elsevier Science: Amsterdam, The Netherlands, 1997; pp. 533–548. [Google Scholar]
- Roberto, S.R.; Youssef, K.; Hashim, A.F.; Ippolito, A. Nanomaterials as alternative control means against postharvest diseases in fruit crops. Nanomaterials 2019, 9, 1752. [Google Scholar] [CrossRef]
- Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing fungi and mechanisms of phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G. The emergence of endophytic microbes and their biological promise. J. Fungi 2018, 4, 57. [Google Scholar] [CrossRef] [Green Version]
- Miller, J. Mycotoxin in Grain, Eagan Press: St. Paul, MA, USA, 1994; 326.
- Alvindia, D.G.; de Guzman, M.F. Survey of Philippine coffee beans for the presence of ochratoxigenic fungi. Mycotoxin Res. 2016, 32, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Daou, R.; Joubrane, K.; Maroun, R.G.; Khabbaz, L.R.; Ismail, A.; El Khoury, A. Mycotoxins: Factors influencing production and control strategies. AIMS Agric. Food 2021, 6, 416–447. [Google Scholar] [CrossRef]
- Proctor, R.H.; McCormick, S.P.; Kim, H.-S.; Cardoza, R.E.; Stanley, A.M.; Lindo, L.; Kelly, A.; Brown, D.W.; Lee, T.; Vaughan, M.M. Evolution of structural diversity of trichothecenes, a family of toxins produced by plant pathogenic and entomopathogenic fungi. PLoS Pathog. 2018, 14, e1006946. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Venancio, E.J.; Ono, M.A.; Fernandes, E.V.; Hirooka, E.Y.; Shimizu, C.F.; Oba, A.; Flaiban, K.K.M.C.; Itano, E.N. Effects of subcutaneous ochratoxin-A exposure on immune system of broiler chicks. Toxins 2019, 11, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in occurrence, importance, and mycotoxin control strategies: Prevention and detoxification in foods. Foods 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.F.; Schwan, R.F.; Dias, Ë.S.; Wheals, A.E. Microbial diversity during maturation and natural processing of coffee cherries of Coffea arabica in Brazil. Int. J. Food Microbiol. 2000, 60, 251–260. [Google Scholar] [CrossRef]
- Bokhari, F.M. Mycotoxins and toxigenic fungi in Arabic coffee beans in Saudi Arabia. Adv. Biol. Res. 2007, 1, 56–66. [Google Scholar]
- Rezende, E.D.F.; Borges, J.G.; Cirillo, M.; Prado, G.; Paiva, L.C.; Batista, L. Ochratoxigenic fungi associated with green coffee beans (Coffea arabica L.) in conventional and organic cultivation in Brazil. Braz. J. Microbiol. 2013, 44, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Aldina, R.F.; Indarti, S.; Wibowo, A. Pathogenicity of Nematofagous Fungus for Control of Pratylenchus Coffeae Nematodes on Coffee Plants; Springer: Cham, Germany, 2017; pp. 243–251. [Google Scholar]
- Schulz, B.; Boyle, C. What Are Endophytes? Microbial Root Endophytes; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Goates, B.J.; Mercier, J. Effect of biofumigation with volatiles from Muscodor albus on the viability of Tilletia spp. teliospores. Can. J. Microbiol. 2009, 55, 203–206. [Google Scholar] [CrossRef]
- Monteiro, M.C.P.; Alves, N.M.; De Queiroz, M.V.; Pinho, D.B.; Pereira, O.L.; De Souza, S.M.C.; Cardoso, P.G. Antimicrobial activity of endophytic fungi from coffee plants. Biosci. J. 2017, 33, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, M.C.; Thongbai, B.; Hyde, K.D.; Broenstrup, M.; Beutling, U.; Lambert, C.; Miller, A.N.; Liu, J.K.; Promputtha, I.; Stadler, M. Elucidation of the life cycle of the endophytic genus Muscodor and itEvaluation of the effects of temperature on processed coffee beans in the presence of fungi and ochratoxin As transfer to Induratia in Induratiaceae fam. nov., based on a polyphasic taxonomic approach. Fungal Divers. 2020, 101, 177–210. [Google Scholar] [CrossRef]
- de Almeida, Â.B.; Corrêa, I.P.; Furuie, J.L.; de Farias Pires, T.; do Rocio Dalzoto, P.; Pimentel, I.C. Inhibition of growth and ochratoxin-A production in Aspergillus species by fungi isolated from coffee beans. Braz. J. Microbiol. 2019, 50, 1091–1098. [Google Scholar] [CrossRef]
- Eida, M.F.; Nagaoka, T.; Wasaki, J.; Kouno, K. Evaluation of cellulolytic and hemicellulolytic abilities of fungi isolated from coffee residue and sawdust composts. Microbes Environ. 2011, 26, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prihastuti, H.; Cai, L.; Chen, H.; McKenzie, E.; Hyde, K. Characterization of Colletotrichum species associated with coffee berries in northern Thailand. Fungal Div. 2009, 39, 89–109. [Google Scholar]
- Ma, L.-J.; Van Der Does, H.C.; Borkovich, K.A.; Coleman, J.J.; Daboussi, M.-J.; Di Pietro, A.; Dufresne, M.; Freitag, M.; Grabherr, M.; Henrissat, B.; et al. Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 2010, 464, 367–373. [Google Scholar] [CrossRef]
- Peck, L.; Nowell, R.; Flood, J.; Ryan, M.; Barraclough, T. Historical genomics reveals the evolutionary mechanisms behind multiple outbreaks of the host-specific coffee wilt pathogen Fusarium xylarioides. bioRxiv 2020, 22, 1–24. [Google Scholar] [CrossRef]
- Rutherford, M.A. Current knowledge of coffee wilt disease, a major constraint to coffee production in Africa. Phytopathology 2006, 96, 663–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musoli, P.C.; Cilas, C.; Pot, D.; Nabaggala, A.; Nakendo, S.; Pande, J.; Charrier, A.; Leroy, T.; Bieysse, D. Inheritance of resistance to coffee wilt disease (Fusarium xylarioides Steyaert) in Robusta coffee (Coffea canephora Pierre) and breeding perspectives. Tree Genet. Genomes 2012, 9, 351–360. [Google Scholar] [CrossRef]
- Gichuru, E.; Alwora, G.; Gimase, J.; Kathurima, C. Coffee leaf rust (Hemileia vastatrix) in Kenya—A review. Agronomy 2021, 11, 2590. [Google Scholar] [CrossRef]
- De Andrade, C.C.L.; Vicentin, R.P.; Costa, J.R.; Perina, F.J.; De Resende, M.L.V.; Alves, E. Alterations in antioxidant metabolism in coffee leaves infected by Cercospora coffeicola. Ciênc. Rural 2016, 46, 1764–1770. [Google Scholar] [CrossRef] [Green Version]
- Otieno, W. Armillaria Root Rot of Tea in Kenya: Characterization of the Pathogen and Approaches to Disease Management; Wageningen University and Research; Wageningen University: Wageningen, The Netherlands, 2002; Available online: https://www.semanticscholar.org/paper/Armillaria-root-rot-of-tea-in-Kenya-%3A-of-the-and-to-Otieno/c0f9753b5d198b0a8eb412d40b560c58304eef02#citing-papers (accessed on 16 February 2022).
- Fang, Y.; Ramasamy, R.P. Current and prospective methods for plant disease detection. Biosensors 2015, 5, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Lima, D.; Carrión, G.; Sánchez-Nava, P.; Desgarennes, D.; Villain, L. Fungal diversity and Fusarium oxysporum pathogenicity associated with coffee corky-root disease in Mexico. Rev. Fac. Cienc. Agrar. UNCuyo 2020, 52, 276–292. [Google Scholar]
- Hariharan, G.; Prasannath, K. Recent advances in molecular diagnostics of fungal plant pathogens: A mini review. Front. Cell. Infect. Microbiol. 2021, 10. [Google Scholar] [CrossRef]
- Senanayake, I.; Rathnayaka, A.; Marasinghe, D.; Calabon, M.; Gentekaki, E.; Lee, H.; Hurdeal, V.; Pem, D.; Dissanayake, L.; Wijesinghe, S. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Arneson, P. Coffee Rust; Plant Health Instructor; The American Phytopathological Society (APS): St. Paul, MN, USA, 2000. [Google Scholar]
- Abu Mettleq, A.S.; Abu-Naser, S.S. A rule based system for the diagnosis of coffee diseases. Int. J. Acad. Inf. Syst. Res. (IJAISR) 2019, 3, 1–8. [Google Scholar]
- Hailu, B.Z. Fungal Disease Dynamics, Genetic Variation and Biodiversity-Yield Relationships: A Study Along a Gradient of Coffee Management in Southwestern Ethiopia. Ph.D. Thesis, Stockholm University, Faculty of Science, Department of Ecology, Environment and Plant Sciences, Stockholm, Sweden, 2020. [Google Scholar]
- de Resende, M.L.V.; Pozza, E.A.; Reichel, T.; Botelho, D.M.S. Strategies for coffee leaf rust management in organic crop systems. Agronomy 2021, 11, 1865. [Google Scholar] [CrossRef]
- Lemma, D.T.; Abewoy, D. Review on integrated pest management of coffee berry disease and coffee berry borer. Int. J. Plant Breed. Crop Sci. 2021, 8, 1001–1008. [Google Scholar]
- Bailey, J.A. Colletotrichum: Biology, Pathology and Control; CAB International: Wallingford, UK, 1992. [Google Scholar]
- De Silva, D.D.; Crous, P.W.; Ades, P.K.; Hyde, K.D.; Taylor, P.W.J. Life styles of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 2017, 31, 155–168. [Google Scholar] [CrossRef]
- Wassie, A.K. Integrated diseased management on coffee wilt disease caused by Fusarium xylarioides and its distribution in Ethiopian review. Agric. Res. Technol. Open Access J. 2019, 23, 302–308. [Google Scholar] [CrossRef]
- Wrigley, G. Coffee; Longman Scientific & Technical: Harlow, UK; Essex, UK, 1988. [Google Scholar]
- Alemu, T. A review of coffee wilt disease, Gibberella xylarioides (Fusarium xylarioides) in Africa with special reference to Ethiopia. Ethiop. J. Biol. Sci. 2012, 11, 65–103. [Google Scholar]
- Nelson, S.C. Cercospora Leaf Spot and Berry Blight of Coffee; Plant Disease; University of Hawaii: Mänoa, Hawaii, 2008; Available online: https://www.ctahr.hawaii.edu/oc/freepubs/pdf/PD-41.pdf (accessed on 16 February 2022).
- Vale, P.A.S.; De Resende, M.L.V.; Botelho, D.M.D.S.; De Andrade, C.C.L.; Alves, E.; Ogoshi, C.; Guimarães, S.D.S.C.; Pfenning, L.H. Epitypification of Cercospora coffeicola and its involvement with two different symptoms on coffee leaves in Brazil. Eur. J. Plant Pathol. 2020, 159, 399–408. [Google Scholar] [CrossRef]
- Schwartz, H.F.; Gent, D.H. Cercospora leaf spot (Cucumber, Melon, Pumpkin, Squash, and Zucchini). In High Plains IPM Guide; University of Wyoming: Laramie, WY, USA; University of Nebraska: Lincoln, NE, USA; Colorado State University: Fort Collins, CO, USA; Montana State University: Bozeman, MT, USA, 2007; Available online: http://wiki.bugwood.org/uploads/CercosporaLeafSpot-Cucurbits.pdf (accessed on 16 February 2022).
- Souza, A.G.C.; Rodrigues, F.; Maffia, L.A.; Mizubuti, E.S.G. Infection process of Cercospora coffeicola on coffee leaf. J. Phytopathol. 2010, 159, 6–11. [Google Scholar] [CrossRef]
- Gezahgne, A.; Coetzee, M.P.A.; Wingfield, B.D.; Wingfield, M.J.; Roux, J. Identification of the Armillaria root rot pathogen in Ethiopian plantations. For. Pathol. 2004, 34, 133–145. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Hyde, K.D.; Chen, Y.J.; Papp, V.; Palla, B.; Papp, D.; Bhunjun, C.S.; Hurdeal, V.G.; Senwanna, C.; Manawasinghe, I.S.; et al. One stop shop IV: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 76–100 (2020). Fungal Divers. 2020, 103, 87–218. [Google Scholar] [CrossRef]
- Culliao, A.G.L.; Barcelo, J.M. Fungal and mycotoxin contamination of coffee beans in Benguet province, Philippines. Food Addit. Contam. Part A 2015, 32, 250–260. [Google Scholar] [CrossRef]
- Dos Santos, D.G.; Coelho, C.C.d.S.; Ferreira, A.B.R.; Freitas-Silva, O. Brazilian coffee production and the future microbiome and mycotoxin profile considering the climate change Scenario. Microorganisms 2021, 9, 858. [Google Scholar] [CrossRef] [PubMed]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarch, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- Shuping, D.; Eloff, J. The use of plants to protect plants and food against fungal pathogens: A review. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Takundwa, M.; Ruzvidzo, O.; Uzabakiriho, J.; Titus, P.; Chimwamurombe, P. Molecular identification of fungal species associated with leaf lesions of Marama bean seedlings in Namibia. Afr. J. Microbiol. Res. 2015, 9, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Cerda, R.; Avelino, J.; Gary, C.; Tixier, P.; Lechevallier, E.; Allinne, C. Primary and secondary yield losses caused by pests and diseases: Assessment and modeling in coffee. PLoS ONE 2017, 12, e0169133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemessa, F.; Abera, A.; Adunga, G.; Garedew, W. Association of mycoflora with coffee (Coffea arabica L.) beans at limmu coffee plantation, Southwestern Ethiopia. Plant Pathol. J. 2015, 14, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Hindorf, H.; Omondi, C.O. A review of three major fungal diseases of Coffea arabica L. in the rainforests of Ethiopia and progress in breeding for resistance in Kenya. J. Adv. Res. 2011, 2, 109–120. [Google Scholar] [CrossRef] [Green Version]
- De Paula, P.V.A.A.; Pozza, E.A.; Santos, L.A.; Chaves, E.; Maciel, M.P.; Paula, J.C.A. Diagrammatic scales for assessing brown eye spot (Cercospora coffeicola) in red and yellow coffee cherries. J. Phytopathol. 2016, 164, 791–800. [Google Scholar] [CrossRef]
- Djuikem, C.; Grognard, F.; Touzeau, S.; Bowong, S. Modelling and controlling fungus Hemileia vastatrix, a coffee pest. In Proceedings of the BIOMATH 2019-International Conference on Mathematical Methods and Models in Biosciences, Bedlevo, Poland, 16–22 June 2019; pp. 16–24. [Google Scholar]
- Douglas, S.M. Leaf spot diseases of ornamental trees and shrubs. In The Connecticut Experiment Station Report; University of Minnesota Extension: Falcon Heights, MN, USA, 2012. [Google Scholar]
- Ghosh, S.K.; Chaudhary, M.; Manjunatha, N. 14 Endophytes: A potential bio-agent for the plant protection. In Innovative Pest Management Approaches for the 21st Century: Harnessing Automated Unmanned Technologies; Springer: Singapore, 2020; p. 273. [Google Scholar]
- Morelli, M.; Bahar, O.; Papadopoulou, K.K.; Hopkins, D.L.; Obradovic, A. Editorial: Role of endophytes in plant health and defense against pathogens. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Salcedo-Sarmiento, S.; Aucique-Pérez, C.E.; Silveira, P.R.; Colmán, A.A.; Silva, A.L.; Mansur, P.S.C.; Rodrigues, F.; Evans, H.C.; Barreto, R.W. Elucidating the interactions between the rust Hemileia vastatrix and a Calonectria mycoparasite and the coffee plant. iScience 2021, 24, 102352. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of Pesticides on Environment; Plant, Soil and Microbes; Springer: Berlin/Heidelberg, Germany, 2016; pp. 253–269. [Google Scholar]
- Petit, A.-N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.; Sriram, S. Biological control of Fusarium wilt in crop plants using non-pathogenic isolates of Fusarium species. Indian Phytopathol. 2020, 73, 11–19. [Google Scholar] [CrossRef]
- Naik, B.S. Biosynthesis of silver nanoparticles from endophytic fungi and their role in plant disease management. Microb. Endophytes. 2020, 16, 307–321. [Google Scholar] [CrossRef]
- Khan, M.R.; Ahamad, F.; Rizvi, T.F. Chapter 8—Effect of nanoparticles on plant pathogens. In Advances in Phytonanotechnology; Academic Press: Cambridge, MA, USA, 2019; pp. 215–240. [Google Scholar]
- Ferreira, F.V.; Herrmann-Andrade, A.M.; Calabrese, C.D.; Bello, F.; Vázquez, D.; Musumeci, M.A. Effectiveness of Trichoderma strains isolated from the rhizosphere of citrus tree to control Alternaria alternata, Colletotrichum gloeosporioides and Penicillium digitatum A21 resistant to pyrimethanil in post-harvest oranges (Citrus sinensis L.(Osbeck)). J. Appl. Microbiol. 2020, 129, 712–727. [Google Scholar] [CrossRef]
- Dowling, M.; Peres, N.; Villani, S.; Schnabel, G. Managing Colletotrichum on fruit crops: A “complex” challenge. Plant Dis. 2020, 104, 2301–2316. [Google Scholar] [CrossRef]
- Gimase, J.M.; Thagana, W.M.; Omondi, C.O.; Cheserek, J.J.; Gichimu, B.M.; Gichuru, E.K. Quantitative trait loci (QTL) mapping of resistance to coffee berry disease (Colletotrichum kahawae Waller Bridge) in Coffea arabica L. variety Rume Sudan. Afr. J. Agric. Res. 2020, 16, 1184–1194. [Google Scholar] [CrossRef]
- Silva, M.D.C.; Várzea, V.; Guerra-Guimarães, L.; Gil Azinheira, H.; Fernandez, D.; Petitot, A.-S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef] [Green Version]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The impact of climate change on indigenous Arabica coffee (Coffea arabica): Predicting future trends and identifying priorities. PLoS ONE 2012, 7, e47981. [Google Scholar] [CrossRef]
- Jackson, D.; Skillman, J.; Vandermeer, J. Indirect biological control of the coffee leaf rust, Hemileia vastatrix, by the entomogenous fungus Lecanicillium lecanii in a complex coffee agroecosystem. Biol. Control 2012, 61, 89–97. [Google Scholar] [CrossRef]
- Joshi, V.; Pandit, A.; Vaidya, D. Ochratoxin-A in food: An overview. Int. J. Food Ferment. Technol. 2017, 7, 13–24. [Google Scholar] [CrossRef]
- McQueen, C. Comprehensive Toxicology; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Santiago, W.D.; Teixeira, A.R.; de Andrade Santiago, J.; Lopes, A.C.A.; Brandão, R.M.; Barbosa, R.B.; Caetano, A.R.S.; das Graças Cardoso, M.; Resende, M.L.V. Standardization of an analytical method to quantify ochratoxin-A in green coffee beans by high performance liquid chromatography. Res. Soc. Dev. 2020, 9, e39985070. [Google Scholar] [CrossRef]
- Viegas, C.; Pacífico, C.; Faria, T.; De Oliveira, A.C.; Caetano, L.A.; Carolino, E.; Gomes, A.; Viegas, S. Fungal contamination in green coffee beans samples: A public health concern. J. Toxicol. Environ. Health Part A 2017, 80, 719–728. [Google Scholar] [CrossRef]
- Suárez-Quiroz, M.; González-Rios, O.; Barel, M.; Guyot, B.; Schorr-Galindo, S.; Guiraud, J.P. Study of ochratoxin-A producing strains in coffee processing. Int. J. Food Sci. Technol. 2004, 39, 501–507. [Google Scholar] [CrossRef]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Samson, R. An Overview of Ochratoxin Research; CABI Publishers: London, UK, 2009. [Google Scholar]
- Cinar, A.; Onbaşı, E. Mycotoxins: The hidden danger in foods. Mycotoxins Food Saf. 2020, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Romano, A.; Masi, P.; Cavella, S. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018, 98, 4003–4013. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, W.; Liu, Y.; Lin, F.; Huang, Z.; Lu, F.; Bie, X.; Lu, Z. Growth inhibition of Fusarium graminearum and reduction of deoxynivalenol production in wheat grain by bacillomycin D. J. Stored Prod. Res. 2018, 75, 21–28. [Google Scholar] [CrossRef]
- Vieira, T.; Cunha, S.; Casal, S. Mycotoxins in coffee. In Coffee in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2015; pp. 225–233. [Google Scholar]
- Gil-Serna, J.; Vázquez, C.; Sandino, F.G.; Valle, A.M.; González-Jaén, M.T.; Patiño, B. Evaluation of growth and ochratoxin A production by Aspergillus steynii and Aspergillus westerdijkiae in green-coffee based medium under different environmental conditions. Food Res. Int. 2014, 61, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Palumbo, J.D.; O’Keeffe, T.L.; Mahoney, N.E. Inhibition of ochratoxin A production and growth of Aspergillus species by phenolic antioxidant compounds. Mycopathologia 2007, 164, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Aroyeun, S.; Adegoke, G. Reduction of ochratoxin-A (OTA) in spiked cocoa powder and beverage using aqueous extracts and essential oils of Aframomum danielli. Afr. J. Biotechnol. 2007, 6, 612–616. [Google Scholar]
- Nguefack, J.; Nguikwie, S.; Fotio, D.; Dongmo, B.; Zollo, P.A.; Leth, V.; Nkengfack, A.; Poll, L. Fungicidal potential of essential oils and fractions from Cymbopogon citratus, Ocimum gratissimum and Thymus vulgaris to control Alternaria padwickii and Bipolaris oryzae, two seed borne fungi of rice (Oryza sativa L.). J. Essent. Oil Res. 2007, 19, 581–587. [Google Scholar] [CrossRef]
- Magan, N.; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Int. J. Food Microbiol. 2007, 119, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Basílico, M.Z.; Basilico, J.C. Inhibitory effects of some spice essential oils on Aspergillus ochraceus NRRL 3174 growth and ochratoxin-A production. Lett. Appl. Microbiol. 1999, 29, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-E.; Park, B.S.; Bayman, P.; Baker, J.L.; Choi, W.S.; Campbell, B.C. Suppression of ochratoxin biosynthesis by naturally occurring alkaloids. Food Addit. Contam. 2007, 24, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Kouadio, I.A.; Ban, L.K.; Dosso, M.B. Prevention of ochratoxin A (OTA) production in coffee beans using natural antifungal derived from Solanum indicum L. green berries. J. Food Secur. 2019, 7, 63–71. [Google Scholar] [CrossRef]
- Gómez, J.V.; Tarazona, A.; Mateo, F.; Jiménez, M.; Mateo, E.M. Potential impact of engineered silver nanoparticles in the control of aflatoxins, ochratoxin A and the main aflatoxigenic and ochratoxigenic species affecting foods. Food Control 2019, 101, 58–68. [Google Scholar] [CrossRef]
- Bemvenuti, R.; Rodrigues, M.H.; Furlong, E. Efficiency of γ-oryzanol against the complex Fusarium graminearum growth and mycotoxins production. Food Sci. Technol. 2019, 39, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Misra, N.N.; Yadav, B.; Roopesh, M.S.; Jo, C. Cold plasma for effective fungal and mycotoxin control in foods: Mechanisms, inactivation effects, and applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 106–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Duong, B.; Marraccini, P.; Maeght, J.-L.; Vaast, P.; Lebrun, M.; Duponnois, R. Coffee microbiota and its potential use in sustainable crop management. A review. Front. Sustain. Food Syst. 2020, 4, 237. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Available online: https://nt.ars-grin.gov/fungaldatabases/fungushost/fungushost.cfm (accessed on 16 February 2022).
- Index Fungorum. 2022. Available online: http://www.indexfungorum.org/Names/IndexFungorumPartnership.htm (accessed on 16 February 2022).
- Mycobank. 2022. Available online: https://www.mycobank.org/ (accessed on 16 February 2022).
- Noonim, P.; Mahakarnchanakul, W.; Nielsen, K.F.; Frisvad, J.C.; Samson, R.A. Isolation, identification and toxigenic potential of ochratoxin A-producing Aspergillus species from coffee beans grown in two regions of Thailand. Int. J. Food Microbiol. 2008, 128, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Estupiñan-López, L.; Campos, V.P.; da Silva Júnior, J.C.; Pedroso, M.P.; Terra, W.C.; da Silva, J.C.P.; de Paula, L.L. Volatile compounds produced by Fusarium spp. isolated from Meloidogyne paranaensis egg masses and corticous root tissues from coffee crops are toxic to Meloidogyne incognita. Trop. Plant Pathol. 2018, 43, 183–193. [Google Scholar] [CrossRef]
- Sousa, T.; Batista, L.; Passamani, F.; Lira, N.; Cardoso, M.; Santiago, W.; Chalfoun, S. Evaluation of the effects of temperature on processed coffee beans in the presence of fungi and ochratoxin A. J. Food Saf. 2019, 39, e12584. [Google Scholar] [CrossRef] [Green Version]
- Kuntawee, S.; Akarapisan, A. Isolation and identification of Aspergillus species producing Ochratoxin a in Arabica coffee beans. J. Agric. Technol. 2015, 11, 1235–1242. [Google Scholar]
- Batista, L.R.; Chalfoun, S.M.; Prado, G.; Schwan, R.F.; Wheals, A.E. Toxigenic fungi associated with processed (green) coffee beans (Coffea arabica L.). Int. J. Food Microbiol. 2003, 85, 293–300. [Google Scholar] [CrossRef]
- Djadjiti, N.; Aziz, M.A.; Esther, O.K.; Dane, S. Isolation and Identification of Fungal Pathogens Contaminating Some Coffee Powder Marketed in the City of Abuja. J. Res. Med. Dent. Sci. 2020, 8, 172–175. [Google Scholar]
- Pasin, L.A.A.P.; Almeida, J.R.d.; Abreu, M.S.d. Fungos associados a grãos de cinco cultivares de café (Coffea arabica L.). Acta. Bot. Bras. 2009, 23, 1129–1132. [Google Scholar] [CrossRef] [Green Version]
- Casas-Junco, P.P.; Ragazzo-Sánchez, J.A.; de Jesus Ascencio-Valle, F.; Calderón-Santoyo, M. Determination of potentially mycotoxigenic fungi in coffee (Coffea arabica L.) from Nayarit. Food Sci. Biotechnol. 2018, 27, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Chaves, F.C.; Gianfagna, T.J.; Aneja, M.; Posada, F.; Peterson, S.W.; Vega, F.E. Aspergillus oryzae NRRL 35191 from coffee, a non-toxigenic endophyte with the ability to synthesize kojic acid. Mycol. Prog. 2012, 11, 263–267. [Google Scholar] [CrossRef]
- González-Osorio, H.; Botero, C.E.G.; Rivera, R.D.M.; Vega, N.W.O. Screening for phosphate-solubilizing fungi from colombian andisols cultivated with coffee (Coffea arabica L.). Coffee Sci. 2020, e151666. [Google Scholar] [CrossRef]
- Hongsanan, S.; Tian, Q.; Hyde, K.; Chomnunti, P. Two new species of sooty moulds, Capnodium coffeicola and Conidiocarpus plumeriae in Capnodiaceae. Mycosp 2015, 6, 814–824. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.; Haituk, S.; Pakdeeniti, P.; Al-Sadi, A.; Hongsanan, S.; Chomnunti, P.; Cheewangkoon, R. Phaeosaccardinula coffeicola and Trichomerium chiangmaiensis, two new species of Chaetothyriales (Eurotiomycetes) from Thailand. Mycosphere 2018, 9, 769–778. [Google Scholar] [CrossRef]
- Bongiorno, V.A.; Rhoden, S.A.; Garcia, A.; Polonio, J.C.; Azevedo, J.L.; Pereira, J.O.; Pamphile, J.A. Genetic diversity of endophytic fungi from Coffea arabica cv. IAPAR-59 in organic crops. Ann. Microbiol. 2016, 66, 855–865. [Google Scholar] [CrossRef]
- Oliveira, R.; Souza, R.; Lima, T.; Cavalcanti, M. Endophytic fungal diversity in coffee leaves (Coffea arabica) cultivated using organic and conventional crop management systems. Mycosphere 2014, 5, 523–530. [Google Scholar] [CrossRef]
- Saucedo-García, A.; Anaya, A.L.; Espinosa-García, F.J.; González, M.C. Diversity and communities of foliar endophytic fungi from different agroecosystems of Coffea arabica L. in two regions of Veracruz, Mexico. PLoS ONE 2014, 9, e98454. [Google Scholar] [CrossRef]
- Fernandes, M.d.R.V.; Silva, T.A.C.; Pfenning, L.H.; Costa-Neto, C.M.d.; Heinrich, T.A.; Alencar, S.M.d.; Lima, M.A.d.; Ikegaki, M. Biological activities of the fermentation extract of the endophytic fungus Alternaria alternata isolated from Coffea arabica L. Braz. J. Pharm. Sci. 2009, 45, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Serrato-Diaz, L.M.; Mariño, Y.A.; Bayman, P. Pathogens causing Anthracnose and fruit rots of coffee associated with the coffee berry borer and the entomopathogenic fungus Beauveria bassiana in Puerto Rico. Phytopathology 2020, 110, 1541–1552. [Google Scholar] [CrossRef]
- Kenny, M.; Galea, V.; Price, T. Germination and growth of Colletotrichum acutatum and Colletotrichum gloeosporioides isolates from coffee in Papua New Guinea and their pathogenicity to coffee berries. Australas. Plant Path. 2012, 41, 519–528. [Google Scholar] [CrossRef]
- María del Carmen, H.R.; Evans, H.C.; de Abreu, L.M.; de Macedo, D.M.; Ndacnou, M.K.; Bekele, K.B.; Barreto, R.W. New species and records of Trichoderma isolated as mycoparasites and endophytes from cultivated and wild coffee in Africa. Sci. Rep. 2021, 11, 1–30. [Google Scholar]
- Gizaw, B.; Tsegaye, Z.; Tefera, G. Isolation, identification and characterization of yeast species from coffee waste collected from Sidama and Gedio zone. J. Yeast Fungal Res. 2016, 7, 47–53. [Google Scholar]
- Lu, L.; Tibpromma, S.; Karunarathna, S.; Thiyagaraja, V.; XU, J.C.; Jayawardena, R.S.; Lumyong, S.; Hyde, K.D. Taxonomic and phylogenic appraisal of a novel species and a new record of Stictidaceae from coffee in Yunnan Province, China. Phytotaxa 2021, 528, 111–124. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Tibpromma, S.; Karunarathna, S.C.; Jayawardena, R.S.; Lumyong, S.; Xu, J.; Hyde, K.D. Comprehensive Review of Fungi on Coffee. Pathogens 2022, 11, 411. https://doi.org/10.3390/pathogens11040411
Lu L, Tibpromma S, Karunarathna SC, Jayawardena RS, Lumyong S, Xu J, Hyde KD. Comprehensive Review of Fungi on Coffee. Pathogens. 2022; 11(4):411. https://doi.org/10.3390/pathogens11040411
Chicago/Turabian StyleLu, Li, Saowaluck Tibpromma, Samantha C. Karunarathna, Ruvishika S. Jayawardena, Saisamorn Lumyong, Jianchu Xu, and Kevin D. Hyde. 2022. "Comprehensive Review of Fungi on Coffee" Pathogens 11, no. 4: 411. https://doi.org/10.3390/pathogens11040411
APA StyleLu, L., Tibpromma, S., Karunarathna, S. C., Jayawardena, R. S., Lumyong, S., Xu, J., & Hyde, K. D. (2022). Comprehensive Review of Fungi on Coffee. Pathogens, 11(4), 411. https://doi.org/10.3390/pathogens11040411