

Natural Killer Repertoire Restoration in TB/HIV Co-Infected Individuals Experienced an Immune Reconstitution Syndrome (CAMELIA Trial, ANRS 12153)

 , , , and
on behalf of the CAMELIA Study Team
, , , and
on behalf of the CAMELIA Study Team {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Individuals and Methods
2.1. Individuals
2.2. Phenotypic Studies of NK Cells
2.3. Statistical Analysis
3. Results
3.1. Clinical and Laboratory Features of Enrolled HIV-Infected Individuals
3.2. Analysis of the Immune Reconstitution of Peripheral NK Cells and Their Subsets in IRIS and Non-IRIS Individuals Enrolled in the Early and Late Arms of the Trial
3.3. Analysis of Activation Marker and KIR Repertoire in IRIS and Non-IRIS Individuals Enrolled in Early and Late Arms
3.4. Evolution of Lectin Receptors in IRIS and Non-IRIS Individuals
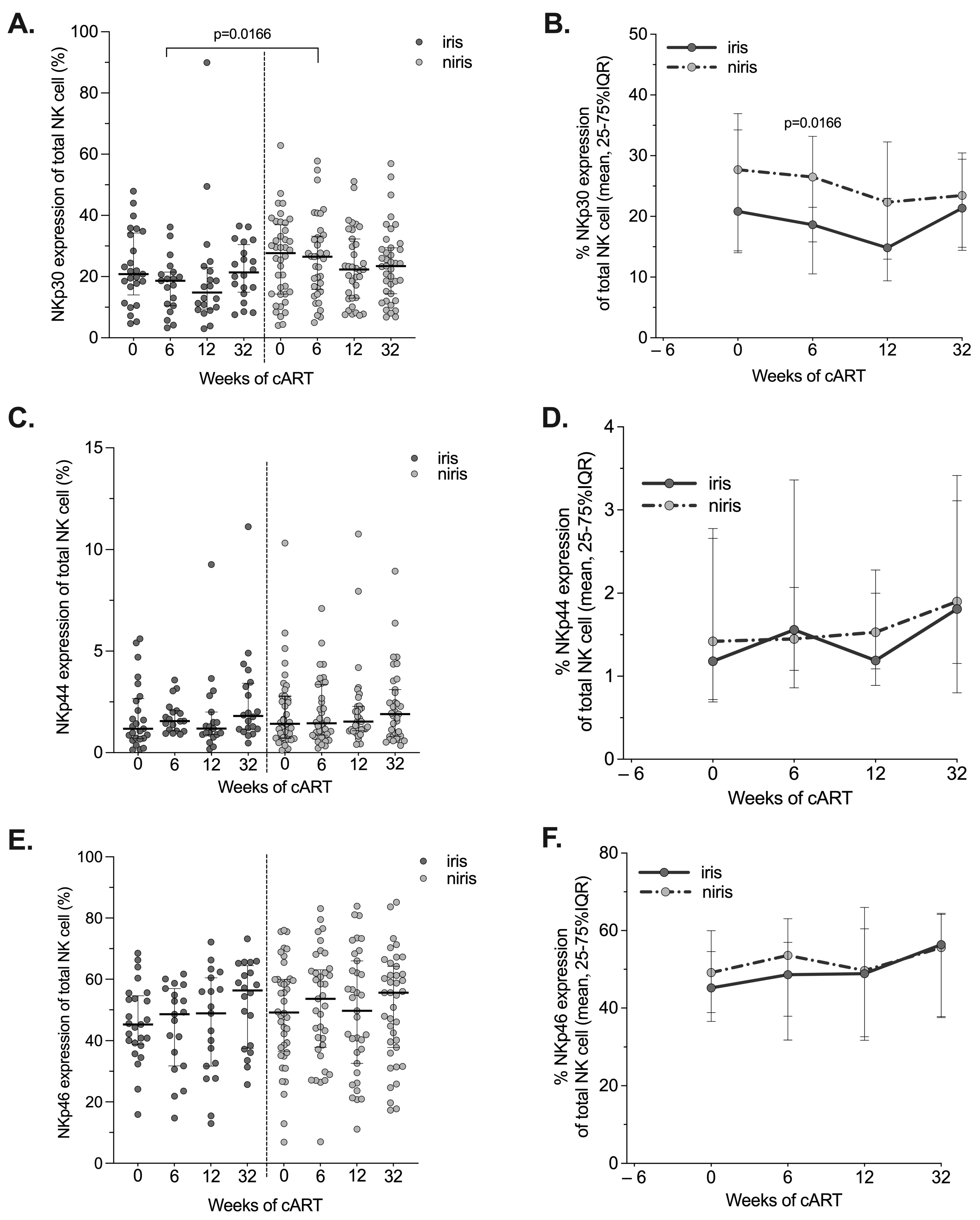
3.5. Expression of Natural Cytotoxic Receptors (NCRs) in IRIS and Non-IRIS during the Follow-Up
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. List of Study Team Members and Investigators in Cambodia
- CAMELIA study team members: Thim SOK, François-Xavier BLANC, Anne E. GOLDFELD, Didier LAUREILLARD, Claire REKACEWICZ, and Sirenda VONG.
- CAMELIA coordination centre: Phearavin PHENG, Manil SAMAN, Chanthy LENG, Sao Sarady AY, Phalla CHEA, Nimul Roat MEN, Sopheap KUN, Sokeo CHEA, Pichda TOUENG, Yong YOEUN, Keo Kunthea DY, Pheakun KRY, and Keolinelyanneth MEARDEY.
- Khmer Soviet Friendship Hospital: Narom PRAK, Chhun Im SIN, Chan Chhaya NGET, Sor KAING, Sovannara SONG, Bora AM, Sok CHEUNG, Tuon Seang Y, Seng Ly SAY, Sarann SUM, Seam CHHUN, Vanny SAY, Sokhoeun KAM, Chanthan UNG, TE Nai Sim, Sokim CHHOEUN, Chuop Chum KOH, Ratanak TONG, Cheung SOK, Saorin TITH, and Sithan CHAN.
- Svay Rieng Hospital: Kim khemarin LAK, Sath SUN, Sean YIM, Nimul CHAN EAR, Dara CHAN, Sokbophang VOEUNG, Phally BE, Chan Nimul EAR, Vanda PRAK, Thoeurn MEN, Thoeun KOY, Somaly THACH, Sinan CHHAY, and Sina MEAS.
- Donkeo Hospital: Chindamony KIM, Thong PHE, Saroeun UORNG, Sokhon KUOCH, Phea LAO, Chanthan YIM, Samros KHIEV, Sareth HEM, KEO Kimthan, UM Samoeun, TEANG Sophary, MEY imtheavy, Chanthon YIM, Bunthoeun HEANG, Hort SIENG, Huch MEAS, Sina SOK, Vibol NEAK, Nyn KEN, Kunthea TES, Soeun SAN, and Soeun SEAN.
References
- Bagcchi, S. WHO’s Global Tuberculosis Report 2022. Lancet Microbe 2023, 4, e20. [Google Scholar] [CrossRef] [PubMed]
- Badri, M.; Wilson, D.; Wood, R. Effect of highly active antiretroviral therapy on incidence of tuberculosis in South Africa: A cohort study. Lancet 2002, 359, 2059–2064. [Google Scholar] [CrossRef] [PubMed]
- Zolopa, A.; Andersen, J.; Powderly, W.; Sanchez, A.; Sanne, I.; Suckow, C.; Hogg, E.; Komarow, L. Early antiretroviral therapy reduces AIDS progression/death in individuals with acute opportunistic infections: A multicenter randomized strategy trial. PLoS ONE 2009, 4, e5575. [Google Scholar] [CrossRef] [PubMed]
- Blanc, F.X.; Sok, T.; Laureillard, D.; Borand, L.; Rekacewicz, C.; Nerrienet, E.; Madec, Y.; Marcy, O.; Chan, S.; Prak, N.; et al. Earlier versus later start of antiretroviral therapy in HIV-infected adults with tuberculosis. N. Engl. J. Med. 2011, 365, 1471–1481. [Google Scholar] [CrossRef] [PubMed]
- Walker, N.F.; Stek, C.; Wasserman, S.; Wilkinson, R.J.; Meintjes, G. The tuberculosis-associated immune reconstitution inflammatory syndrome: Recent advances in clinical and pathogenesis research. Curr. Opin. HIV AIDS 2018, 13, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Namale, P.E.; Abdullahi, L.H.; Fine, S.; Kamkuemah, M.; Wilkinson, R.J.; Meintjes, G. Paradoxical TB-IRIS in HIV-infected adults: A systematic review and meta-analysis. Future Microbiol. 2015, 10, 1077–1099. [Google Scholar] [CrossRef] [PubMed]
- Quinn, C.M.; Poplin, V.; Kasibante, J.; Yuquimpo, K.; Gakuru, J.; Cresswell, F.V.; Bahr, N.C. Tuberculosis IRIS: Pathogenesis, Presentation, and Management across the Spectrum of Disease. Life 2020, 10, 262. [Google Scholar] [CrossRef] [PubMed]
- Cevaal, P.M.; Bekker, L.-G.; Hermans, S. TB-IRIS pathogenesis and new strategies for intervention: Insights from related inflammatory disorders. Tuberculosis 2019, 118, 101863. [Google Scholar] [CrossRef]
- Silveira-Mattos, P.S.; Narendran, G.; Akrami, K.; Fukutani, K.F.; Anbalagan, S.; Nayak, K.; Subramanyam, S.; Subramani, R.; Vinhaes, C.L.; Souza, D.O.-d.; et al. Differential expression of CXCR3 and CCR6 on CD4+ T-lymphocytes with distinct memory phenotypes characterizes tuberculosis-associated immune reconstitution inflammatory syndrome. Sci. Rep. 2019, 9, 1502. [Google Scholar] [CrossRef]
- Tadokera, R.; Meintjes, G.; Skolimowska, K.H.; Wilkinson, K.A.; Matthews, K.; Seldon, R.; Chegou, N.N.; Maartens, G.; Rangaka, M.X.; Rebe, K.; et al. Hypercytokinaemia accompanies HIV-tuberculosis immune reconstitution inflammatory syndrome. Eur. Respir. J. 2011, 37, 1248–1259. [Google Scholar] [CrossRef]
- Lai, R.P.J.; Meintjes, G.; Wilkinson, K.A.; Graham, C.M.; Marais, S.; Van der Plas, H.; Deffur, A.; Schutz, C.; Bloom, C.; Munagala, I.; et al. HIV–tuberculosis-associated immune reconstitution inflammatory syndrome is characterized by Toll-like receptor and inflammasome signalling. Nat. Commun. 2015, 6, 8451. [Google Scholar] [CrossRef] [PubMed]
- Andrade, B.B.; Singh, A.; Narendran, G.; Schechter, M.E.; Nayak, K.; Subramanian, S.; Anbalagan, S.; Jensen, S.M.R.; Porter, B.O.; Antonelli, L.R.; et al. Mycobacterial Antigen Driven Activation of CD14++CD16− Monocytes Is a Predictor of Tuberculosis-Associated Immune Reconstitution Inflammatory Syndrome. PLoS Pathog. 2014, 10, e1004433. [Google Scholar] [CrossRef] [PubMed]
- Nakiwala, J.K.; Walker, N.F.; Diedrich, C.R.; Worodria, W.; Meintjes, G.; Wilkinson, R.J.; Mayanja-Kizza, H.; Colebunders, R.; Kestens, L.; Wilkinson, K.A.; et al. Neutrophil Activation and Enhanced Release of Granule Products in HIV-TB Immune Reconstitution Inflammatory Syndrome. JAIDS J. Acquir. Immune Defic. Syndr. 2018, 77, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhao, F.; Su, W.; Li, Q.; Li, J.; Ji, J.; Deng, Y.; Zhou, Y.; Wang, X.; Yang, H.; et al. Zinc finger and interferon-stimulated genes play a vital role in TB-IRIS following HAART in AIDS. Pers. Med. 2018, 15, 251–269. [Google Scholar] [CrossRef] [PubMed]
- de Sa, N.B.R.; de Souza, N.C.S.; Neira-Goulart, M.; Ribeiro-Alves, M.; Da Silva, T.P.; Pilotto, J.H.; Rolla, V.C.; Giacoia-Gripp, C.B.W.; de Oliveira Pinto, L.M.; Scott-Algara, D.; et al. Inflammasome genetic variants are associated with tuberculosis, HIV-1 infection, and TB/HIV-immune reconstitution inflammatory syndrome outcomes. Front. Cell Infect. Microbiol. 2022, 12, 962059. [Google Scholar] [CrossRef] [PubMed]
- Rocco, J.M.; Laidlaw, E.; Galindo, F.; Anderson, M.; Sortino, O.; Kuriakose, S.; Lisco, A.; Manion, M.; Sereti, I. Mycobacterial Immune Reconstitution Inflammatory Syndrome in HIV is Associated With Protein-Altering Variants in Hemophagocytic Lymphohistiocytosis-Related Genes. J. Infect. Dis. 2023, 228, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Laureillard, D.; Marcy, O.; Madec, Y.; Chea, S.; Chan, S.; Borand, L.; Fernandez, M.; Prak, N.; Kim, C.; Dim, B.; et al. Paradoxical tuberculosis-associated immune reconstitution inflammatory syndrome after early initiation of antiretroviral therapy in the camelia randomized. Aids 2013, 27, 2577–2586. [Google Scholar] [CrossRef] [PubMed]
- Michailidis, C.; Pozniak, A.L.; Mandalia, S.; Basnayake, S.; Nelson, M.R.; Gazzard, B.G. Clinical characteristics of IRIS syndrome in patients with HIV and tuberculosis. Antivir. Ther. 2005, 10, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef]
- Abel, A.M.; Yang, C.; Thakar, M.S.; Malarkannan, S. Natural Killer Cells: Development, Maturation, and Clinical Utilization. Front. Immunol. 2018, 9, 1869. [Google Scholar] [CrossRef]
- Pohlmeyer, C.W.; Gonzalez, V.D.; Irrinki, A.; Ramirez, R.N.; Li, L.; Mulato, A.; Murry, J.P.; Arvey, A.; Hoh, R.; Deeks, S.G.; et al. Identification of NK Cell Subpopulations That Differentiate HIV-Infected Subject Cohorts with Diverse Levels of Virus Control. J. Virol. 2019, 93, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Flórez-Álvarez, L.; Hernandez, J.C.; Zapata, W. NK Cells in HIV-1 Infection: From Basic Science to Vaccine Strategies. Front. Immunol. 2018, 9, 2290. [Google Scholar] [CrossRef] [PubMed]
- Scott-Algara, D.; Arnold, V.; Didier, C.; Kattan, T.; Pirozzi, G.; Barre-Sinoussi, F.; Pancino, G. The CD85j+ NK cell subset potently controls HIV-1 replication in autologous dendritic cells. PLoS ONE 2008, 3, e1975. [Google Scholar] [CrossRef] [PubMed]
- Nabatanzi, R.; Cose, S.; Joloba, M.; Jones, S.R.; Nakanjako, D. Effects of HIV infection and ART on phenotype and function of circulating monocytes, natural killer, and innate lymphoid cells. AIDS Res. Ther. 2018, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Li, Z.; Martin, L.; Hu, Z.; Wu, H.; Wan, Z.; Kilby, M.; Heath, S.L.; Huang, L.; Jiang, W. Increased Natural Killer Cell Activation in HIV-Infected Immunologic Non-Responders Correlates with CD4+ T Cell Recovery after Antiretroviral Therapy and Viral Suppression. PLoS ONE 2017, 12, e0167640. [Google Scholar] [CrossRef] [PubMed]
- Mavilio, D.; Benjamin, J.; Daucher, M.; Lombardo, G.; Kottilil, S.; Planta, M.A.; Marcenaro, E.; Bottino, C.; Moretta, L.; Moretta, A.; et al. Natural killer cells in HIV-1 infection: Dichotomous effects of viremia on inhibitory and activating receptors and their functional correlates. Proc. Natl. Acad. Sci. USA 2003, 100, 15011–15016. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Dai, Y.; Wang, Y.; Yang, Q.; Guo, J.; Wei, C.; Chen, W.; Huang, H.; Zhu, J.; Zhang, C.; et al. Single-cell transcriptomics of blood reveals a natural killer cell subset depletion in tuberculosis. EBioMedicine 2020, 53, 102686. [Google Scholar] [CrossRef] [PubMed]
- Roy Chowdhury, R.; Vallania, F.; Yang, Q.; Lopez Angel, C.J.; Darboe, F.; Penn-Nicholson, A.; Rozot, V.; Nemes, E.; Malherbe, S.T.; Ronacher, K.; et al. A multi-cohort study of the immune factors associated with M. tuberculosis infection outcomes. Nature 2018, 560, 644–648. [Google Scholar] [CrossRef]
- Kim, K.H.; An, D.R.; Song, J.; Yoon, J.Y.; Kim, H.S.; Yoon, H.J.; Im, H.N.; Kim, J.; Kim do, J.; Lee, S.J.; et al. Mycobacterium tuberculosis Eis protein initiates suppression of host immune responses by acetylation of DUSP16/MKP-7. Proc. Natl. Acad. Sci. USA 2012, 109, 7729–7734. [Google Scholar] [CrossRef]
- Abebe, F. Immunological basis of early clearance of Mycobacterium tuberculosis infection: The role of natural killer cells. Clin. Exp. Immunol. 2021, 204, 32–40. [Google Scholar] [CrossRef]
- Vankayalapati, R.; Barnes, P.F. Innate and adaptive immune responses to human Mycobacterium tuberculosis infection. Tuberculosis 2009, 89, S77–S80. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Triebold, K.J.; Dalton, D.K.; Stewart, T.A.; Bloom, B.R. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J. Exp. Med. 1993, 178, 2249–2254. [Google Scholar] [CrossRef] [PubMed]
- Brillantes, M.; Beaulieu, A.M. Memory and Memory-Like NK Cell Responses to Microbial Pathogens. Front. Cell Infect. Microbiol. 2020, 10, 102. [Google Scholar] [CrossRef] [PubMed]
- Venkatasubramanian, S.; Cheekatla, S.; Paidipally, P.; Tripathi, D.; Welch, E.; Tvinnereim, A.R.; Nurieva, R.; Vankayalapati, R. IL-21-dependent expansion of memory-like NK cells enhances protective immune responses against Mycobacterium tuberculosis. Mucosal Immunol. 2017, 10, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R.K.; Li, H.; Jost, S.; Blass, E.; Li, H.; Schafer, J.L.; Varner, V.; Manickam, C.; Eslamizar, L.; Altfeld, M.; et al. Antigen-specific NK cell memory in rhesus macaques. Nat. Immunol. 2015, 16, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Ramana Rao, P.V.; Rajasekaran, S.; Raja, A. Augumentation of natural killer activity with exogenous interleukins in patients with HIV and pulmonary tuberculosis coinfection. AIDS Res. Hum. Retroviruses 2008, 24, 1435–1443. [Google Scholar] [PubMed]
- Pean, P.; Nerrienet, E.; Madec, Y.; Borand, L.; Laureillard, D.; Fernandez, M.; Marcy, O.; Sarin, C.; Phon, K.; Taylor, S.; et al. Natural killer cell degranulation capacity predicts early onset of the immune reconstitution inflammatory syndrome (IRIS) in HIV-infected patients with tuberculosis. Blood 2012, 119, 3315–3320. [Google Scholar] [CrossRef]
- Pean, P.; Nouhin, J.; Ratana, M.; Madec, Y.; Borand, L.; Marcy, O.; Laureillard, D.; Fernandez, M.; Barré-Sinoussi, F.; Weiss, L.; et al. High Activation of γδ T Cells and the γδ2pos T-Cell Subset Is Associated With the Onset of Tuberculosis-Associated Immune Reconstitution Inflammatory Syndrome, ANRS 12153 CAPRI NK. Front. Immunol. 2019, 10, 2018. [Google Scholar] [CrossRef]
- Maria, A.D.; Fogli, M.; Costa, P.; Murdaca, G.; Puppo, F.; Mavilio, D.; Moretta, A.; Moretta, L. The impaired NK cell cytolytic function in viremic HIV-1 infection is associated with a reduced surface expression of natural cytotoxicity receptors (NKp46, NKp30 and NKp44). Eur. J. Immunol. 2003, 33, 2410–2418. [Google Scholar] [CrossRef]
- Nirmala, R.; Narayanan, P.R.; Mathew, R.; Maran, M.; Deivanayagam, C.N. Reduced NK activity in pulmonary tuberculosis patients with/without HIV infection: Identifying the defective stage and studying the effect of interleukins on NK activity. Tuberculosis 2001, 81, 343–352. [Google Scholar] [CrossRef]
- Harris, L.D.; Khayumbi, J.; Ongalo, J.; Sasser, L.E.; Tonui, J.; Campbell, A.; Odhiambo, F.H.; Ouma, S.G.; Alter, G.; Gandhi, N.R.; et al. Distinct Human NK Cell Phenotypes and Functional Responses to Mycobacterium tuberculosis in Adults From TB Endemic and Non-endemic Regions. Front. Cell Infect. Microbiol. 2020, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Bozzano, F.; Costa, P.; Passalacqua, G.; Dodi, F.; Ravera, S.; Pagano, G.; Canonica, G.W.; Moretta, L.; De Maria, A. Functionally relevant decreases in activatory receptor expression on NK cells are associated with pulmonary tuberculosis in vivo and persist after successful treatment. Int. Immunol. 2009, 21, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Schierloh, P.; Yokobori, N.; Aleman, M.; Musella, R.M.; Beigier-Bompadre, M.; Saab, M.A.; Alves, L.; Abbate, E.; de la Barrera, S.S.; Sasiain, M.C. Increased susceptibility to apoptosis of CD56dimCD16+ NK cells induces the enrichment of IFN-gamma-producing CD56bright cells in tuberculous pleurisy. J. Immunol. 2005, 175, 6852–6860. [Google Scholar] [CrossRef] [PubMed]
- Esaulova, E.; Das, S.; Singh, D.K.; Choreño-Parra, J.A.; Swain, A.; Arthur, L.; Rangel-Moreno, J.; Ahmed, M.; Singh, B.; Gupta, A.; et al. The immune landscape in tuberculosis reveals populations linked to disease and latency. Cell Host Microbe 2021, 29, 165–178.e168. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.; Meyer, D.; Grosse, V.; Stoll, M.; Schmidt, R.E.; Heiken, H. Reconstitution of NK cell activity in HIV-1 infected individuals receiving antiretroviral therapy. Immunobiology 2000, 202, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Zhang, X.; Su, B.; Liu, L.-F.; Yang, X.-D.; Tang, B.; Xia, H.; Ma, P.; Zhang, T.; Wu, H. Increased early activation of CD56dimCD16dim/- natural killer cells in immunological non-responders correlates with CD4+ T-cell recovery. Chin. Med. J. 2020, 133, 2928–2939. [Google Scholar] [CrossRef] [PubMed]
- Azzoni, L.; Chehimi, J.; Zhou, L.; Foulkes, A.S.; June, R.; Maino, V.C.; Landay, A.; Rinaldo, C.; Jacobson, L.P.; Montaner, L.J. Early and delayed benefits of HIV-1 suppression: Timeline of recovery of innate immunity effector cells. Aids 2007, 21, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Frias, M.; Rivero-Juarez, A.; Gordon, A.; Camacho, A.; Cantisan, S.; Cuenca-Lopez, F.; Torre-Cisneros, J.; Pena, J.; Rivero, A. Persistence of pathological distribution of NK cells in HIV-infected patients with prolonged use of HAART and a sustained immune response. PLoS ONE 2015, 10, e0121019. [Google Scholar] [CrossRef]
- Bayigga, L.; Nabatanzi, R.; Sekiziyivu, P.N.; Mayanja-Kizza, H.; Kamya, M.R.; Kambugu, A.; Olobo, J.; Kiragga, A.; Kirimunda, S.; Joloba, M.; et al. High CD56++CD16− natural killer (NK) cells among suboptimal immune responders after four years of suppressive antiretroviral therapy in an African adult HIV treatment cohort. BMC Immunol. 2014, 15, 2. [Google Scholar] [CrossRef]
- Nabatanzi, R.; Bayigga, L.; Cose, S.; Rowland-Jones, S.; Canderan, G.; Joloba, M.; Nakanjako, D. Aberrant natural killer (NK) cell activation and dysfunction among ART-treated HIV-infected adults in an African cohort. Clin. Immunol. 2019, 201, 55–60. [Google Scholar] [CrossRef]
- Bisio, F.; Bozzano, F.; Marras, F.; Di Biagio, A.; Moretta, L.; De Maria, A. Successfully treated HIV-infected patients have differential expression of NK cell receptors (NKp46 and NKp30) according to AIDS status at presentation. Immunol. Lett. 2013, 152, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, L.R.; Mahnke, Y.; Hodge, J.N.; Porter, B.O.; Barber, D.L.; DerSimonian, R.; Greenwald, J.H.; Roby, G.; Mican, J.; Sher, A.; et al. Elevated frequencies of highly activated CD4+ T cells in HIV+ patients developing immune reconstitution inflammatory syndrome. Blood 2010, 116, 3818–3827. [Google Scholar] [CrossRef] [PubMed]
- Conradie, F.; Foulkes, A.S.; Ive, P.; Yin, X.; Roussos, K.; Glencross, D.K.; Lawrie, D.; Stevens, W.; Montaner, L.J.; Sanne, I.; et al. Natural Killer Cell Activation Distinguishes Mycobacterium tuberculosis-Mediated Immune Reconstitution Syndrome From Chronic HIV and HIV/MTB Coinfection. J. Acquir. Immune Defic. Syndr. 2011, 58, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hearps, A.C.; Agius, P.A.; Zhou, J.; Brunt, S.; Chachage, M.; Angelovich, T.A.; Cameron, P.U.; Giles, M.; Price, P.; Elliott, J.; et al. Persistence of Activated and Adaptive-Like NK Cells in HIV+ Individuals despite 2 Years of Suppressive Combination Antiretroviral Therapy. Front. Immunol. 2017, 8, 731. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tian, Z.; Wei, H. Developmental and Functional Control of Natural Killer Cells by Cytokines. Front. Immunol. 2017, 8, 930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xu, J.; Hong, K.; Yuan, L.; Peng, H.; Tang, H.; Ma, P.; Zhang, Y.; Xing, H.; Ruan, Y.; et al. Increased NKG2A found in cytotoxic natural killer subset in HIV-1 patients with advanced clinical status. AIDS 2007, 21 (Suppl. S8), S9–S17. [Google Scholar] [CrossRef] [PubMed]
- Parasa, V.R.; Sikhamani, R.; Raja, A. Effect of recombinant cytokines on the expression of natural killer cell receptors from patients with TB or/and HIV infection. PLoS ONE 2012, 7, e37448. [Google Scholar] [CrossRef] [PubMed]
- Brunetta, E.; Fogli, M.; Varchetta, S.; Bozzo, L.; Hudspeth, K.L.; Marcenaro, E.; Moretta, A.; Mavilio, D. Chronic HIV-1 viremia reverses NKG2A/NKG2C ratio on natural killer cells in patients with human cytomegalovirus co-infection. Aids 2010, 24, 27–34. [Google Scholar] [CrossRef]
- Giacoia-Gripp, C.B.W.; Cazote, A.D.S.; da Silva, T.P.; Sant’Anna, F.M.; Schmaltz, C.A.S.; Brum, T.S.; de Matos, J.A.; Silva, J.; Benjamin, A.; Pilotto, J.H.; et al. Changes in the NK Cell Repertoire Related to Initiation of TB Treatment and Onset of Immune Reconstitution Inflammatory Syndrome in TB/HIV Co-infected Patients in Rio de Janeiro, Brazil-ANRS 12274. Front. Immunol. 2019, 10, 1800. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pean, P.; Madec, Y.; Nerrienet, E.; Borand, L.; Laureillard, D.; Fernandez, M.; Marcy, O.; Scott-Algara, D., on behalf of the CAMELIA Study Team. Natural Killer Repertoire Restoration in TB/HIV Co-Infected Individuals Experienced an Immune Reconstitution Syndrome (CAMELIA Trial, ANRS 12153). Pathogens 2023, 12, 1241. https://doi.org/10.3390/pathogens12101241
Pean P, Madec Y, Nerrienet E, Borand L, Laureillard D, Fernandez M, Marcy O, Scott-Algara D on behalf of the CAMELIA Study Team. Natural Killer Repertoire Restoration in TB/HIV Co-Infected Individuals Experienced an Immune Reconstitution Syndrome (CAMELIA Trial, ANRS 12153). Pathogens. 2023; 12(10):1241. https://doi.org/10.3390/pathogens12101241
Chicago/Turabian StylePean, Polidy, Yoann Madec, Eric Nerrienet, Laurence Borand, Didier Laureillard, Marcelo Fernandez, Olivier Marcy, and Daniel Scott-Algara on behalf of the CAMELIA Study Team. 2023. "Natural Killer Repertoire Restoration in TB/HIV Co-Infected Individuals Experienced an Immune Reconstitution Syndrome (CAMELIA Trial, ANRS 12153)" Pathogens 12, no. 10: 1241. https://doi.org/10.3390/pathogens12101241
APA StylePean, P., Madec, Y., Nerrienet, E., Borand, L., Laureillard, D., Fernandez, M., Marcy, O., & Scott-Algara, D., on behalf of the CAMELIA Study Team. (2023). Natural Killer Repertoire Restoration in TB/HIV Co-Infected Individuals Experienced an Immune Reconstitution Syndrome (CAMELIA Trial, ANRS 12153). Pathogens, 12(10), 1241. https://doi.org/10.3390/pathogens12101241